
Probiotic and Metabolic Characterization of Vaginal Lactobacilli for a Potential Use in Functional Foods

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Biolog Phenotype Microarray Analysis
2.3. Hydrophobicity
2.4. Auto-Aggregation Assay
2.5. Bile Salts Deconjugation
2.6. Caco-2 Cell Adhesion Tests
2.7. Strain Tolerance to Simulated Digestion Process in Milk
2.8. Statistical Analysis
3. Results and Discussion
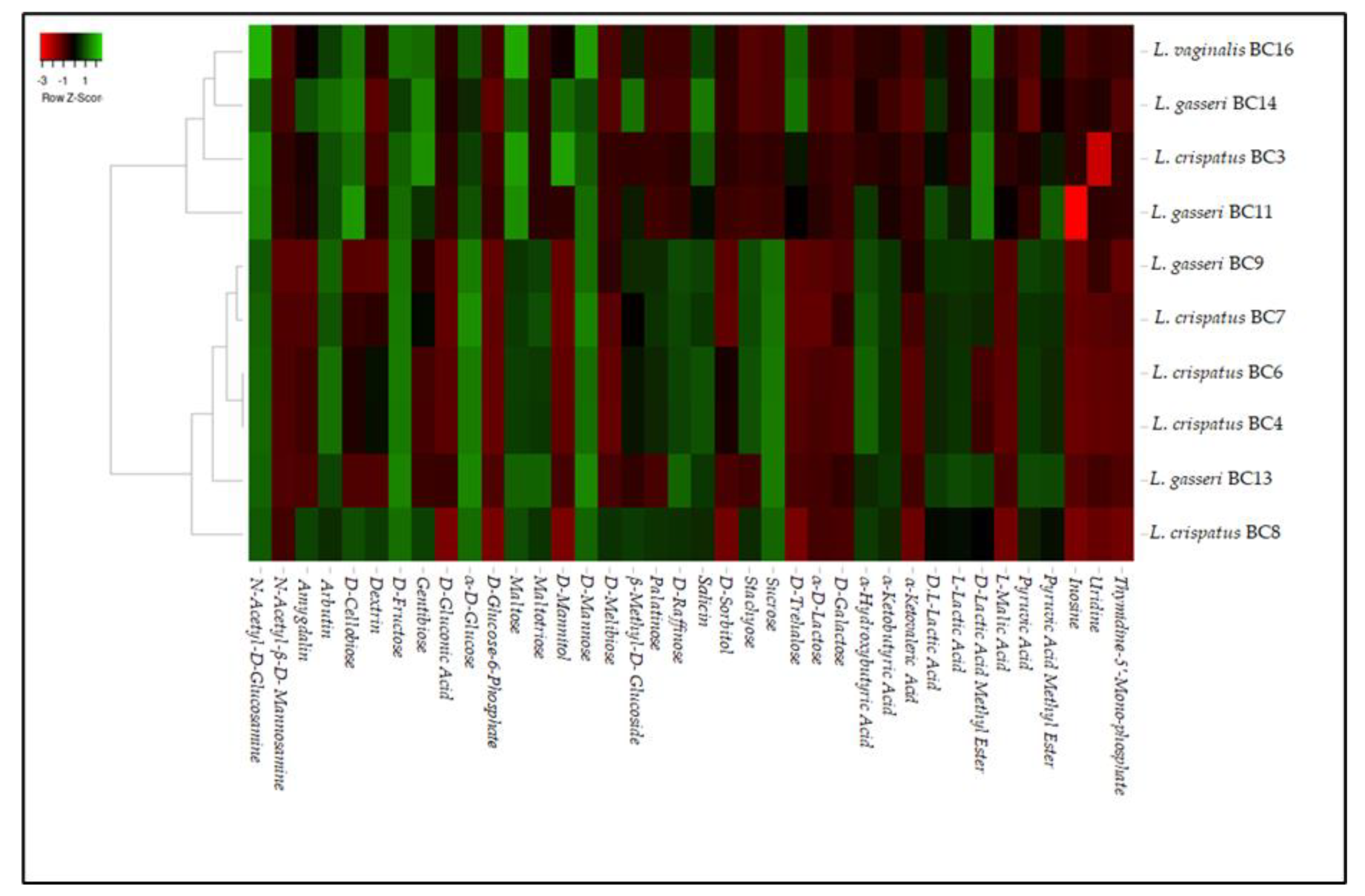
3.1. Phenotype Microarray Analysis
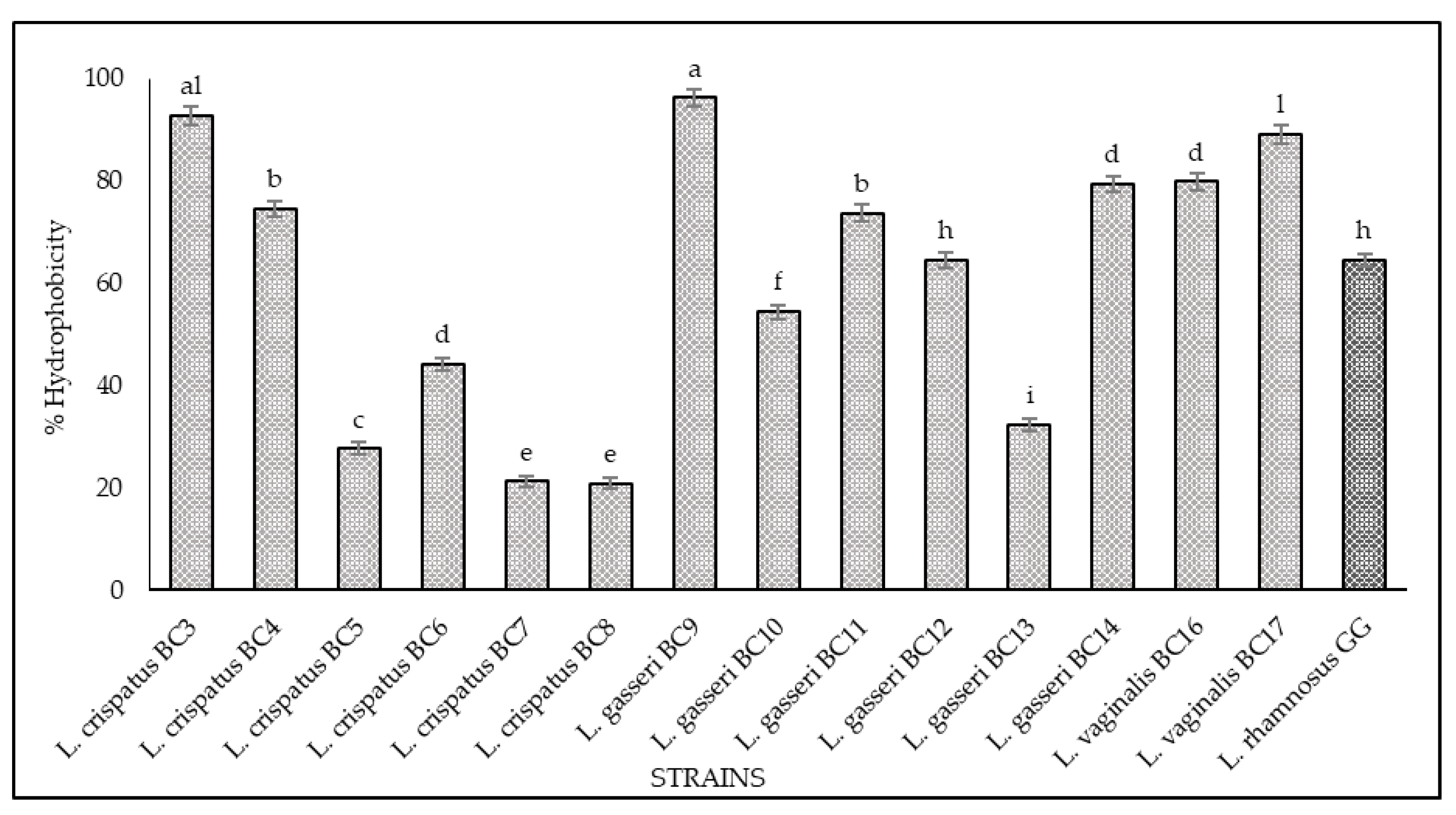
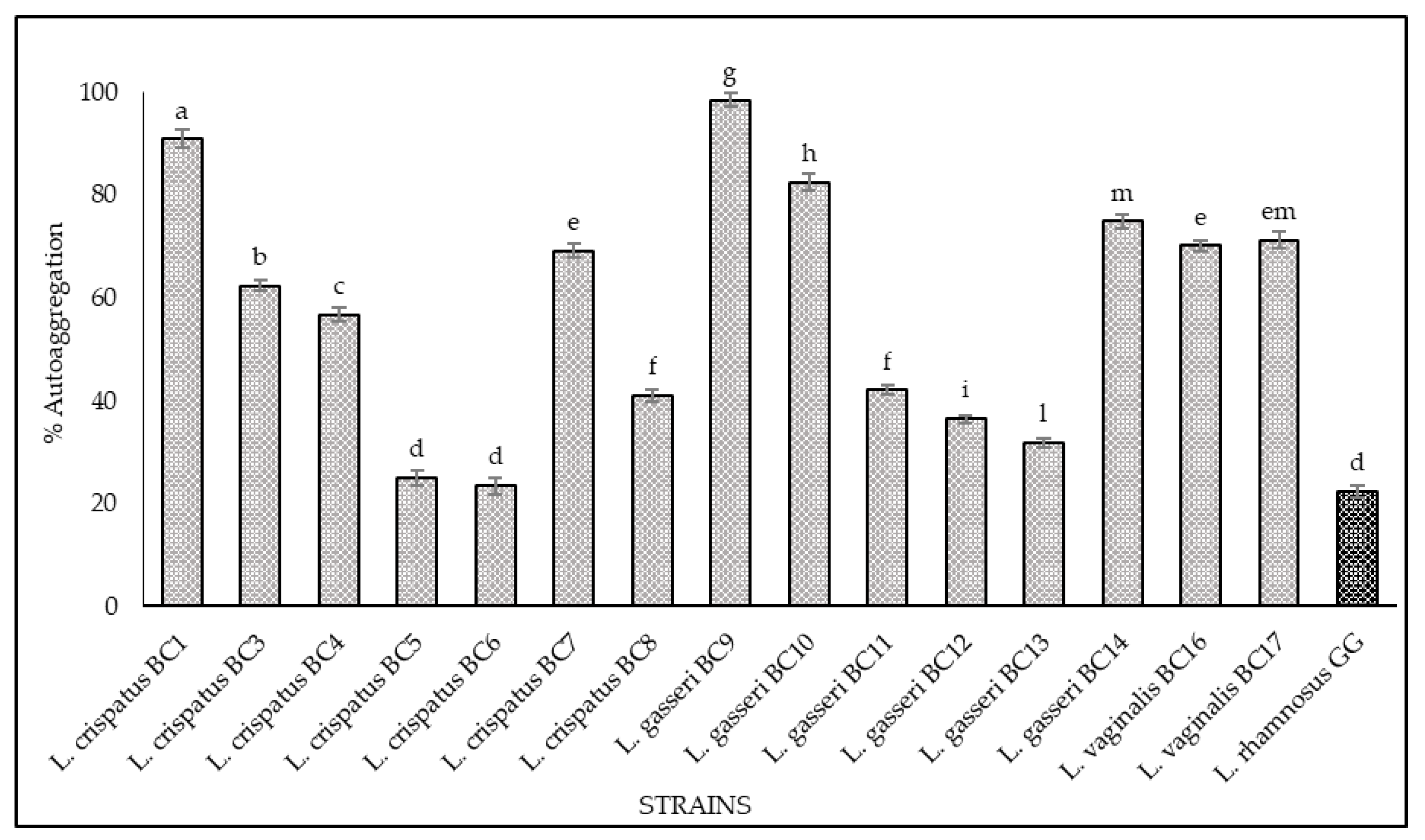
3.2. Hydrophobicity and Auto-Aggregation
3.3. Bile salts Deconjugation
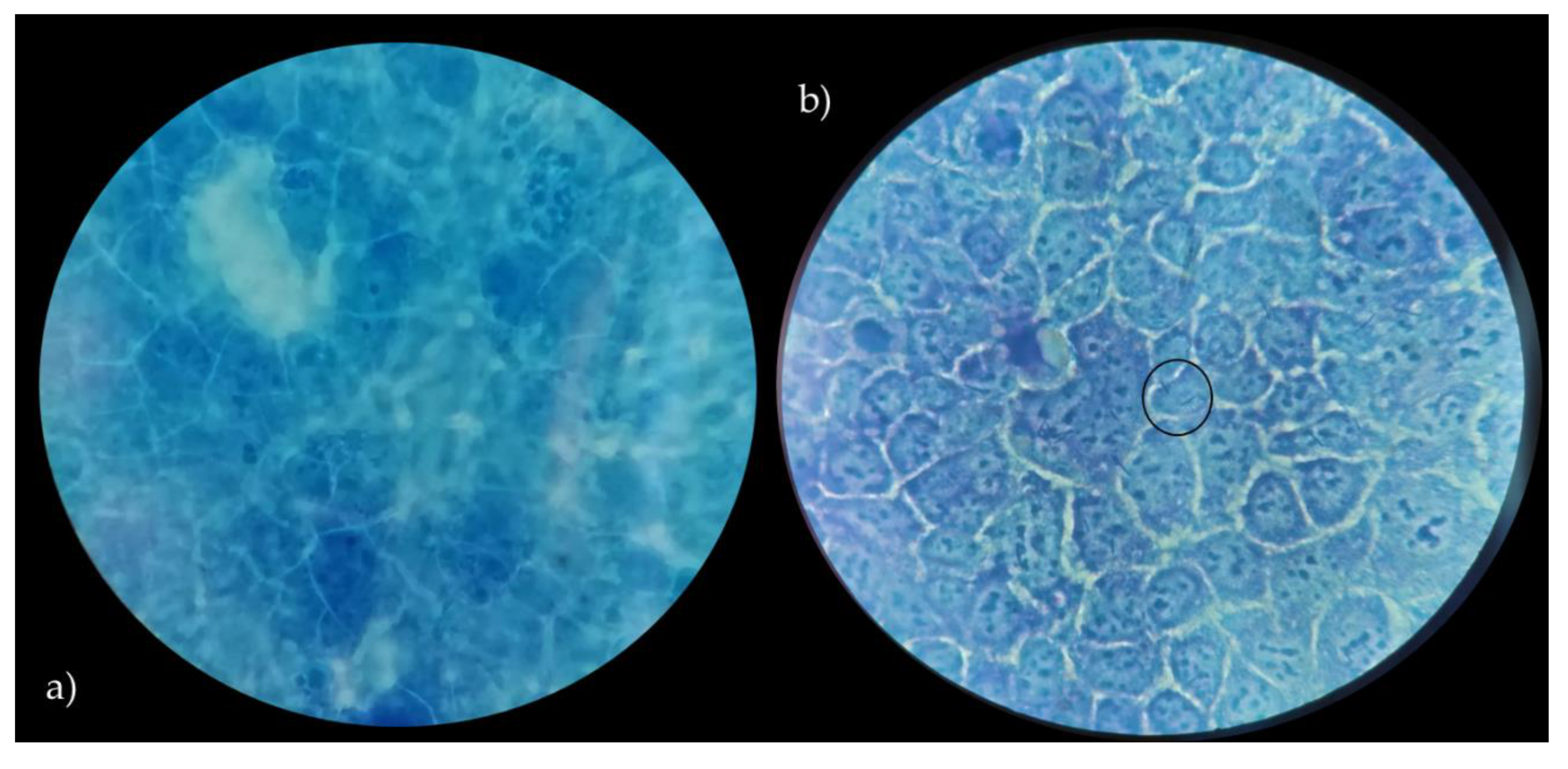
3.4. Adhesion to Caco-2 Cells
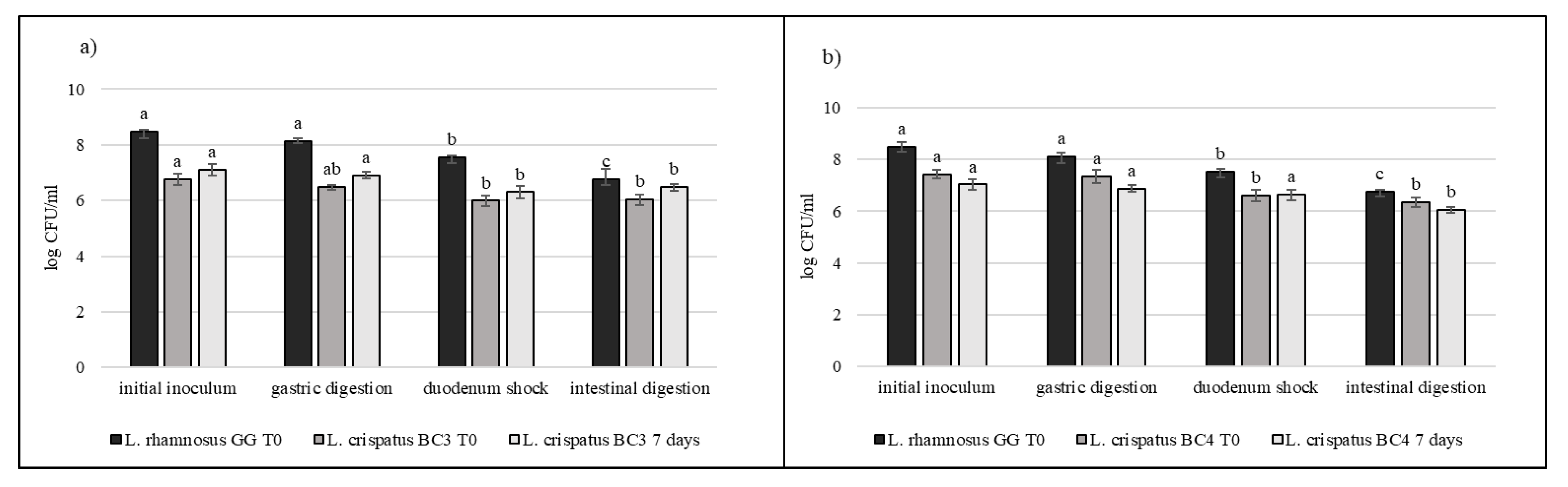
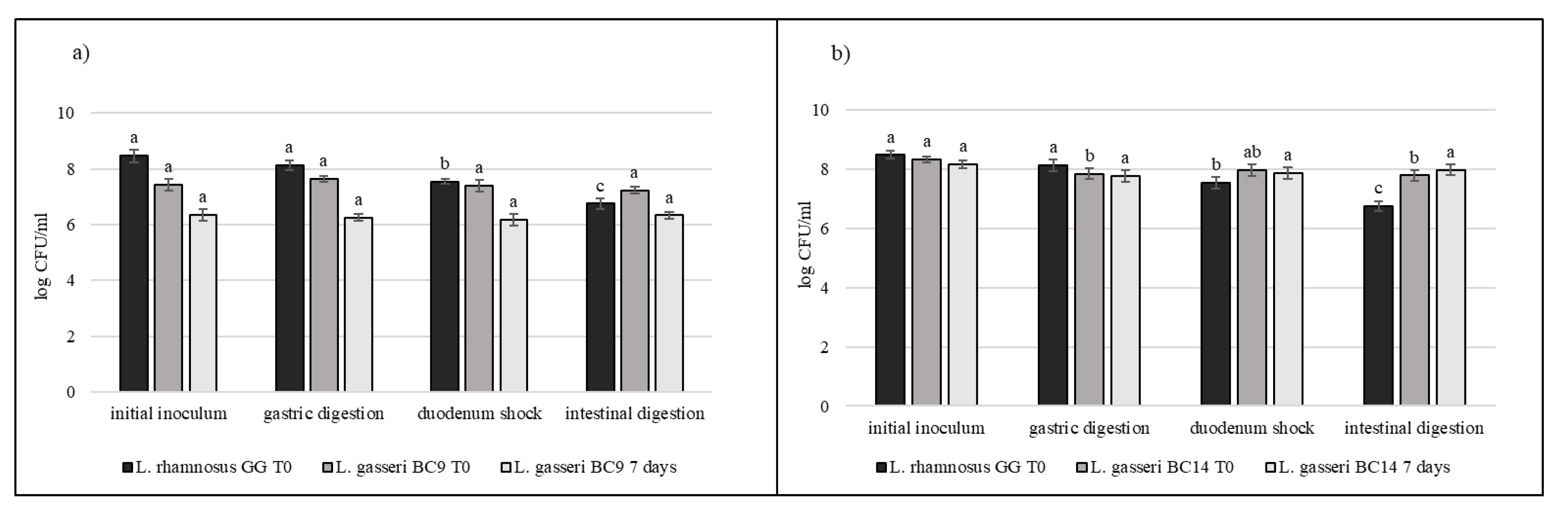
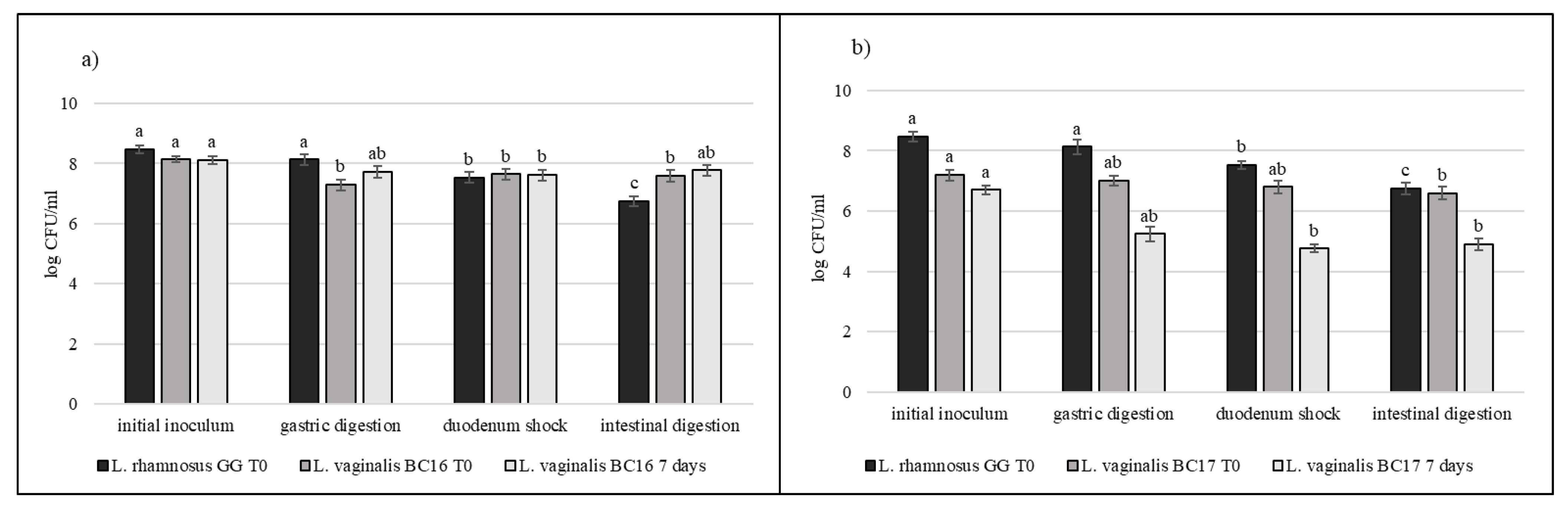
3.5. Strain Tolerance to Simulated Digestion Process in Milk
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Younes, J.A.; Lievens, E.; Hummelen, R.; van der Westen, R.; Reid, G.; Petrova, M.I. Women and their microbes: The unexpected friendship. Trends Microbiol. 2018, 26, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Vitali, B.; Cruciani, F.; Baldassarre, M.E.; Capursi, T.; Spisni, E.; Valerii, M.C.; Candela, M.; Turroni, S.; Brigidi, P. Dietary supplementation with probiotics during late pregnancy: Outcome on vaginal microbiota and cytokine secretion. BMC Microbiol. 2012, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirmonsef, P.; Hotton, A.L.; Gilbert, D.; Burgad, D.; Landay, A.; Weber, K.M.; Cohen, M.; Ravel, J.; Spear, G.T. Free glycogen in vaginal fluids is associated with Lactobacillus colonization and low vaginal pH. PLoS ONE 2014, 9, e102467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, G.; Younes, J.A.; Van Der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Silva, J.; Teixeira, P. The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 2014, 289, 479–489. [Google Scholar] [CrossRef]
- Parolin, C.; Marangoni, A.; Laghi, L.; Foschi, C.; Palomino, R.A.Ñ.; Calonghi, N.; Cevenini, R.; Vitali, B. Isolation of vaginal lactobacilli and characterization of anti-candida activity. PLoS ONE 2015, 10, e0131220. [Google Scholar] [CrossRef] [PubMed]
- Strus, M.; Chmielarczyk, A.; Kochan, P.; Adamski, P.; Chełmicki, Z.; Chełmicki, A.; Pałucha, A.; Heczko, P.B. Studies on the effects of probiotic Lactobacillus mixture given orally on vaginal and rectal colonization and on parameters of vaginal health in women with intermediate vaginal flora. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 163, 210–215. [Google Scholar] [CrossRef]
- Vujic, G.; Jajac Knez, A.; Despot Stefanovic, V.; Kuzmic Vrbanovic, V. Efficacy of orally applied probiotic capsules for bacterial vaginosis and other vaginal infections: A double-blind, randomized, placebo-controlled study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 12, 236. [Google Scholar] [CrossRef]
- Heczko, P.B.; Tomusiak, A.; Adamski, P.; Jakimiuk, A.J.; Stefanski, G.; Mikolajczyk-Cichonska, A.; Suda-Szczurek, M.; Strus, M. Supplementation of standard antibiotic therapy with oral probiotics for bacterial vaginosis and aerobic vaginitis: A randomised, double-blind, placebocontrolled trial. BMC Womens Health 2015, 15, 115. [Google Scholar] [CrossRef] [Green Version]
- Calonghi, N.; Parolin, C.; Sartor, G.; Verardi, L.; Giordani, B.; Frisco, G.; Marangoni, A.; Vitali, B. Interaction of vaginal Lactobacillus strains with HeLa cells plasma membrane. Benef. Microbes 2017, 8, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Nardini, P.; Nãhui Palomino, R.A.; Parolin, C.; Laghi, L.; Foschi, C.; Cevenini, R.; Vitali, B.; Marangoni, A. Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Parolin, C.; Frisco, G.; Foschi, C.; Giordani, B.; Salvo, M.; Vitali, B.; Marangoni, A.; Calonghi, N. Lactobacillus crispatus BC5 interferes with Chlamydia trachomatis infectivity through integrin modulation in cervical cells. Front. Microbiol. 2018, 16, 40–51. [Google Scholar] [CrossRef]
- Foschi, C.; Salvo, M.; Cevenini, R.; Parolin, C.; Vitali, B.; Marangoni, A. Vaginal lactobacilli reduce Neisseria gonorrhoeae viability through multiple strategies: An in vitro study. Front. Cell. Infect. Microbiol. 2017, 7, 502. [Google Scholar] [CrossRef]
- Marziali, G.; Foschi, C.; Parolin, C.; Vitali, B.; Marangoni, A. In-vitro effect of vaginal lactobacilli against group B Streptococcus. Microb. Pathog. 2019, 136, 103692. [Google Scholar] [CrossRef] [PubMed]
- Ñahui Palomino, R.A.; Zicari, S.; Vanpouille, C.; Vitali, B.; Margolis, L. Vaginal lactobacillus inhibits HIV-1 replication in human tissues ex vivo. Front. Microbiol. 2017, 8, 906. [Google Scholar] [CrossRef] [Green Version]
- Ñahui Palomino, R.A.; Vanpouille, C.; Laghi, L.; Parolin, C.; Melikov, K.; Backlund, P.; Vitali, B.; Margolis, L. Extracellular vesicles from symbiotic vaginal lactobacilli inhibit HIV-1 infection of human tissues. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Siroli, L.; Patrignani, F.; Serrazanetti, D.I.; Parolin, C.; Palomino, R.A.Ñ.; Vitali, B.; Lanciotti, R. Determination of antibacterial and technological properties of vaginal lactobacilli for their potential application in dairy products. Front. Microbiol. 2017, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; Siragusa, S.; Caputo, L.; Ragni, A.; Burzigotti, R.; Gobbetti, M. Survival and persistence of Lactobacillus plantarum 4.1 and Lactobacillus reuteri 3S7 in the gastrointestinal tract of pigs. Vet. Microbiol. 2007. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Reinheimer, J.A. Lactic acid starter and probiotic bacteria: A comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Res. Int. 2003, 64, 269–275. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Mathara, J.M.; Schillinger, U.; Guigas, C.; Franz, C.; Kutima, P.M.; Mbugua, S.K.; Shin, H.K.; Holzapfel, W.H. Functional characteristics of Lactobacillus spp. from traditional Maasai fermented milk products in Kenya. Int. J. Food Microbiol. 2008, 126, 57–64. [Google Scholar] [CrossRef]
- Lea, T. Caco-2 cell line. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer Nature Switzerland AG: Cham, Switzerland, 2015; ISBN 9783319161044. [Google Scholar]
- Vinderola, G.; Céspedes, M.; Mateolli, D.; Cárdenas, P.; Lescano, M.; Aimaretti, N.; Reinheimer, J. Changes in gastric resistance of Lactobacillus casei in flavoured commercial fermented milks during refrigerated storage. Int. J. Dairy Technol. 2011, 64, 269–275. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, W147–W153. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.K.; Liong, M.T. Effect of prebiotics on viability and growth characteristics of probiotics in soymilk. J. Sci. Food Agric. 2010, 90, 267–275. [Google Scholar] [CrossRef]
- Pokusaeva, K.; O’Connell-Motherway, M.; Zomer, A.; MacSharry, J.; Fitzgerald, G.F.; van Sinderen, D. Cellodextrin utilization by Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2011, 77, 1681–1690. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.L.; Gibson, G.R.; Rastall, R.A. Influence of disaccharide structure on prebiotic selectivity in vitro. J. Agric. Food Chem. 2005, 53. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Mikš, M.H. Uncovering carbohydrate metabolism through a genotype-phenotype association study of 56 lactic acid bacteria genomes. Appl. Microbiol. Biotechnol. 2019, 103, 3135–3152. [Google Scholar] [CrossRef] [Green Version]
- Perpetuini, G.; Tittarelli, F.; Mattarelli, P.; Modesto, M.; Cilli, E.; Suzzi, G.; Tofalo, R. Intraspecies polymorphisms of Kluyveromyces marxianus strains from Yaghnob valley. FEMS Microbiol. Lett. 2018, 365, fny028. [Google Scholar] [CrossRef]
- Campaniello, D.; Speranza, B.; Petruzzi, L.; Bevilacqua, A.; Corbo, M.R. How to routinely assess transition, adhesion and survival of probiotics into the gut: A case study on propionibacteria. Int. J. Food Sci. Technol. 2018, 53, 484–490. [Google Scholar] [CrossRef]
- Schillinger, U.; Guigas, C.; Holzapfel, W.H. In vitro adherence and other properties of lactobacilli used in probiotic yoghurt-like products. Int. Dairy J. 2005, 15, 1289–1297. [Google Scholar] [CrossRef]
- Tabanelli, G.; Patrignani, F.; Vinderola, G.; Reinheimer, J.A.; Gardini, F.; Lanciotti, R. Effect of sub-lethal high pressure homogenization treatments on the invitro functional and biological properties of lactic acid bacteria. LWT Food Sci. Technol. 2013, 53, 580–586. [Google Scholar] [CrossRef]
- Abdulla, A. Adhesion, Autoaggregation and Hydrophobicity of Six Lactobacillus Strains. Br. Microbiol. Res. J. 2014, 381–391. [Google Scholar] [CrossRef]
- Boris, S.; Suárez, J.E.; Vázquez, F.; Barbés, C. Adherence of human vaginal lactobacilli to vaginal epithelial cells and interaction with uropathogens. Infect. Immun. 1998. [Google Scholar] [CrossRef] [Green Version]
- Martín, R.; Olivares, M.; Marín, M.L.; Fernández, L.; Xaus, J.; Rodríguez, J.M. Probiotic potential of 3 lactobacilli strains isolated from breast milk. J. Hum. Lact. 2005, 21, 8–17. [Google Scholar] [CrossRef]
- Noriega, L.; Cuevas, I.; Margolles, A.; de los Reyes-Gavilán, C.G. Deconjugation and bile salts hydrolase activity by Bifidobacterium strains with acquired resistance to bile. Int. Dairy J. 2006, 16, 850–855. [Google Scholar] [CrossRef]
- Shehata, M.G.; El Sohaimy, S.A.; El-Sahn, M.A.; Youssef, M.M. Screening of isolated potential probiotic lactic acid bacteria for cholesterol lowering property and bile salt hydrolase activity. Ann. Agric. Sci. 2016, 61, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Hill, C.; Gahan, C.G.M. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef] [Green Version]
- Candela, M.; Seibold, G.; Vitali, B.; Lachenmaier, S.; Eikmanns, B.J.; Brigidi, P. Real-time PCR quantification of bacterial adhesion to Caco-2 cells: Competition between bifidobacteria and enteropathogens. Res. Microbiol. 2005, 156, 887–895. [Google Scholar] [CrossRef]
- Kim, J.H.; Baik, S.H. Probiotic properties of Lactobacillus strains with high cinnamoyl esterase activity isolated from jeot-gal, a high-salt fermented seafood. Ann. Microbiol. 2019, 69, 407–417. [Google Scholar] [CrossRef]
- Patrignani, F.; Parolin, C.; D’Alessandro, M.; Siroli, L.; Vitali, B.; Lanciotti, R. Evaluation of the fate of Lactobacillus crispatus BC4, carried in Squacquerone cheese, throughout the simulator of the human intestinal microbial ecosystem (SHIME). Food Res. Int. 2020, 137, 109580. [Google Scholar] [CrossRef]
- Chung, H.J.; Bang, W.; Drake, M.A. Stress response of Escherichia coli. Compr. Rev. Food Sci. Food Saf. 2006, 5, 52–64. [Google Scholar] [CrossRef]
- Patrignani, F.; Siroli, L.; Parolin, C.; Serrazanetti, D.I.; Vitali, B.; Lanciotti, R. Use of Lactobacillus crispatus to produce a probiotic cheese as potential gender food for preventing gynaecological infections. PLoS ONE 2019. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Species | Isolation Source | Collection |
|---|---|---|---|
| BC1 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC3 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC4 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC5 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC6 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC7 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC8 | Lactobacillus crispatus | Female genital tract [13] | FABIT |
| BC9 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC10 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC11 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC12 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC13 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC14 | Lactobacillus gasseri | Female genital tract [13] | FABIT |
| BC16 | Limosilactobacillus vaginalis | Female genital tract [13] | FABIT |
| BC17 | Limosilactobacillus vaginalis | Female genital tract [13] | FABIT |
| Strains | Adhesion (Lactobacilli Cells/Caco-2 Cell) |
|---|---|
| Lactobacillus crispatus BC1 | 0.63 a (±0.21) |
| Lactobacillus crispatus BC3 | 0.45 ad (±0.19) |
| Lactobacillus crispatus BC4 | 0.24 b (±0.09) |
| Lactobacillus crispatus BC5 | 0.43 ad (±0.17) |
| Lactobacillus crispatus BC6 | 0.74 a (±0.21) |
| Lactobacillus crispatus BC8 | 5.14 c (±2.29) |
| Lactobacillus gasseri BC9 | 0.26 d (±0.10) |
| Lactobacillus gasseri BC11 | 0.27 d (±0.11) |
| Lactobacillus gasseri BC12 | 0.75 a (±0.23) |
| Lactobacillus gasseri BC14 | 0.15 e (±0.06) |
| Limosilactobacillus vaginalis BC16 | 0.34 ad (±0.16) |
| Limosilactobacillus vaginalis BC17 | 2.32 f (±1.17) |
| Lacticaseibacillus rhamnosus GG | 8.90 g (±1.24) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Alessandro, M.; Parolin, C.; Bukvicki, D.; Siroli, L.; Vitali, B.; De Angelis, M.; Lanciotti, R.; Patrignani, F. Probiotic and Metabolic Characterization of Vaginal Lactobacilli for a Potential Use in Functional Foods. Microorganisms 2021, 9, 833. https://doi.org/10.3390/microorganisms9040833
D’Alessandro M, Parolin C, Bukvicki D, Siroli L, Vitali B, De Angelis M, Lanciotti R, Patrignani F. Probiotic and Metabolic Characterization of Vaginal Lactobacilli for a Potential Use in Functional Foods. Microorganisms. 2021; 9(4):833. https://doi.org/10.3390/microorganisms9040833
Chicago/Turabian StyleD’Alessandro, Margherita, Carola Parolin, Danka Bukvicki, Lorenzo Siroli, Beatrice Vitali, Maria De Angelis, Rosalba Lanciotti, and Francesca Patrignani. 2021. "Probiotic and Metabolic Characterization of Vaginal Lactobacilli for a Potential Use in Functional Foods" Microorganisms 9, no. 4: 833. https://doi.org/10.3390/microorganisms9040833
APA StyleD’Alessandro, M., Parolin, C., Bukvicki, D., Siroli, L., Vitali, B., De Angelis, M., Lanciotti, R., & Patrignani, F. (2021). Probiotic and Metabolic Characterization of Vaginal Lactobacilli for a Potential Use in Functional Foods. Microorganisms, 9(4), 833. https://doi.org/10.3390/microorganisms9040833