
Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Bacterial Inocula
2.2. Design of the Experiment
2.3. Phagocytic Activity Analysis
2.4. Homogenization of Tissues and Isolation of Total RNA of IgA, MUC-1 and MUC-2
2.5. Relative Expression of IgA, MUC-1 and MUC-2 by Quantitative Real-Time PCR
2.6. Isolation of Lymphocytes from Mesenteric Lymph Nodes
2.7. Lymphocyte Identification
2.8. Serum Biochemical Analysis
2.9. Histological Processing of Stomach and Histo-Fluorescent In-Situ Hybridisation Analysis
2.10. Statistical Analysis
3. Results
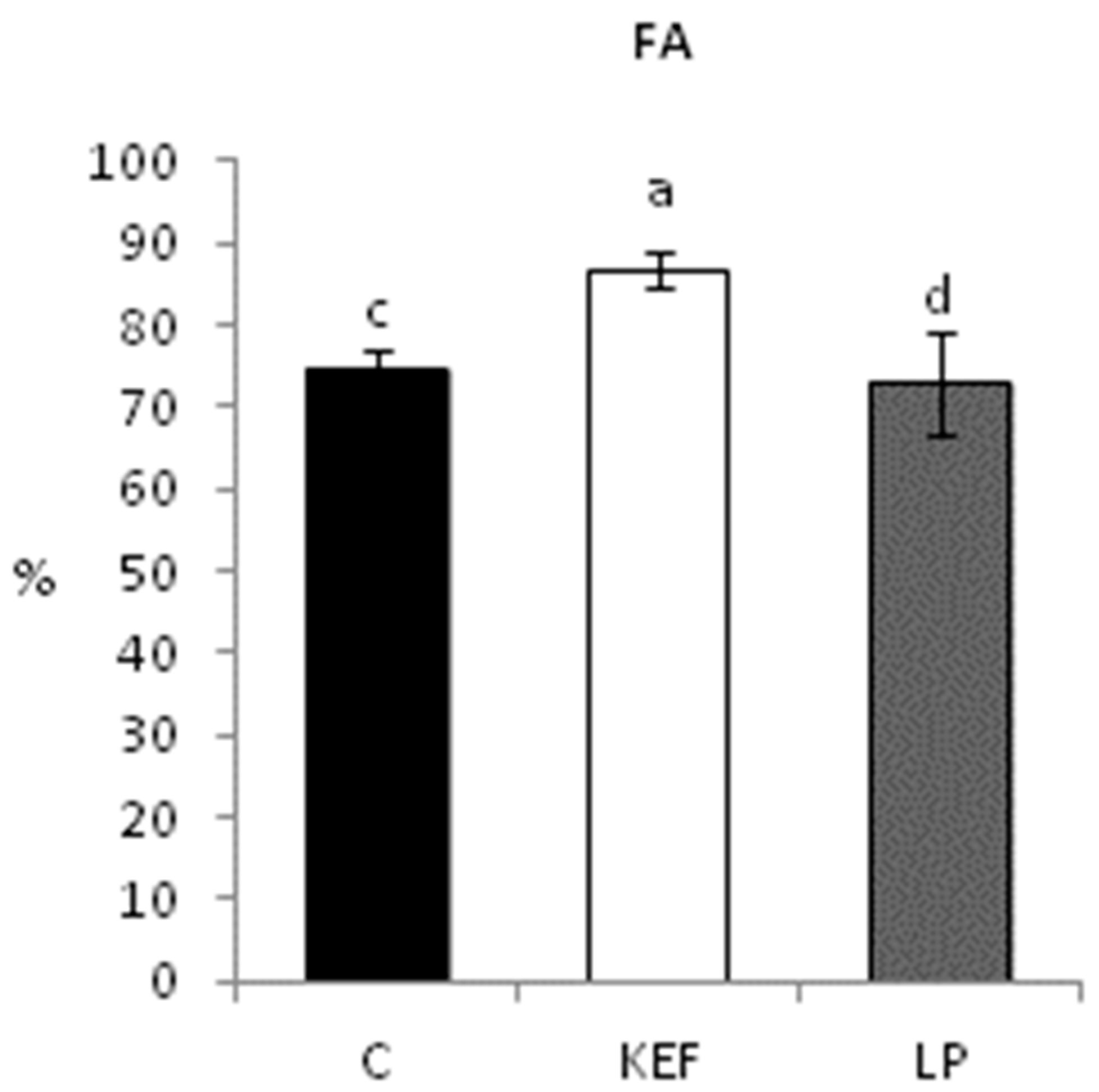
3.1. Phagocytic Activity
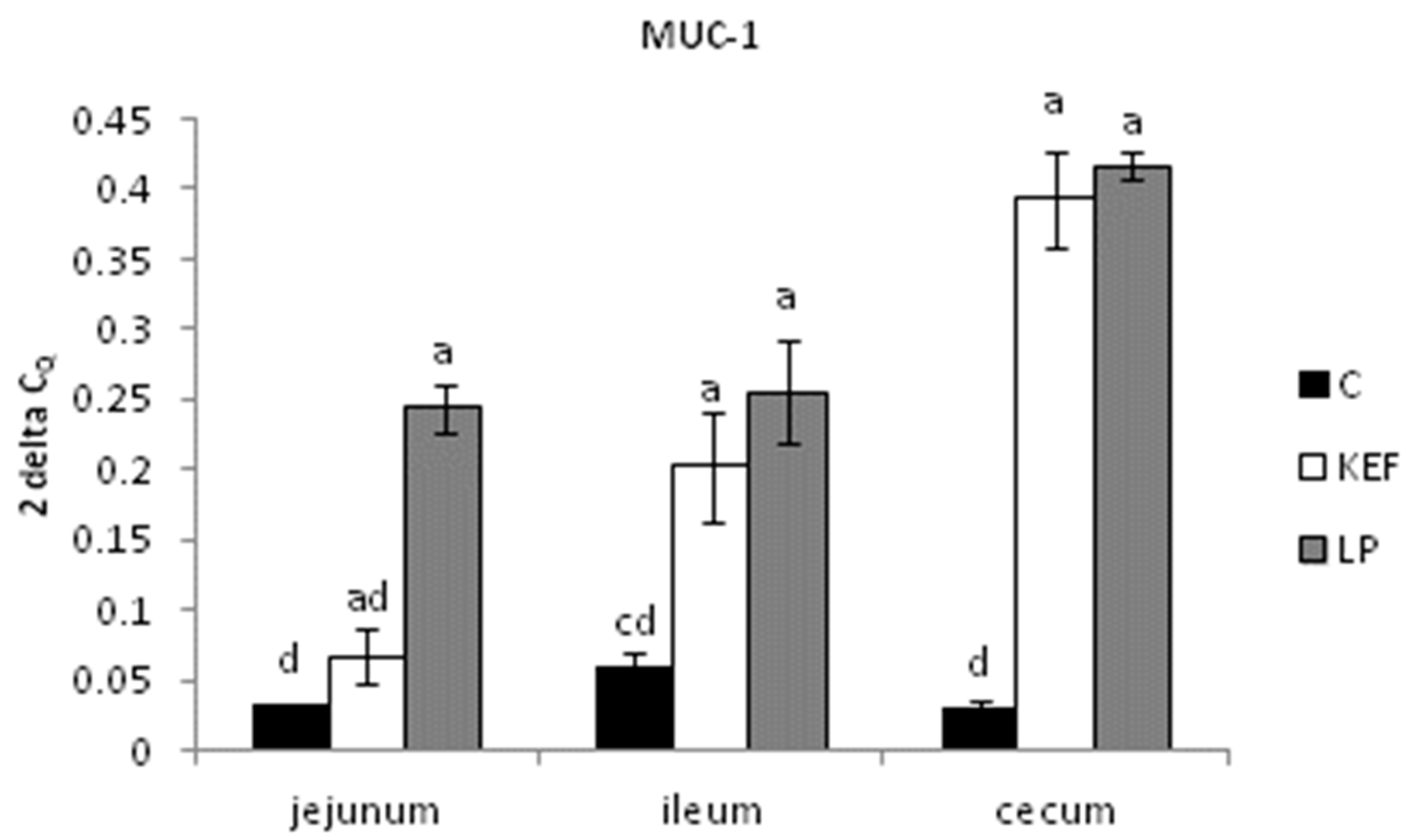
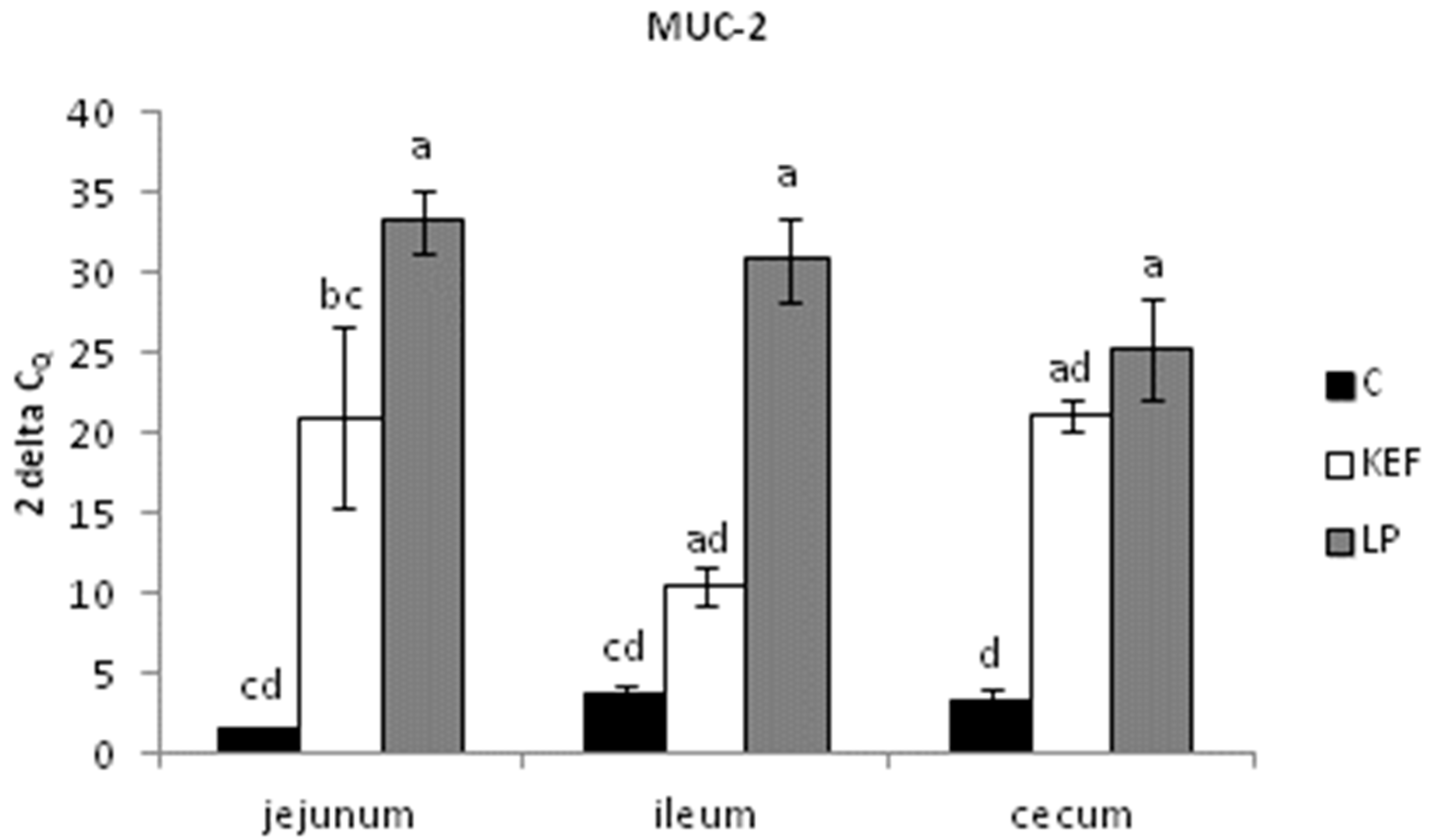
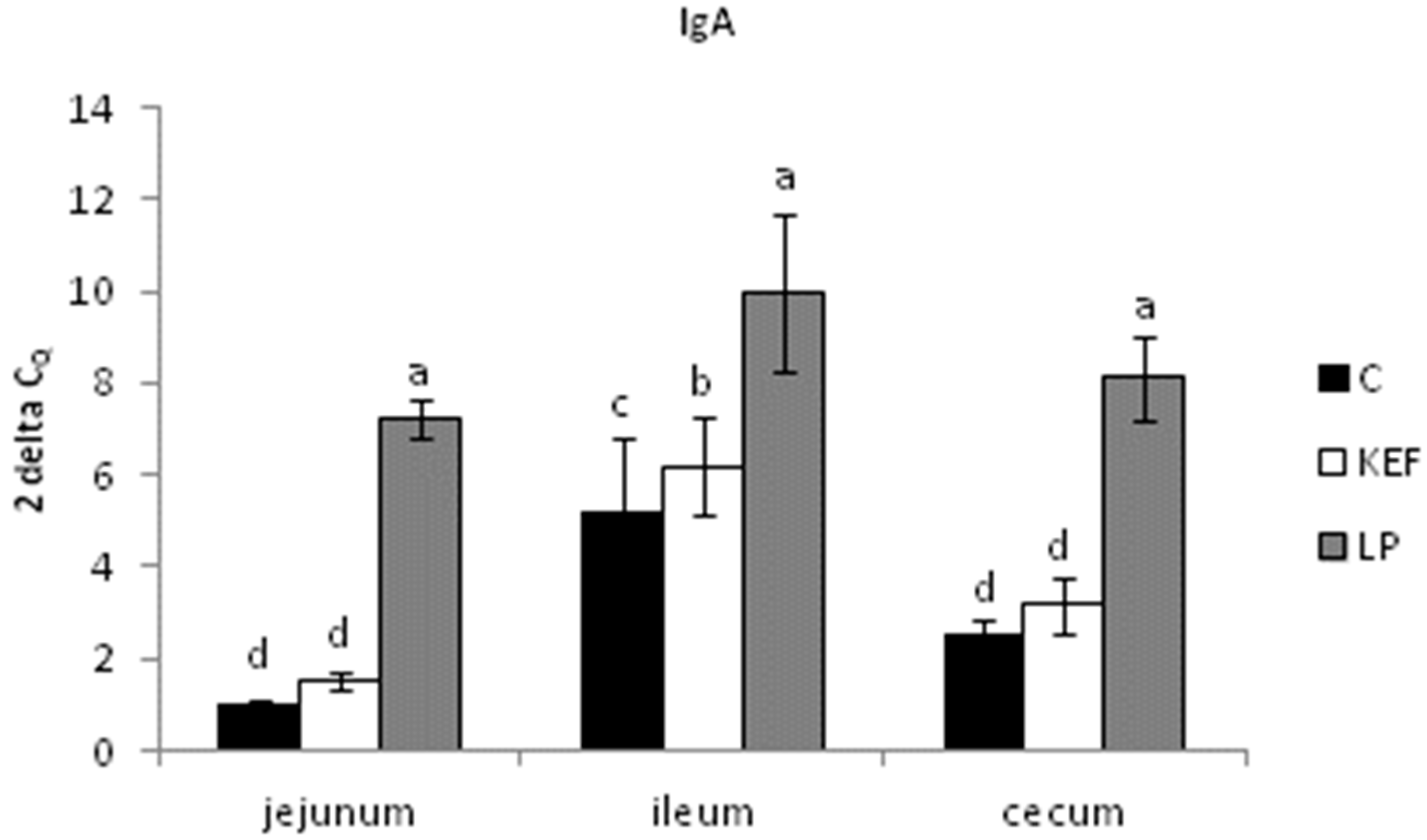
3.2. Relative Expression of MUC-1, MUC-2 and IgA, Evaluated by Quantitative Real-Time PCR
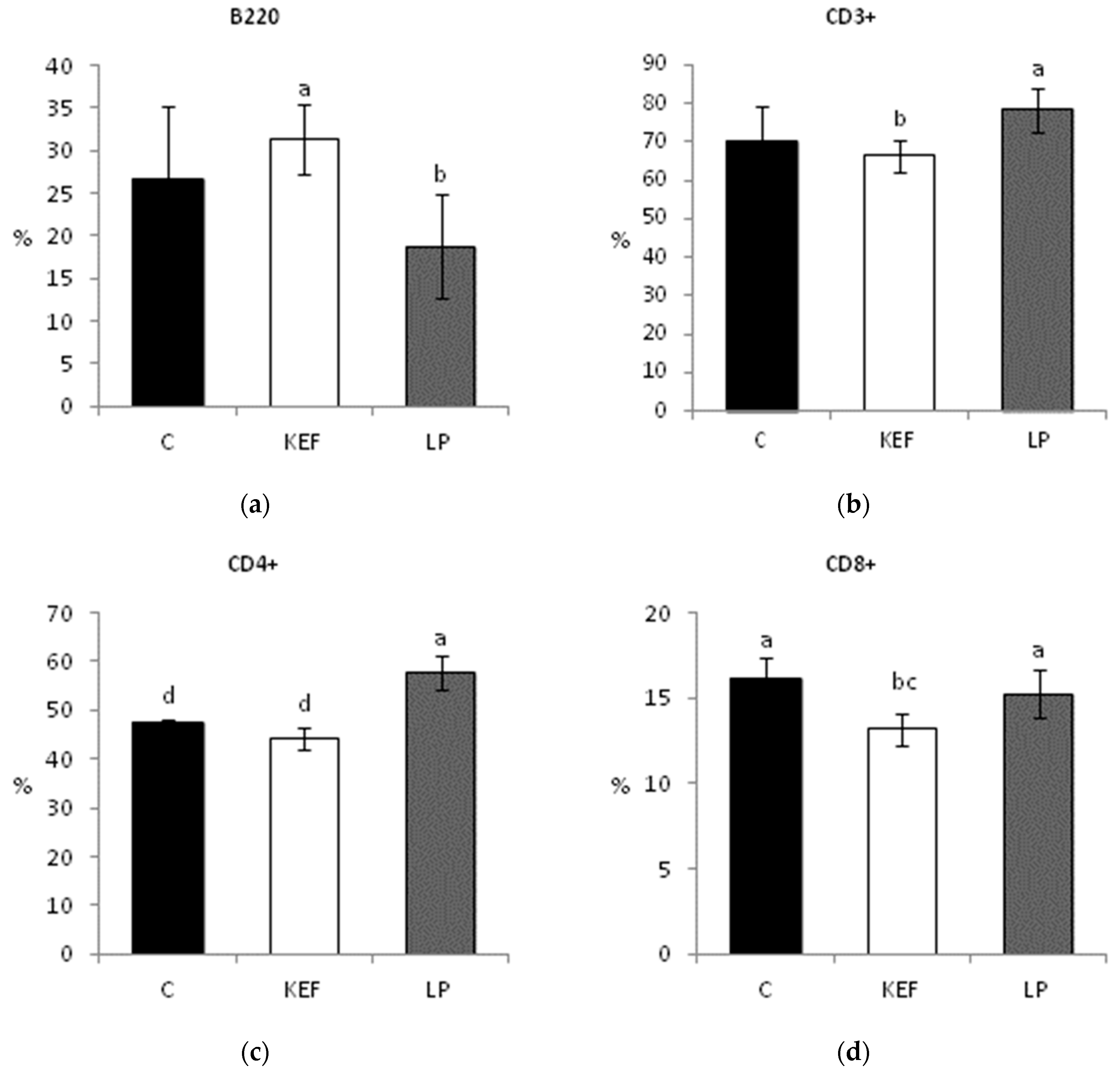
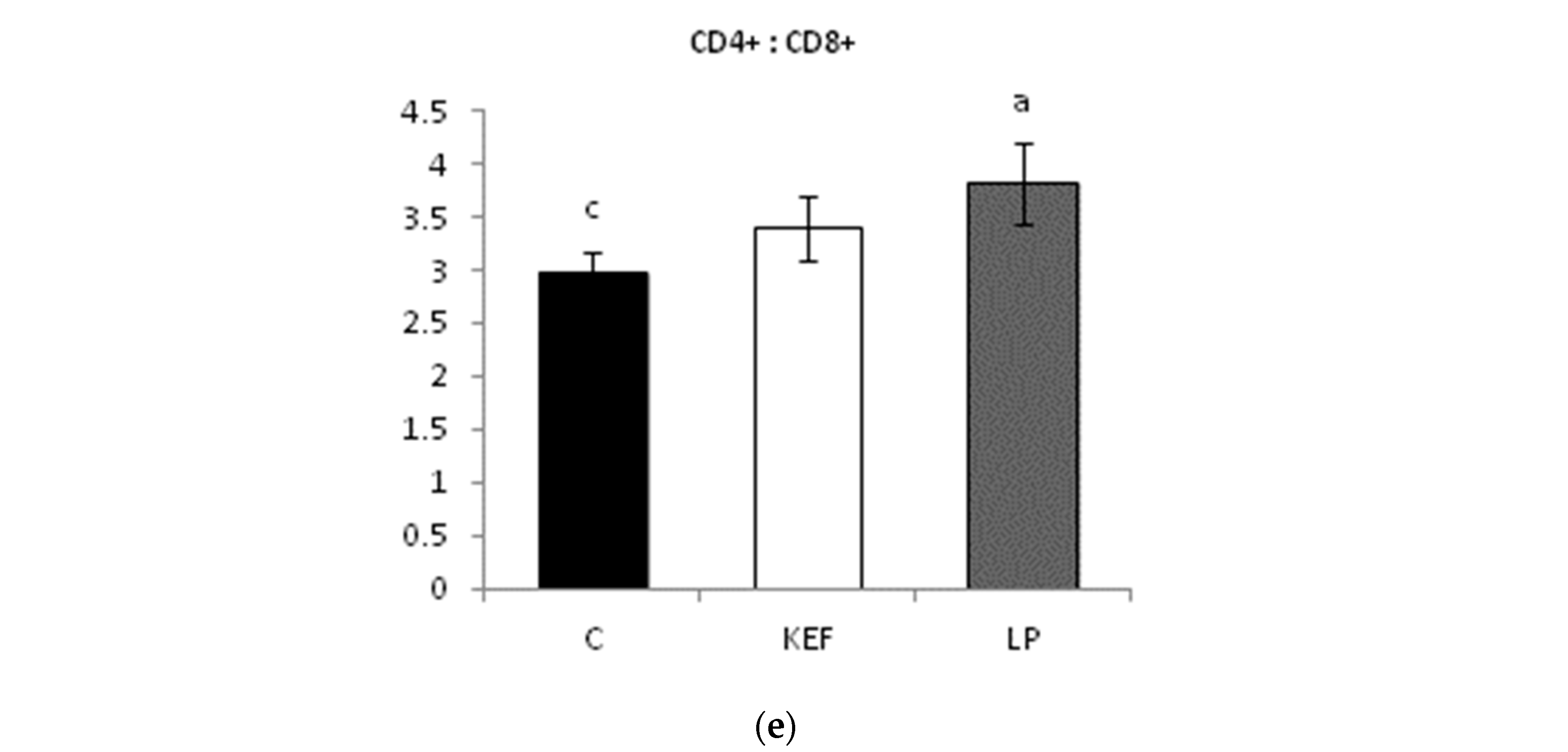
3.3. Proportions of Lymphocyte Subpopulations in Peripheral Blood
3.4. Proportions of Lymphocyte Subpopulations in Mesenteric Lymph Nodes
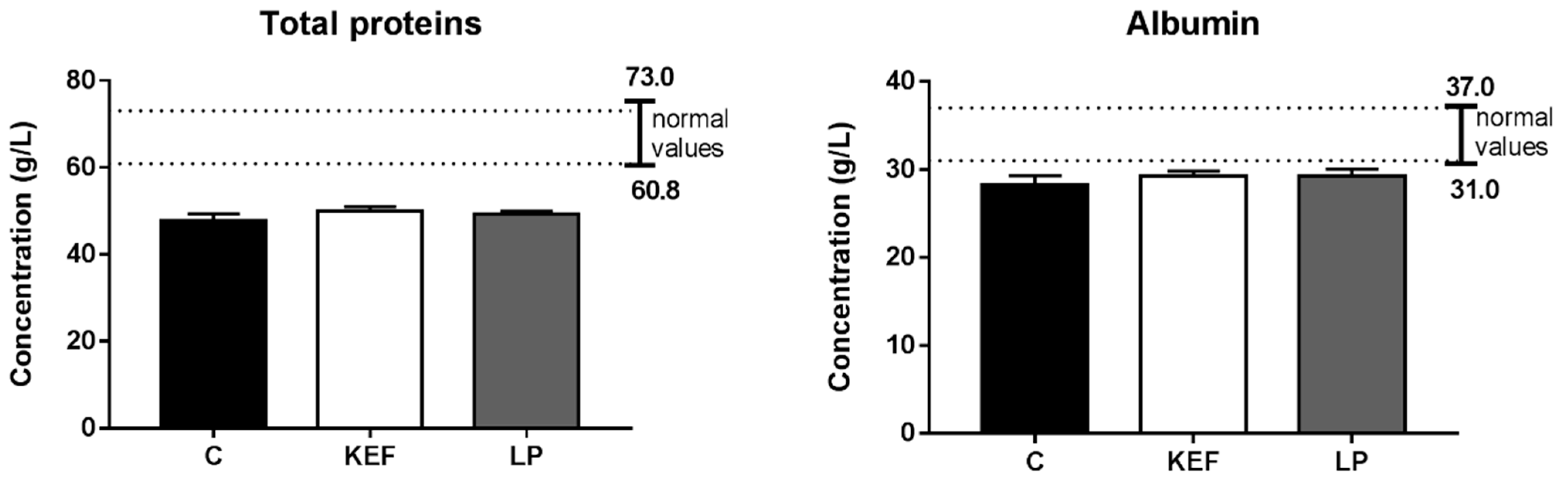
3.5. Nitrogen Profile
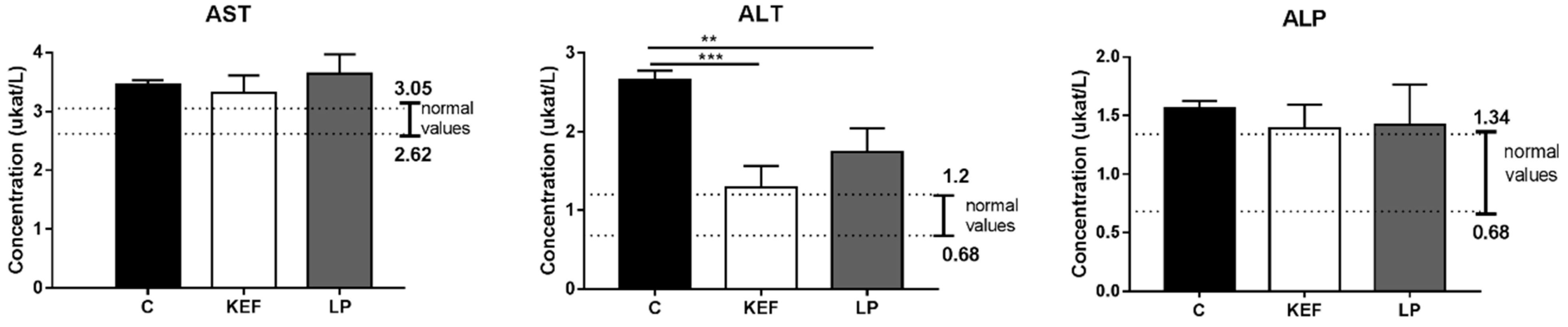
3.6. Enzymatic Profile
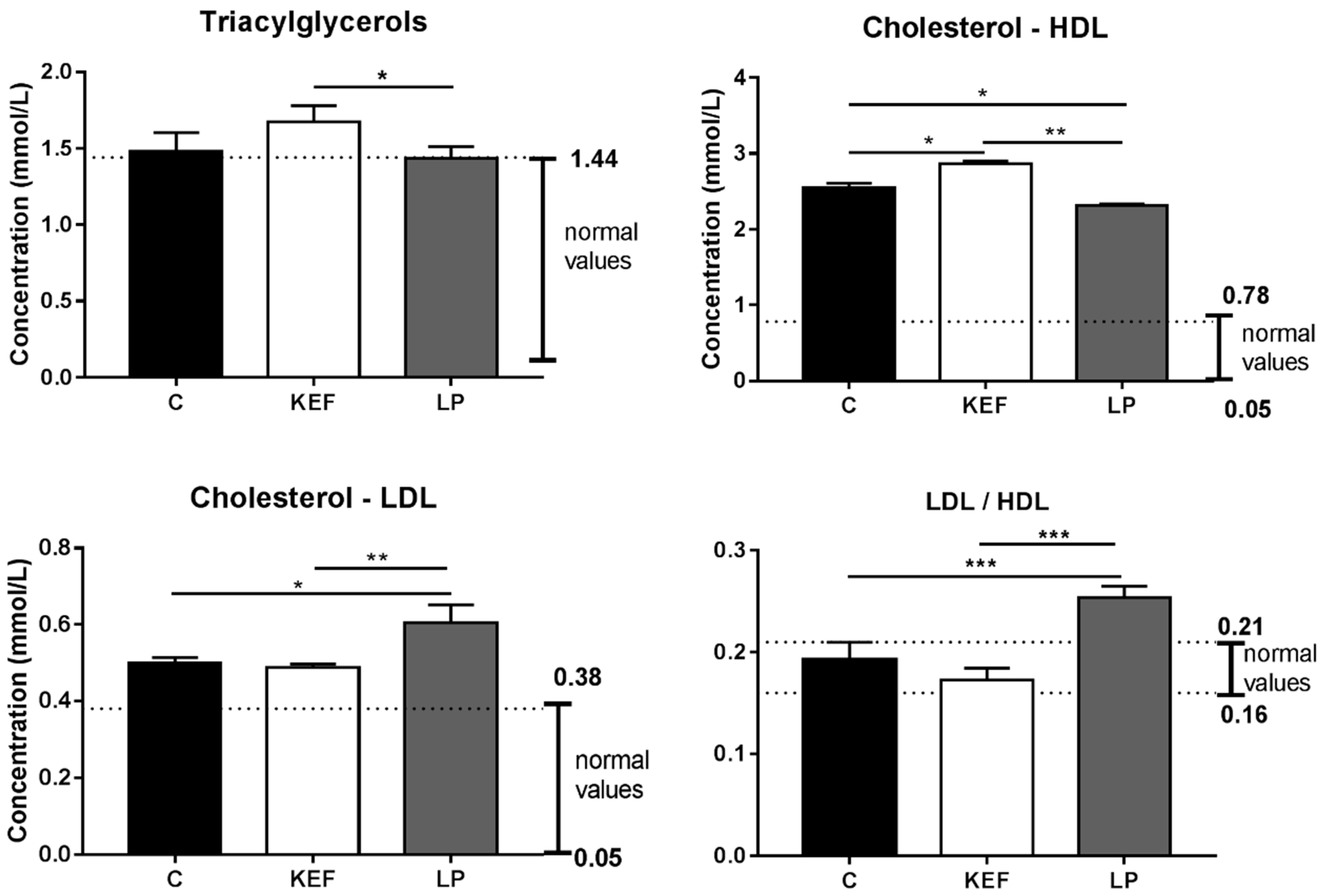
3.7. Lipid Profile
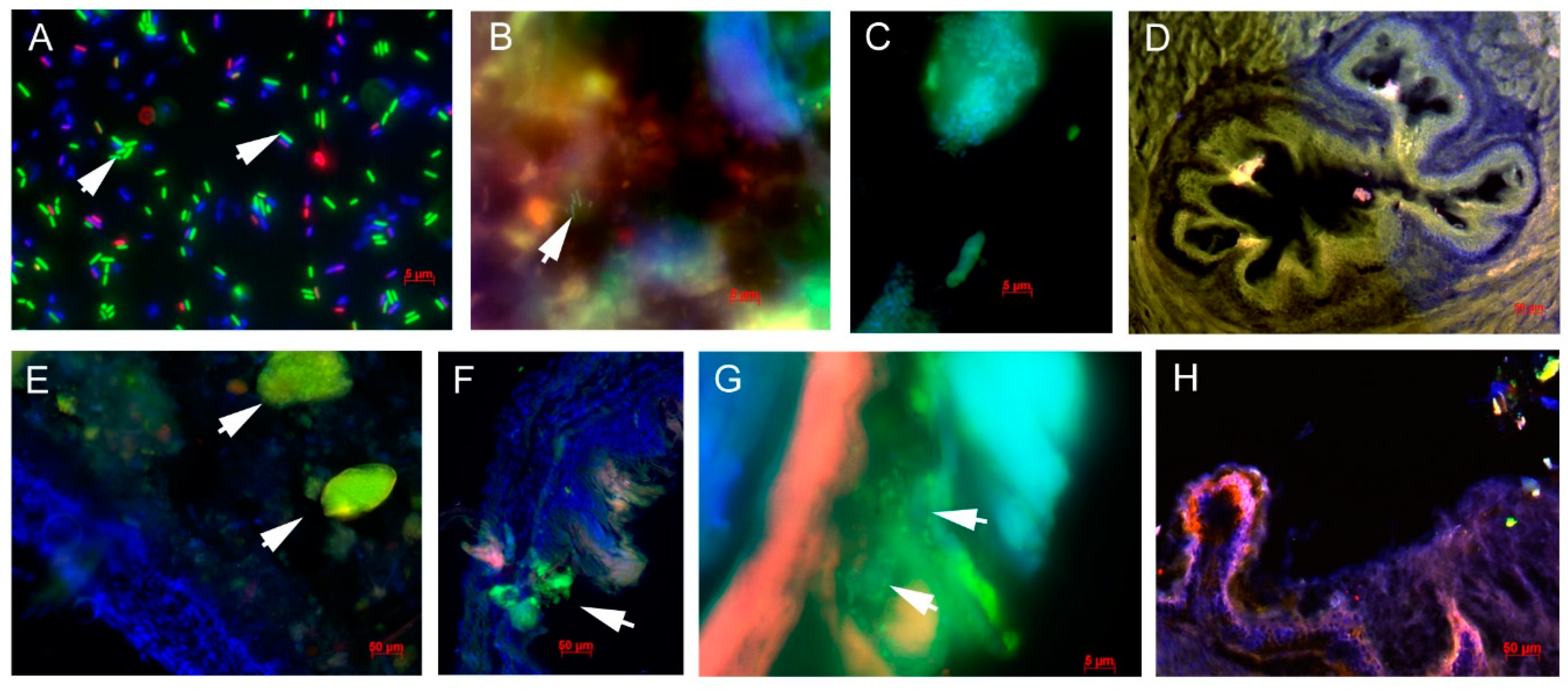
3.8. Biofilm Formation by Lactobacilli in the GIT
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leite, A.M.O.; Leite, D.C.A.; Del Aguila, E.M.; Alvares, T.S.; Peixoto, R.S.; Miguel, M.A.L.; Silva, J.T.; Paschoalin, V.M.F. Microbiological and chemical characteristics of Brazilian kefir during fermentation and storage processes. J. Dairy Sci. 2013, 96, 4149–4159. [Google Scholar] [CrossRef]
- Prado, M.R.; Blandón, L.M.; Vandenberghe, L.P.; Rodrigues, C.; Castro, G.R.; Thomaz-Soccol, V.; Soccol, C.R. Milk kefir: Composition, microbial cultures, biological activities, and related products. Front. Microbiol. 2015, 6, 1177. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, F.S.; Luaces-Regueira, M.; Ton, A.M.; Campagnaro, B.P.; Campos-Toimil, M.; Pereira, T.M.; Vasquez, E.C. Mechanisms of action of kefir in chronic cardiovascular and metabolic diseases. Cell Physiol. Biochem. 2018, 48, 1901–1914. [Google Scholar] [CrossRef]
- Yılmaz, İ.; Dolar, M.E.; Özpınar, H. Effect of administering kefir on the changes in fecal microbiota and symptoms of inflammatory bowel disease: A randomized controlled trial. Turk. J. Gastroenterol. 2019, 30, 242–253. [Google Scholar] [CrossRef]
- Englerová, K.; Nemcová, R.; Mudroňová, D. The study of the probiotic potential of the beneficial bacteria isolated from kefir grains. Folia Vet. 2017, 61, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Rosa, D.D.; Dias, M.M.S.; Grześkowiak, Ł.M.; Reis, S.A.; Conceição, L.L.; Peluzio, M.D.C.G. Milk kefir: Nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, J.; Zhao, L.; Zhang, M.; Guo, H.; Ren, F. Effect of oral administration of Lactobacillus paracasei Lí on mouse systemic immunity and the immune response in the intestine. Arch. Biol. Sci. 2016, 68, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Human intestinal dendritic cells decrease cytokine release against Salmonella infection in the presence of Lactobacillus paracasei upon TLR activation. PLoS ONE 2012, 7, e43197. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.; Sugrue, I.; Tobin, C.; Hill, C.; Stanton, C.; Ross, R.P. The Lactobacillus casei group: History and health related applications. Front. Microbiol. 2018, 9, 2107. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Invernici, M.M.; Furlaneto, F.A.C.; Messora, M.R. Effectiveness of multi-strain versus single-strain probiotics–current status and recommendations for the future. J. Clin. Gastroenterol. 2018, 52, S35–S40. [Google Scholar] [CrossRef]
- Sabico, S.; Al-Mashharawi, A.; Al-Daghri, N.M.; Yakout, S.; Alnaami, A.M.; Alokail, M.S.; McTernan, P.G. Effects of a multi-strain probiotic supplement for 12 weeks in circulating endotoxin levels and cardio-metabolic profiles of medication naïve T2DM patients: A randomized clinical trial. J. Transl. Med. 2017, 15, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.C.; de Sousa, R.G.; Botelho, P.B.; Gomes, T.L.; Prada, P.O.; Mota, J.F. The additional effects of a probiotic mix on abdominal adiposity and antioxidant status. A double-blind, randomized trial. Obesity 2017, 25, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Van Hemert, S.; Ormel, G. Influence of the multispecies probiotic Ecologic® Barrier on parameters of intestinal barrier function. Food Nutr. Sci. 2014, 5, 1739–1745. [Google Scholar] [CrossRef] [Green Version]
- Knight, P.A.; Wright, S.H.; Brown, J.K.; Huang, X.; Sheppard, D.; Miller, H.R.P. Enteric expression of the integrin αvβ6 is essential for nematode-induced mucosal mast cell hyperplasia and expression of the granule chymase, mouse mast cell protease-1. Am. J. Pathol. 2002, 161, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Rachagani, S.; Torres, M.P.; Kumar, S.; Haridas, D.; Baine, M.; Macha, M.A.; Kaur, S.; Ponnusamy, M.P.; Dey, P.; Seshacharyulu, P.; et al. Mucin (Muc) expression during pancreatic cancer progression in spontaneous mouse model: Potential implications for diagnosis and therapy. J. Haematol. Oncol. 2012, 5, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wlodarska, M.; Willing, B.; Keeney, K.M.; Menendez, A.; Bergstrom, K.S.; Gill, N.; Russell, S.L.; Vallance, B.A.; Finlay, B.B. Antibiotic treatment alters the colonic mucus layer and predisposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect. Immun. 2011, 79, 1536–1545. [Google Scholar] [CrossRef] [Green Version]
- Aravindaram, K.; Kuo, T.Y.; Lan, C.W.; Yu, H.H.; Wang, P.H.; Chen, Y.S.; Chen, G.H.; Yang, N.S. Protective immunity against porcine circovirus 2 in mice induced by a gene-based combination vaccination. J. Gene Med. 2009, 11, 288–301. [Google Scholar] [CrossRef]
- Madar, M.; Slizova, M.; Czerwinski, J.; Hrckova, G.; Mudronova, D.; Gancarcikova, S.; Popper, M.; Pistl, J.; Soltys, J.; Nemcova, R. Histo-FISH protocol to detect bacterial compositions and biofilms formation in vivo. Benef. Microbes 2015, 6, 899–907. [Google Scholar] [CrossRef]
- Czerwinski, J.; Hojberg, O.; Smulikowska, S.; Engberg, R.M.; Mieczkowska, A. Effects of sodium butyrate and salinomycin upon intestinal microbiota, mucosal morphology and performance of broiler chickens. Arch. Anim. Nutr. 2012, 66, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Popper, M.; Gancarčíková, S.; Maďar, M.; Mudroňová, D.; Hrčková, G.; Nemcová, R. Amoxicillin-clavulanic acid and ciprofloxacin-treated SPF mice as gnotobiotic model. Appl. Microbiol. Biotechnol. 2016, 100, 9671–9682. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.A.M.; Miguel, M.A.; Peixoto, R.S.; Rosado, A.S.; Silva, J.T.; Paschoalin, V.M. Microbiological, technological and therapeutic properties of kefir: A natural probiotic beverage. Braz. J. Microbiol. 2013, 44, 341–349. [Google Scholar] [CrossRef]
- Vinderola, G.; Perdigon, G.; Duarte, J.; Thangavel, D.; Farnworth, E.; Matar, C. Effects of kefir fractions on innate immunity. Immunobiology 2006, 211, 149–156. [Google Scholar] [CrossRef]
- Llewellyn, A.; Foey, A. Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events. Nutrients 2017, 9, 1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extra-cellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Kusumo, P.D.; Bela, B.; Wibowo, H.; Munasir, Z.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation increases faecal sIgA and immune response in children younger than two years. Benef. Microbes 2019, 10, 245–252. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Zavala, L.; Carasi, P.; Trejo, S.A.; Bronsoms, S.; Serradell, M.L.A.; Garrote, G.L.; Abraham, A.G. (2018) Simulated gastrointestinal conditions increase adhesion ability of Lactobacillus paracasei strains isolated from kefir to CACO-2 cells and mucin. Food Res. Int. 2018, 103, 462–467. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.H.; Whang, K.Y.; Kim, Y.J.; Oh, S. Inhibition of Escherichia coli O157:H7 attachment by interactions between lactic acid bacteria and intestinal epithelial cells. J. Microbiol. Biotechnol. 2008, 18, 1278–1285. [Google Scholar] [PubMed]
- Kumar, M.; Kissoon-Singh, V.; Coria, A.L.; Moreau, F.; Chadee, K. Probiotic mixture VSL#3 reduces colonic inflammation and improves intestinal barrier function in MUC2 mucin-deficient mice. Am. J. Physiol. Gastrointest. Physiol. 2017, 312, G34–G45. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Lagstrom, H.; Suomalainen, T.; Salminen, S. Effect of probiotics on constipation, fecal azoreductase activity and fecal mucin content in the elderly. Ann. Nutr. Metab. 2002, 46, 159–162. [Google Scholar] [CrossRef]
- Carasi, P.; Racedo, S.M.; Jacquot, C.; Romanin, D.E.; Serradell, M.A.; Urdaci, M.C. Impact of kefir derived Lactobacillus kefiri on the mucosal immune response and gut microbiota. J. Immunol. Res. 2015, 2015, 361604. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.T.; Cheng, P.C.; Pan, T.M. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Appl. Microbiol. Biotechnol. 2012, 96, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Iwabuchi, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Hachimura, S. Orally administered heat-killed Lactobacillus paracasei MCC1849 enhances antigen-specific IgA secretion and induces follicular helper T cells in mice. PLoS ONE 2018, 13, e0199018. [Google Scholar] [CrossRef]
- Yamasaki-Yashiki, S.; Miyoshi, Y.; Nakayama, T.; Kunisawa, J.; Katakura, Y. IgA-enhancing effects of membrane vesicles derived from Lactobacillus sakei subsp. sakei NBRC15893. Biosci. Microbiota Food Health 2019, 38, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef]
- Bunker, J.J.; Bendelac, A. IgA responses to microbiota. Immunity 2018, 49, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Nurliyani, N.; Harmayani, E.; Soesatyo, M.H. Leukocytes count and lymphocyte proliferation of dinitrochlorobenzene sensitized rat supplemented with fermented goat milk. World Acad. Sci. Eng. Technol. Int. J. Nutr. Food Eng. 2011, 5, 357–361. [Google Scholar] [CrossRef]
- Kim, H.W.; Ju, D.B.; Kye, Y.C.; Ju, Y.J.; Kim, C.G.; Lee, I.K.; Park, S.M.; Choi, I.S.; Cho, K.K.; Lee, S.H.; et al. Galectin-9 induced by dietary probiotic mixture regulates immune balance to reduce atopic dermatitis symptoms in mice. Front. Immunol. 2020, 10, 3063. [Google Scholar] [CrossRef]
- Hong, W.S.; Chen, Y.P.; Chen, M.J. The antiallergic effect of kefir Lactobacilli. J. Food Sci. 2010, 75, H244–H253. [Google Scholar] [CrossRef]
- Adiloğlu, A.K.; Gönülateş, N.; Işler, M.; Senol, A. The effect of kefir consumption on human immune system: A cytokine study. Mikrobiyol. Bul. 2013, 47, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Franco, M.C.; Golowczyc, M.A.; De Antoni, G.L.; Pérez, P.F.; Humen, M.; Serradell, M.L.A. Administration of kefir-fermented milk protects mice against Giardia intestinalis infection. J. Med. Microbiol. 2013, 62, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Raras, T.Y.M.; Rusmini, H.; Wisudanti, D.D.; Chozin, I.N. Kefir stimulates anti-inflammatory response in TB-AFB (+) patients. Pak. J. Nutr. 2015, 14, 330–334. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, C.G.; Duarte, J.; Thangavel, D.; Perdigón, G.; Farnworth, E.; Matar, C. Immunomodulating capacity of kefir. J. Dairy Res. 2005, 72, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.R.; Wang, S.Y.; Chen, M.J.; Chen, H.L.; Yueh, P.Y.; Lin, C.W. Hypocholesterolaemic effects of milk-kefir and soyamilk-kefir in cholesterol-fed hamsters. Br. J. Nutr. 2006, 95, 939–946. [Google Scholar] [CrossRef]
- Maeda, H.; Zhu, X.; Omura, K.; Suzuki, S.; Kitamura, S. Effects of an exopolysaccharide (kefiran) on lipids, blood pressure, blood glucose, and constipation. Biofactors 2004, 22, 197–200. [Google Scholar] [CrossRef]
- Choi, J.W.; Kang, H.W.; Lim, W.C.; Kim, M.K.; Lee, I.Y.; Cho, H.Y. Kefir prevented excess fat accumulation in diet-induced obese mice. Biosci. Biotechnol. Biochem. 2017, 81, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The microbiota and health promoting characteristics of the fermented beverage kefir. Front. Microbiol. 2016, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.L.; Tsai, T.C.; Tsai, Y.C.; Liao, J.W.; Yen, C.C.; Chen, C.M. Kefir peptides prevent high-fructose corn syrup-induced non-alcoholic fatty liver disease in a murine model by modulation of inflammation and the JAK2 signaling pathway. Nutr. Diabetes 2016, 6, e237. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, N.; Xi, A.; Ahmed, Z.; Zhang, B.; Bai, X. Effects of Lactobacillus plantarum MA2 isolated from Tibet kefir on lipid metabolism and intestinal microflora of rats fed on high-cholesterol diet. Appl. Microbiol. Biotechnol. 2009, 84, 341–347. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, X.; Wang, J.; Wu, F.; Sui, Y.; Yang, L.; Wang, Z. Lactobacillus plantarum strains as potential probiotic cultures with cholesterol-lowering activity. J Dairy Sci. 2013, 96, 2746–2753. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′–3′ | Annealing/Temperature Time | References |
|---|---|---|---|
| IgA For | TCTCCTCCTCTTCTTGTCATACGC | 58 °C/40 s | [14] |
| IgA Rev | GGAGGTAAGTACCACAGGAGCGTTT | ||
| MUC-1 For | CCACCACTCCAGTTTACAGT | 56 °C/30 s | [15] |
| MUC-1 Rev | GAATGATAGCTGAGCCTGAC | ||
| MUC-2 For | GCTGACGAGTGGTTGGTGAATG | 60 °C/30 s | [16] |
| MUC-2 Rev | GATGAGGTGGCAGACAGGAGAC | ||
| GAPDH For | CATCACTGCCACCCAGAAGACTGTGGA | 60 °C/30 s | [17] |
| GAPDH Rev | TACTCCTTGGAGGCCATGTAGGCCATG |
| Type | Fluorochrome | Clone | Isotype | Concentration | Amount |
|---|---|---|---|---|---|
| anti-CD4 | FITC | GK 1.5 | IgG2b, κ | 0.5 mg/mL | 2 μL |
| anti-CD8a | R-PE | 53–6.7 | IgG2a, κ | 0.2 mg/mL | 1 μL |
| anti-CD3 | PerCP-eFluor710 | 17A2 | IgG2b, κ | 0.2 mg/mL | 2 μL |
| anti-B220 | APC | RA3–6B2 | IgG2a, κ | 0.2 mg/mL | 1 μL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaffová, V.; Mudroňová, D.; Mad’ar, M.; Hrčková, G.; Faixová, D.; Gancarčíková, S.; Ševčíková, Z.; Nemcová, R. Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms 2021, 9, 831. https://doi.org/10.3390/microorganisms9040831
Karaffová V, Mudroňová D, Mad’ar M, Hrčková G, Faixová D, Gancarčíková S, Ševčíková Z, Nemcová R. Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms. 2021; 9(4):831. https://doi.org/10.3390/microorganisms9040831
Chicago/Turabian StyleKaraffová, Viera, Dagmar Mudroňová, Marián Mad’ar, Gabriela Hrčková, Dominika Faixová, Soňa Gancarčíková, Zuzana Ševčíková, and Radomíra Nemcová. 2021. "Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains" Microorganisms 9, no. 4: 831. https://doi.org/10.3390/microorganisms9040831
APA StyleKaraffová, V., Mudroňová, D., Mad’ar, M., Hrčková, G., Faixová, D., Gancarčíková, S., Ševčíková, Z., & Nemcová, R. (2021). Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms, 9(4), 831. https://doi.org/10.3390/microorganisms9040831