
Ambroxol Treatment Suppresses the Proliferation of Chlamydia pneumoniae in Murine Lungs


 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of C. pneumoniae
2.2. Infection of Mice and Preparation of Lung Tissues
2.3. Culturing of C. pneumoniae from Murine Lungs
2.4. Total RNA Extraction and cDNA Synthesis
2.5. qPCR Amplification of Ido1, Ido2, Il12, Il23, Il17a, Il17f, Ifng, Sftpa, Sftpb, Sftpc, and Sftpd
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Western Blotting Analysis of ERK
2.8. Apoptosis Assay and Flow Cytometry
2.9. In Vitro Effects of SP-A and SP-D on C. pneumoniae Proliferation and Attachment
2.10. Statistical Analysis
3. Results
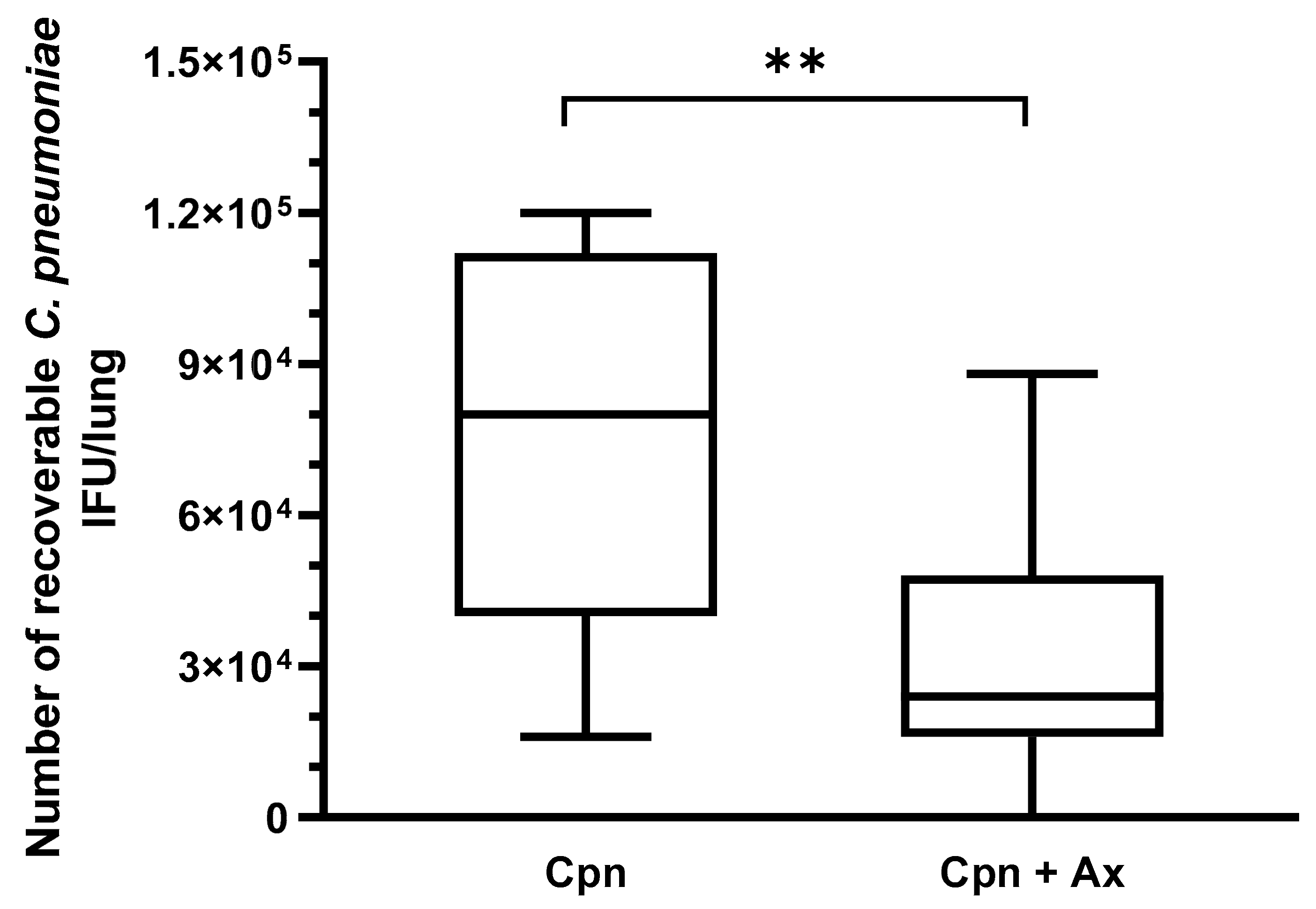
3.1. Ax Treatment at Elevated Concentration Suppressed C. pneumoniae Proliferation in Mouse Lungs
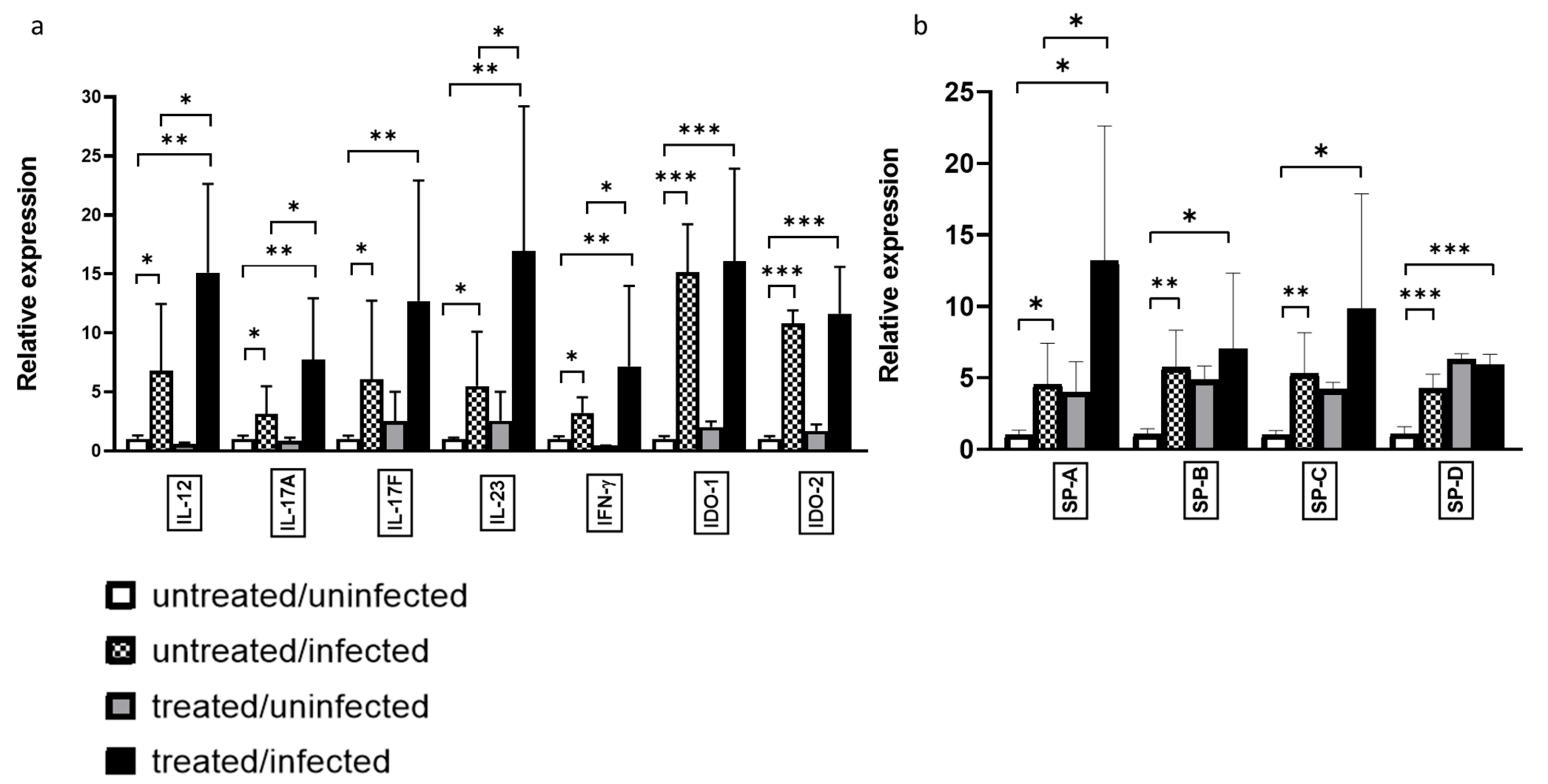
3.2. Ax Treatment Altered the Gene Expression and Protein Level of IFN-γ in the Lungs of C. pneumoniae-Infected Mice
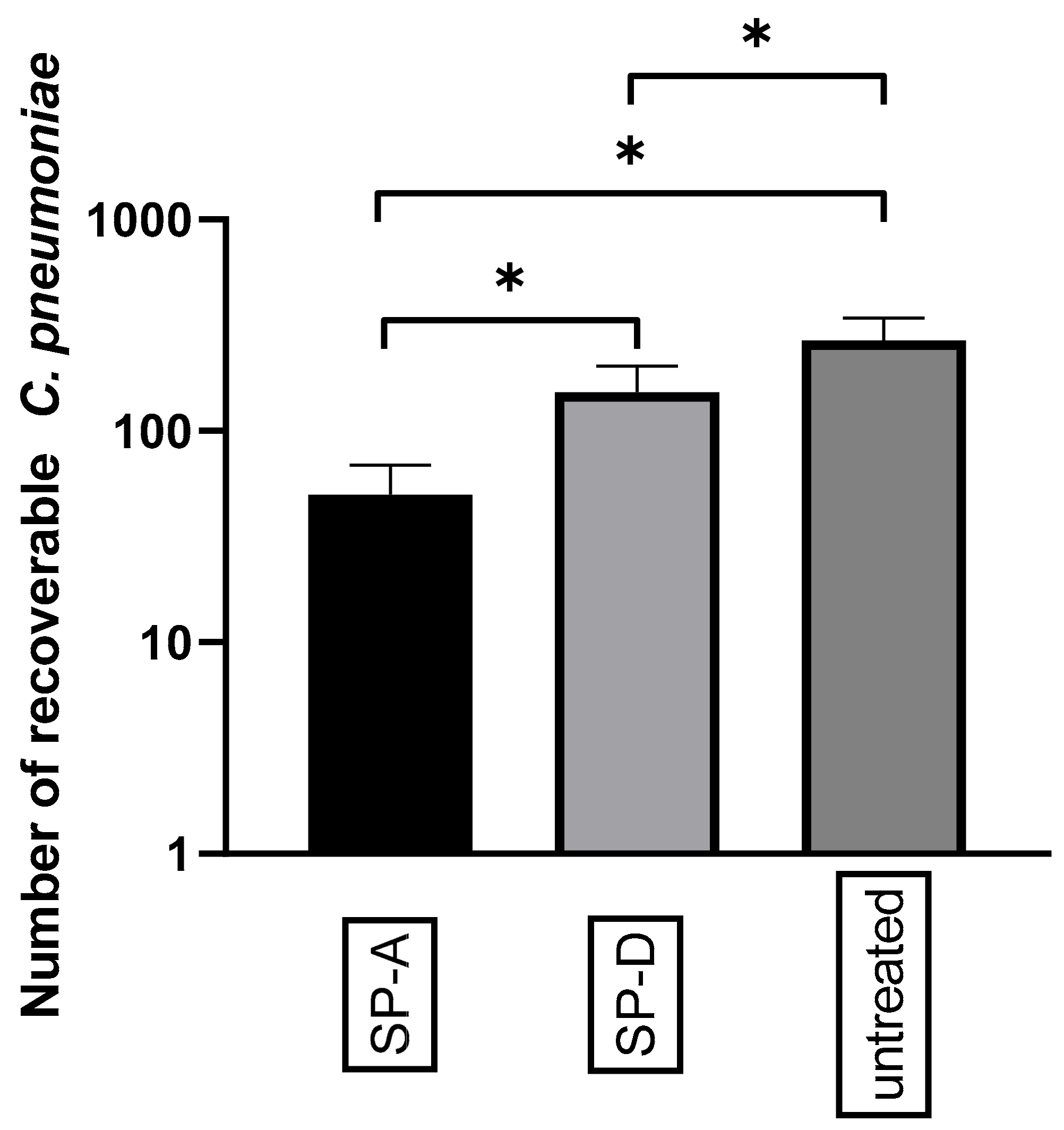
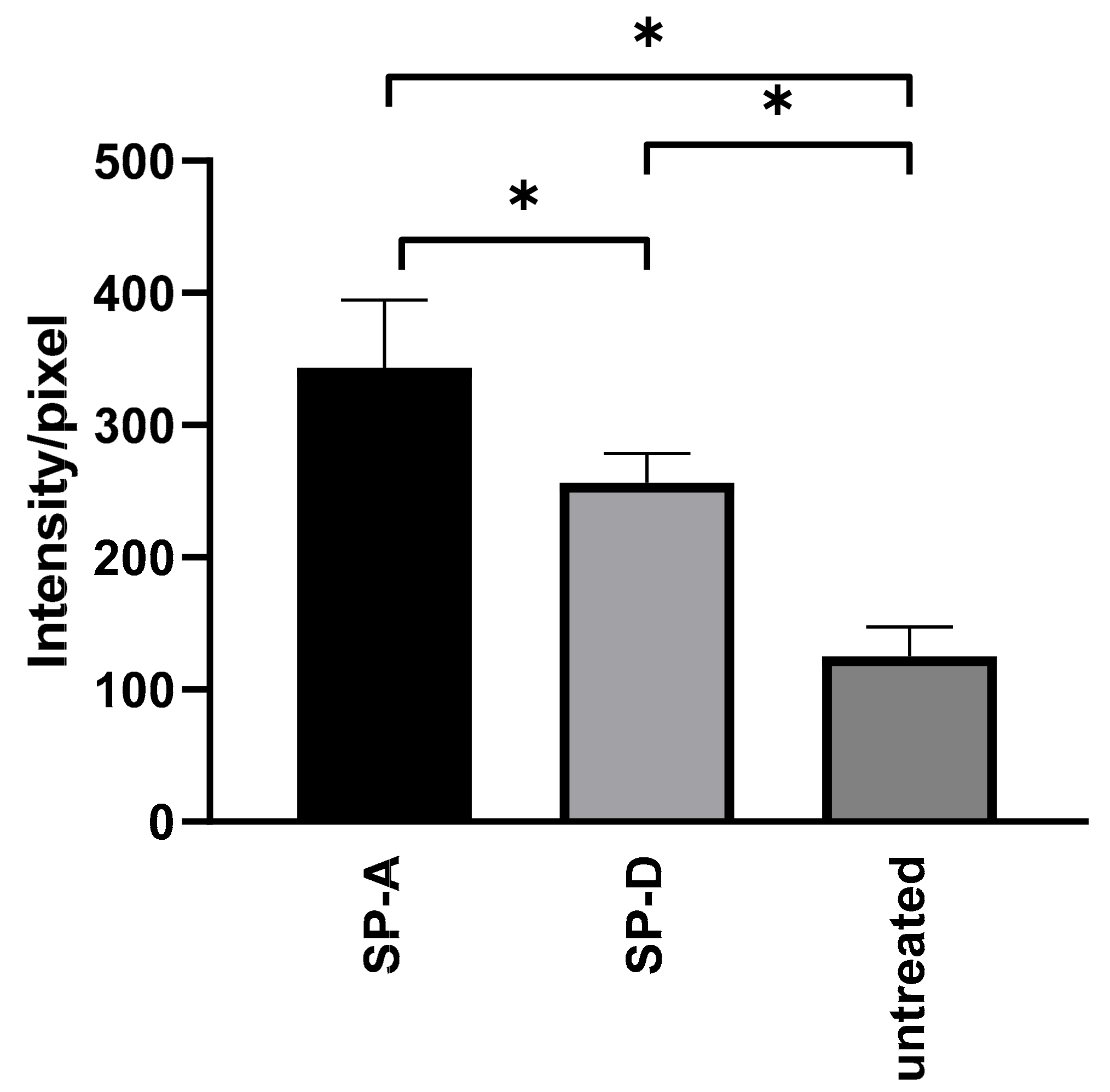
3.3. SP Treatment Increased the Attachment of C. pneumoniae to Macrophages and Decreased Bacterial Proliferation
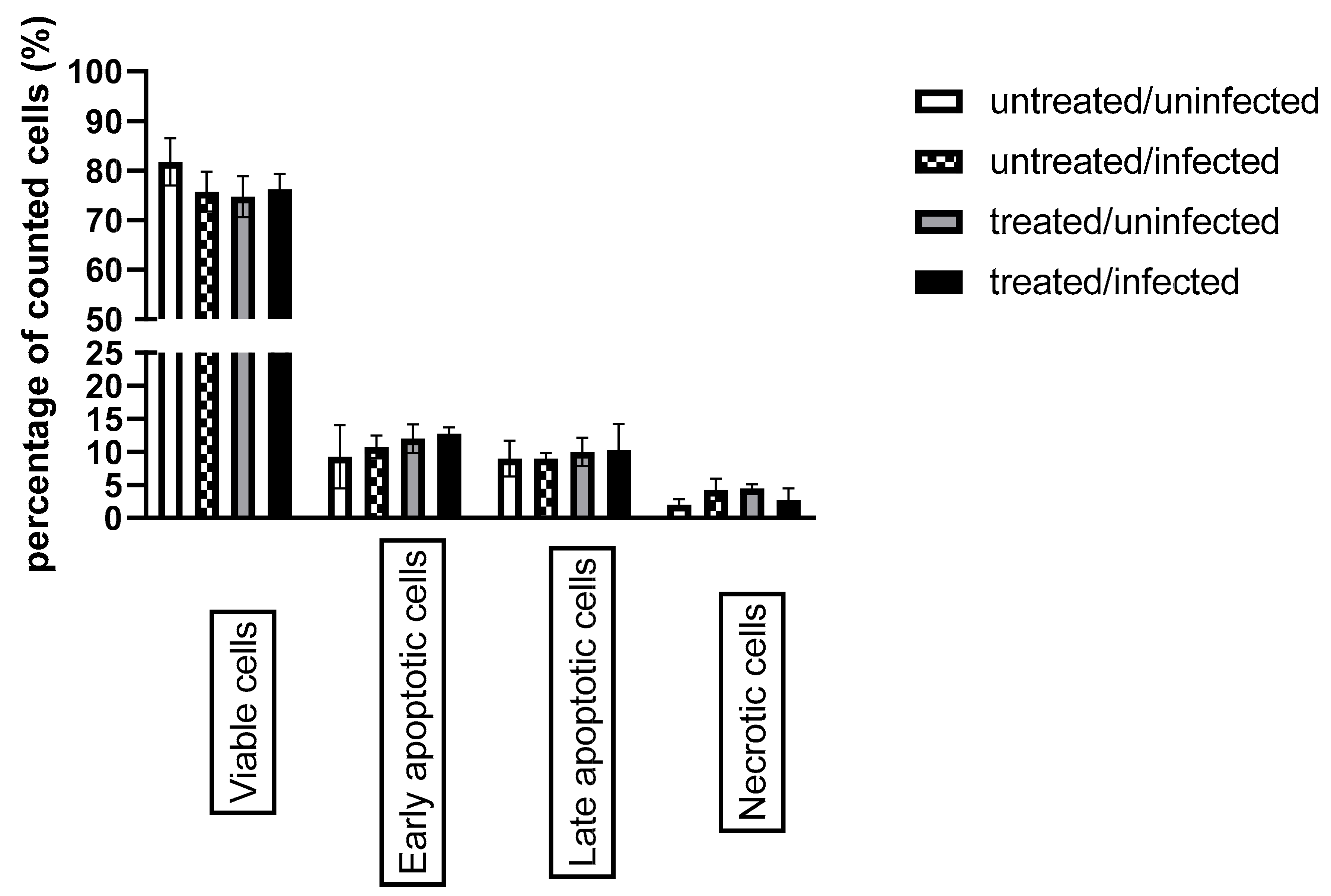
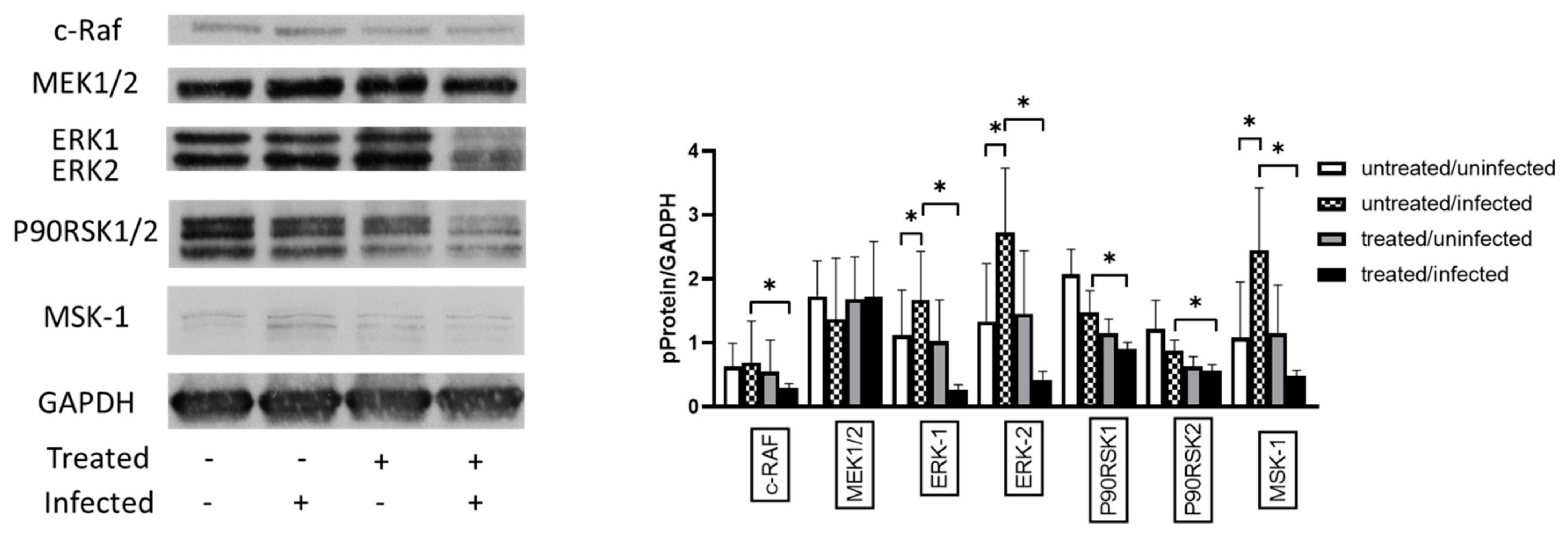
3.4. Ax Treatment Did Not Induce Apoptosis via the Caspase-Dependent Pathway But Decreased ERK 1/2 Activation in C. pneumoniae-Infected Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Germouty, J.; Jirou-Najou, J.L. Clinical Efficacy of Ambroxol in the Treatment of Bronchial Stasis. Clinical Trial in 120 Patients at Two Different Doses. Respiration 1987, 51 (Suppl. 1), 37–41. [Google Scholar] [CrossRef] [PubMed]
- Cazan, D.; Klimek, L.; Sperl, A.; Plomer, M.; Kölsch, S. Safety of Ambroxol in the Treatment of Airway Diseases in Adult Patients. Expert Opin. Drug Saf. 2018, 17, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A.H.V. Glucocerebrosidase and Parkinson Disease: Molecular, Clinical, and Therapeutic Implications. Neuroscientist 2018, 24, 540–559. [Google Scholar] [CrossRef] [PubMed]
- Gillissen, A.; Schärling, B.; Jaworska, M.; Bartling, A.; Rasche, K.; Schultze-Werninghaus, G. Oxidant Scavenger Function of Ambroxol in Vitro: A Comparison with N-Acetylcysteine. Res. Exp. Med. 1997, 196, 389–398. [Google Scholar] [CrossRef]
- Aihara, M.; Dobashi, K.; Akiyama, M.; Naruse, I.; Nakazawa, T.; Mori, M. Effects of N-Acetylcysteine and Ambroxol on the Production of IL-12 and IL-10 in Human Alveolar Macrophages. Respiration 2000, 67, 662–671. [Google Scholar] [CrossRef]
- He, W.; Xiao, W.; Zhang, X.; Sun, Y.; Chen, Y.; Chen, Q.; Fang, X.; Du, S.; Sha, X. Pulmonary-Affinity Paclitaxel Polymer Micelles in Response to Biological Functions of Ambroxol Enhance Therapeutic Effect on Lung Cancer. Int. J. Nanomed. 2020, 15, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Jiang, J.; Ren, Q.; Jia, Y.; Shen, J.; Shen, H.; Lin, X.; Lu, H.; Xie, Q. Ambroxol Inhalation Ameliorates LPS-Induced Airway Inflammation and Mucus Secretion through the Extracellular Signal-Regulated Kinase 1/2 Signaling Pathway. Eur. J. Pharmacol. 2016, 775, 138–148. [Google Scholar] [CrossRef]
- de Mey, C.; Koelsch, S.; Richter, E.; Pohlmann, T.; Sousa, R. Efficacy and Safety of Ambroxol Lozenges in the Treatment of Acute Uncomplicated Sore Throat - a Pooled Analysis. Drug Res. 2016, 66, 384–392. [Google Scholar] [CrossRef]
- Yang, B.; Yao, D.F.; Ohuchi, M.; Ide, M.; Yano, M.; Okumura, Y.; Kido, H. Ambroxol Suppresses Influenza-Virus Proliferation in the Mouse Airway by Increasing Antiviral Factor Levels. Eur. Respir. J. 2002, 19, 952–958. [Google Scholar] [CrossRef]
- Kun, D.; Xiang-lin, C.; Ming, Z.; Qi, L. Chlamydia Inhibit Host Cell Apoptosis by Inducing Bag-1 via the MAPK/ERK Survival Pathway. Apoptosis 2013, 18, 1083–1092. [Google Scholar] [CrossRef]
- Su, H.; McClarty, G.; Dong, F.; Hatch, G.M.; Pan, Z.K.; Zhong, G. Activation of Raf/MEK/ERK/CPLA2 Signaling Pathway Is Essential for Chlamydial Acquisition of Host Glycerophospholipids. J. Biol. Chem. 2004, 279, 9409–9416. [Google Scholar] [CrossRef] [Green Version]
- Oberley, R.E.; Ault, K.A.; Neff, T.L.; Khubchandani, K.R.; Crouch, E.C.; Snyder, J.M. Surfactant Proteins A and D Enhance the Phagocytosis of Chlamydia into THP-1 Cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L296–L306. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Rudel, T. Apoptosis Resistance in Chlamydia-Infected Cells: A Fate Worse than Death? FEMS Immunol. Med. Microbiol. 2009, 55, 154–161. [Google Scholar] [CrossRef]
- Choroszy-Król, I.; Frej-Mądrzak, M.; Hober, M.; Sarowska, J.; Jama-Kmiecik, A. Infections Caused by Chlamydophila Pneumoniae. Adv. Clin. Exp. Med. 2014, 23, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Petrovay, F.; Heltai, K.; Kis, Z.; Treso, B.; Gonczol, E.; Burian, K.; Endresz, V.; Valyi-Nagy, I. Chronic Infections and Histamine, CRP and IL-6 Levels after Percutaneous Transluminal Coronary Angioplasty. Inflamm. Res. 2007, 56, 362–367. [Google Scholar] [CrossRef]
- Burian, K.; Berencsi, K.; Endresz, V.; Gyulai, Z.; Valyi-Nagy, T.; Valyi-Nagy, I.; Bakay, M.; Geng, Y.; Virok, D.; Kari, L.; et al. Chlamydia Pneumoniae Exacerbates Aortic Inflammatory Foci Caused by Murine Cytomegalovirus Infection in Normocholesterolemic Mice. Clin. Diagn. Lab. Immunol. 2001, 8, 1263–1266. [Google Scholar] [CrossRef] [Green Version]
- Kókai, D.; Mosolygó, T.; Virók, D.P.; Endrész, V.; Burián, K. N-Acetyl-Cysteine Increases the Replication of Chlamydia Pneumoniae and Prolongs the Clearance of the Pathogen from Mice. J. Med. Microbiol. 2018, 67, 702–708. [Google Scholar] [CrossRef]
- Burián, K.; Hegyesi, H.; Buzás, E.; Endrész, V.; Kis, Z.; Falus, A.; Gönczöl, E. Chlamydophila (Chlamydia) Pneumoniae Induces Histidine Decarboxylase Production in the Mouse Lung. Immunol. Lett. 2003, 89, 229–236. [Google Scholar] [CrossRef]
- Balogh, E.P.; Faludi, I.; Virók, D.P.; Endrész, V.; Burián, K. Chlamydophila Pneumoniae Induces Production of the Defensin-like MIG/CXCL9, Which Has in Vitro Antichlamydial Activity. Int. J. Med. Microbiol. 2011, 301, 252–259. [Google Scholar] [CrossRef]
- Caldwell, H.D.; Kromhout, J.; Schachter, J. Purification and Partial Characterization of the Major Outer Membrane Protein of Chlamydia Trachomatis. Infect. Immun. 1981, 31, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Mosolygó, T.; Szabó, Á.M.; Balogh, E.P.; Faludi, I.; Virók, D.P.; Endrész, V.; Samu, A.; Krenács, T.; Burián, K. Protection Promoted by PGP3 or PGP4 against Chlamydia Muridarum Is Mediated by CD4+ Cells in C57BL/6N Mice. Vaccine 2014, 32, 5228–5233. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gáspár, R.; Pipicz, M.; Hawchar, F.; Kovács, D.; Djirackor, L.; Görbe, A.; Varga, Z.V.; Kiricsi, M.; Petrovski, G.; Gácser, A.; et al. The Cytoprotective Effect of Biglycan Core Protein Involves Toll-like Receptor 4 Signaling in Cardiomyocytes. J. Mol. Cell. Cardiol. 2016, 99, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Szabó, M.R.; Gáspár, R.; Pipicz, M.; Zsindely, N.; Diószegi, P.; Sárközy, M.; Bodai, L.; Csont, T. Hypercholesterolemia Interferes with Induction of MiR-125b-1-3p in Preconditioned Hearts. Int. J. Mol. Sci. 2020, 21, 3744. [Google Scholar] [CrossRef]
- Virok, D.P.; Raffai, T.; Kókai, D.; Paróczai, D.; Bogdanov, A.; Veres, G.; Vécsei, L.; Poliska, S.; Tiszlavicz, L.; Somogyvári, F.; et al. Indoleamine 2,3-Dioxygenase Activity in Chlamydia Muridarum and Chlamydia Pneumoniae Infected Mouse Lung Tissues. Front. Cell. Infect. Microbiol. 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Roshick, C.; Wood, H.; Caldwell, H.D.; McClarty, G. Comparison of Gamma Interferon-Mediated Antichlamydial Defense Mechanisms in Human and Mouse Cells. Infect. Immun. 2006, 74, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Xu, M.; Wang, Y.; Wang, T.; Wu, N.; Zheng, W.; Duan, H. Air Particulate Matter Pollution and Circulating Surfactant Protein: A Systemic Review and Meta-Analysis. Chemosphere 2021, 272, 129564. [Google Scholar] [CrossRef]
- Johansson, J.; Curstedt, T. Synthetic Surfactants with SP-B and SP-C Analogues to Enable Worldwide Treatment of Neonatal Respiratory Distress Syndrome and Other Lung Diseases. J. Intern. Med. 2019, 285, 165–186. [Google Scholar] [CrossRef] [Green Version]
- Oberley, R.E.; Goss, K.L.; Ault, K.A.; Crouch, E.C.; Snyder, J.M. Surfactant Protein D Is Present in the Human Female Reproductive Tract and Inhibits Chlamydia Trachomatis Infection. Mol. Hum. Reprod. 2004, 10, 861–870. [Google Scholar] [CrossRef]
- Galle, J.N.; Fechtner, T.; Eierhoff, T.; Römer, W.; Hegemann, J.H. A Chlamydia Pneumoniae Adhesin Induces Phosphatidylserine Exposure on Host Cells. Nat. Commun. 2019, 10, 4644. [Google Scholar] [CrossRef] [Green Version]
- McCOY, C.E.; Campbell, D.G.; Deak, M.; Bloomberg, G.B.; Arthur, J.S.C. MSK1 Activity Is Controlled by Multiple Phosphorylation Sites. Biochem. J. 2005, 387, 507–517. [Google Scholar] [CrossRef]
- Seifart, C.; Clostermann, U.; Seifart, U.; Müller, B.; Vogelmeier, C.; von Wichert, P.; Fehrenbach, H. Cell-Specific Modulation of Surfactant Proteins by Ambroxol Treatment. Toxicol. Appl. Pharmacol. 2005, 203, 27–35. [Google Scholar] [CrossRef]
- Fois, G.; Hobi, N.; Felder, E.; Ziegler, A.; Miklavc, P.; Walther, P.; Radermacher, P.; Haller, T.; Dietl, P. A New Role for an Old Drug: Ambroxol Triggers Lysosomal Exocytosis via PH-Dependent Ca2+ Release from Acidic Ca2+ Stores. Cell Calcium 2015, 58, 628–637. [Google Scholar] [CrossRef]
- Weiser, T. Comparison of the Effects of Four Na+ Channel Analgesics on TTX-Resistant Na+ Currents in Rat Sensory Neurons and Recombinant Nav1.2 Channels. Neurosci. Lett. 2006, 395, 179–184. [Google Scholar] [CrossRef]
- McNeill, A.; Magalhaes, J.; Shen, C.; Chau, K.-Y.; Hughes, D.; Mehta, A.; Foltynie, T.; Cooper, J.M.; Abramov, A.Y.; Gegg, M.; et al. Ambroxol Improves Lysosomal Biochemistry in Glucocerebrosidase Mutation-Linked Parkinson Disease Cells. Brain 2014, 137, 1481–1495. [Google Scholar] [CrossRef] [Green Version]
- Deretic, V.; Timmins, G.S. Enhancement of Lung Levels of Antibiotics by Ambroxol and Bromhexine. Expert Opin. Drug Metab. Toxicol. 2019, 15, 213–218. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Y.; Huang, X.; Yu, C.; Yang, Y.; Sun, S. Ambroxol Hydrochloride Combined with Fluconazole Reverses the Resistance of Candida Albicans to Fluconazole. Front. Cell. Infect. Microbiol. 2017, 7, 124. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Y.; Yu, J.; Ai, Q.; Li, J.; Peng, N.; Song, S.; He, Y.; Wang, Z. Synergy of Ambroxol with Vancomycin in Elimination of Catheter-Related Staphylococcus Epidermidis Biofilm in Vitro and in Vivo. J. Infect. Chemother. 2015, 21, 808–815. [Google Scholar] [CrossRef]
- Yamaya, M.; Nishimura, H.; Nadine, L.K.; Ota, C.; Kubo, H.; Nagatomi, R. Ambroxol Inhibits Rhinovirus Infection in Primary Cultures of Human Tracheal Epithelial Cells. Arch. Pharm. Res. 2014, 37, 520–529. [Google Scholar] [CrossRef]
- Cataldi, M.; Sblendorio, V.; Leo, A.; Piazza, O. Biofilm-Dependent Airway Infections: A Role for Ambroxol? Pulm. Pharmacol. Ther. 2014, 28, 98–108. [Google Scholar] [CrossRef]
- Mosolygó, T.; Korcsik, J.; Balogh, E.P.; Faludi, I.; Virók, D.P.; Endrész, V.; Burián, K. Chlamydophila Pneumoniae Re-Infection Triggers the Production of IL-17A and IL-17E, Important Regulators of Airway Inflammation. Inflamm. Res. 2013, 62, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Neutrophils, Interleukin-17A and Lung Disease | European Respiratory Society. Available online: https://erj.ersjournals.com/content/25/1/159.abstract?ijkey=26df422c417132c793da0a7c0d7a6f922caad19d&keytype2=tf_ipsecsha (accessed on 31 July 2020).
- Frazer, L.C.; Scurlock, A.M.; Zurenski, M.A.; Riley, M.M.; Mintus, M.; Pociask, D.A.; Sullivan, J.E.; Andrews, C.W.; Darville, T. IL-23 Induces IL-22 and IL-17 Production in Response to Chlamydia Muridarum Genital Tract Infection, but the Absence of These Cytokines Does Not Influence Disease Pathogenesis. Am. J. Reprod. Immunol. 2013, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, J.; Haga, S.; Hashimoto, K.; Okubo, T.; Ozawa, T.; Ozaki, M.; Yamaguchi, H. Activation of Caspase-3 during Chlamydia Trachomatis-Induced Apoptosis at a Late Stage. Can. J. Microbiol. 2019, 65, 135–143. [Google Scholar] [CrossRef] [PubMed]
- She, Q.-B.; Ma, W.-Y.; Zhong, S.; Dong, Z. Activation of JNK1, RSK2, and MSK1 Is Involved in Serine 112 Phosphorylation of Bad by Ultraviolet B Radiation. J. Biol. Chem. 2002, 277, 24039–24048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, S.J.; Stuart, K.; Gilley, R.; Sale, M.J. Control of Cell Death and Mitochondrial Fission by ERK1/2 MAP Kinase Signalling. FEBS J. 2017, 284, 4177–4195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortesoja, M.; Trofin, R.E.; Hanski, L. A Platform for Studying the Transfer of Chlamydia Pneumoniae Infection between Respiratory Epithelium and Phagocytes. J. Microbiol. Methods 2020, 171, 105857. [Google Scholar] [CrossRef]
- Lara, R.; Seckl, M.J.; Pardo, O.E. The P90 RSK Family Members: Common Functions and Isoform Specificity. Cancer Res. 2013, 73, 5301–5308. [Google Scholar] [CrossRef] [Green Version]
- Mullin, S.; Smith, L.; Lee, K.; D’Souza, G.; Woodgate, P.; Elflein, J.; Hällqvist, J.; Toffoli, M.; Streeter, A.; Hosking, J.; et al. Ambroxol for the Treatment of Patients With Parkinson Disease With and Without Glucocerebrosidase Gene Mutations: A Nonrandomized, Noncontrolled Trial. JAMA Neurol. 2020, 77, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Olaleye, O.A.; Kaur, M.; Onyenaka, C.C. Ambroxol Hydrochloride Inhibits the Interaction between Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein’s Receptor Binding Domain and Recombinant Human ACE2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Alkotaji, M. Azithromycin and Ambroxol as Potential Pharmacotherapy for SARS-CoV-2. Int. J. Antimicrob. Agents 2020, 56, 106192. [Google Scholar] [CrossRef]
- Bradfute, S.B.; Ye, C.; Clarke, E.C.; Kumar, S.; Timmins, G.S.; Deretic, V. Ambroxol and Ciprofloxacin Show Activity Against SARS-CoV2 in Vero E6 Cells at Clinically-Relevant Concentrations. bioRxiv 2020. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kókai, D.; Paróczai, D.; Virok, D.P.; Endrész, V.; Gáspár, R.; Csont, T.; Bozó, R.; Burián, K. Ambroxol Treatment Suppresses the Proliferation of Chlamydia pneumoniae in Murine Lungs. Microorganisms 2021, 9, 880. https://doi.org/10.3390/microorganisms9040880
Kókai D, Paróczai D, Virok DP, Endrész V, Gáspár R, Csont T, Bozó R, Burián K. Ambroxol Treatment Suppresses the Proliferation of Chlamydia pneumoniae in Murine Lungs. Microorganisms. 2021; 9(4):880. https://doi.org/10.3390/microorganisms9040880
Chicago/Turabian StyleKókai, Dávid, Dóra Paróczai, Dezső Peter Virok, Valéria Endrész, Renáta Gáspár, Tamás Csont, Renáta Bozó, and Katalin Burián. 2021. "Ambroxol Treatment Suppresses the Proliferation of Chlamydia pneumoniae in Murine Lungs" Microorganisms 9, no. 4: 880. https://doi.org/10.3390/microorganisms9040880
APA StyleKókai, D., Paróczai, D., Virok, D. P., Endrész, V., Gáspár, R., Csont, T., Bozó, R., & Burián, K. (2021). Ambroxol Treatment Suppresses the Proliferation of Chlamydia pneumoniae in Murine Lungs. Microorganisms, 9(4), 880. https://doi.org/10.3390/microorganisms9040880