
Honeybees Exposure to Natural Feed Additives: How Is the Gut Microbiota Affected?






Abstract
:1. Introduction
2. Materials and Methods
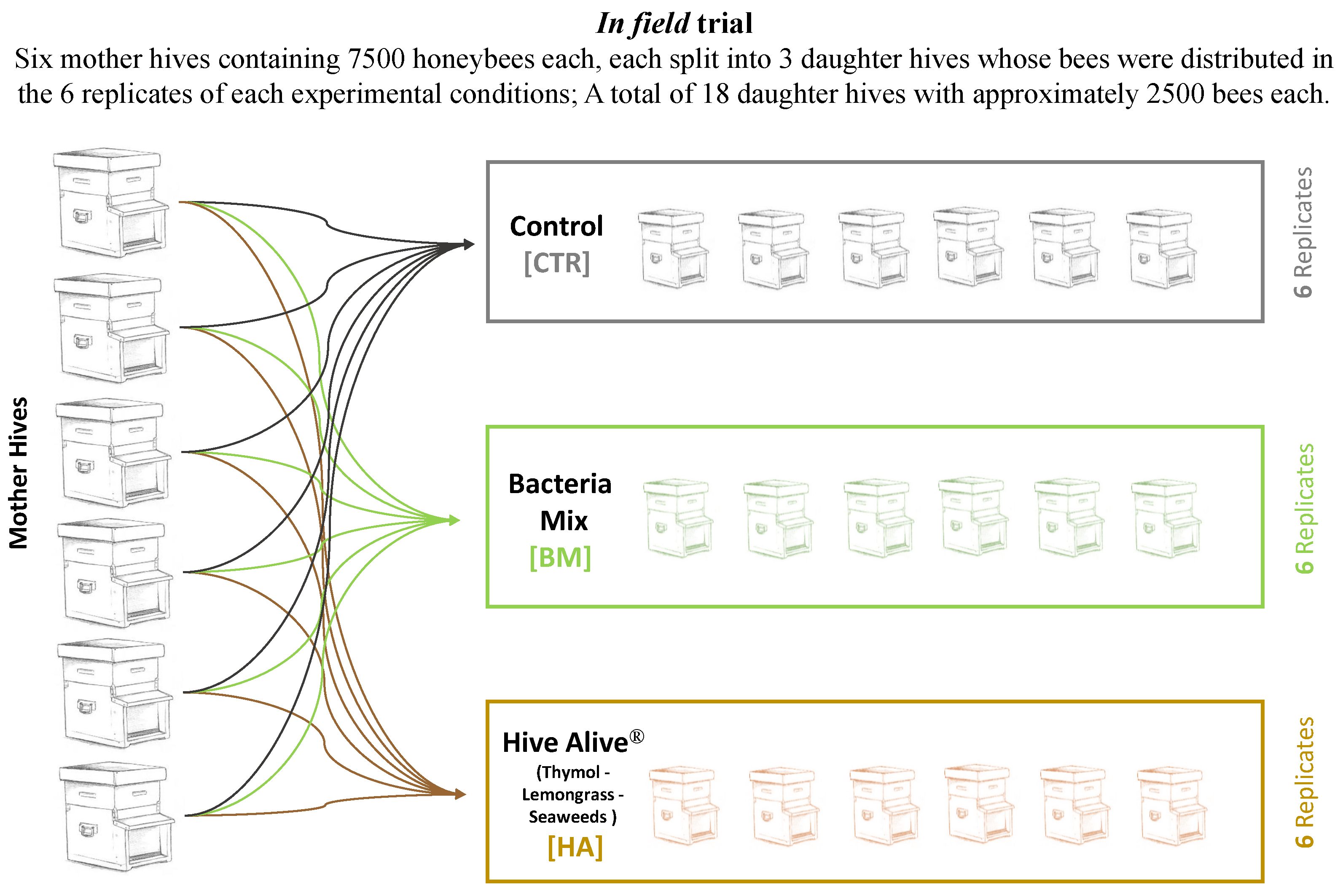
2.1. Experimental Design
2.2. Treatments and Samplings
2.3. DNA Extraction and NGS Sequencing
2.4. Quantification of Target Microbial Groups
2.5. Data Adjustments and Classification of Microbial Genera
2.6. Statistical Analysis
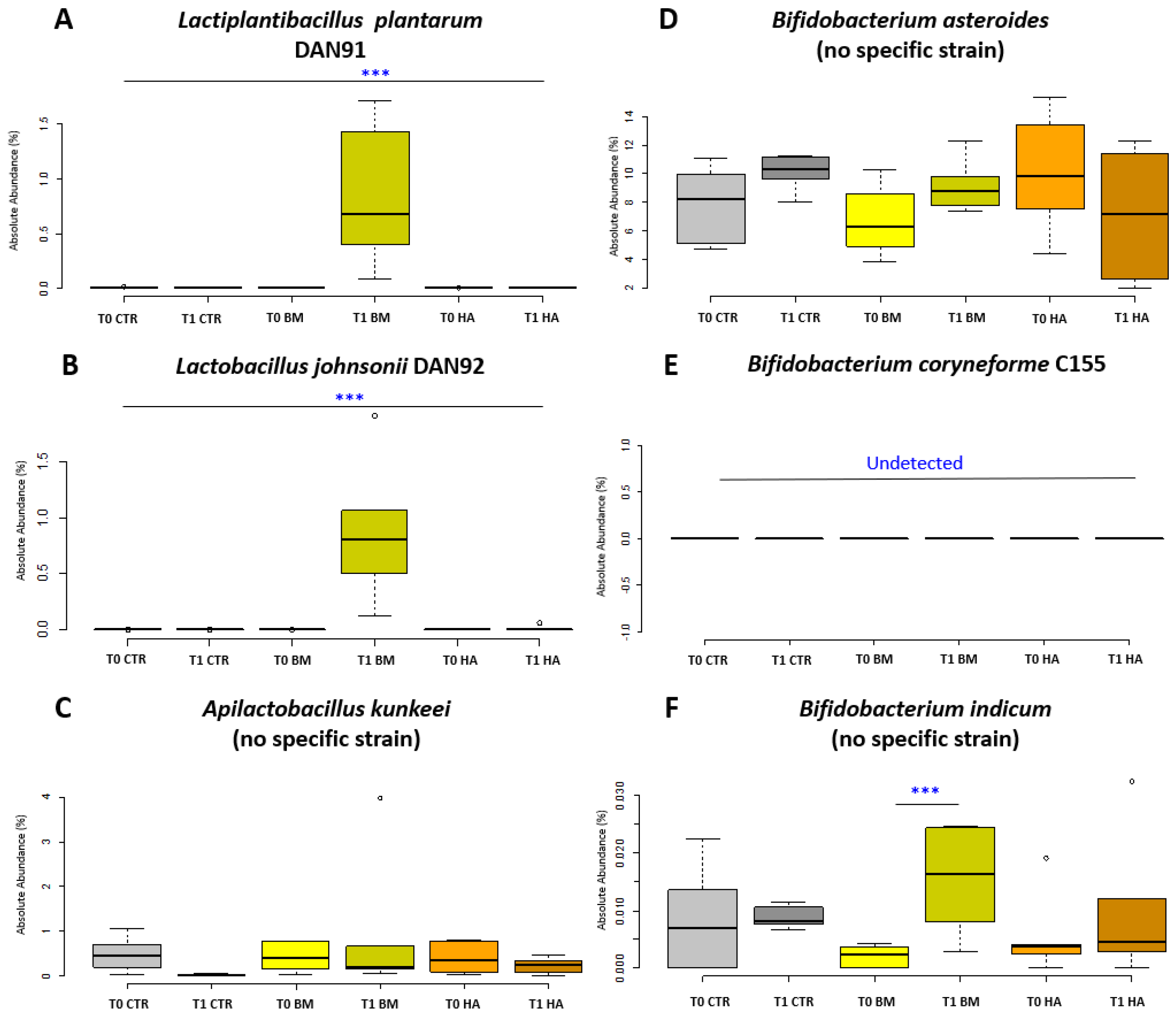
3. Results
3.1. General Observations on the Colony’S Status Pre and Post Treatment
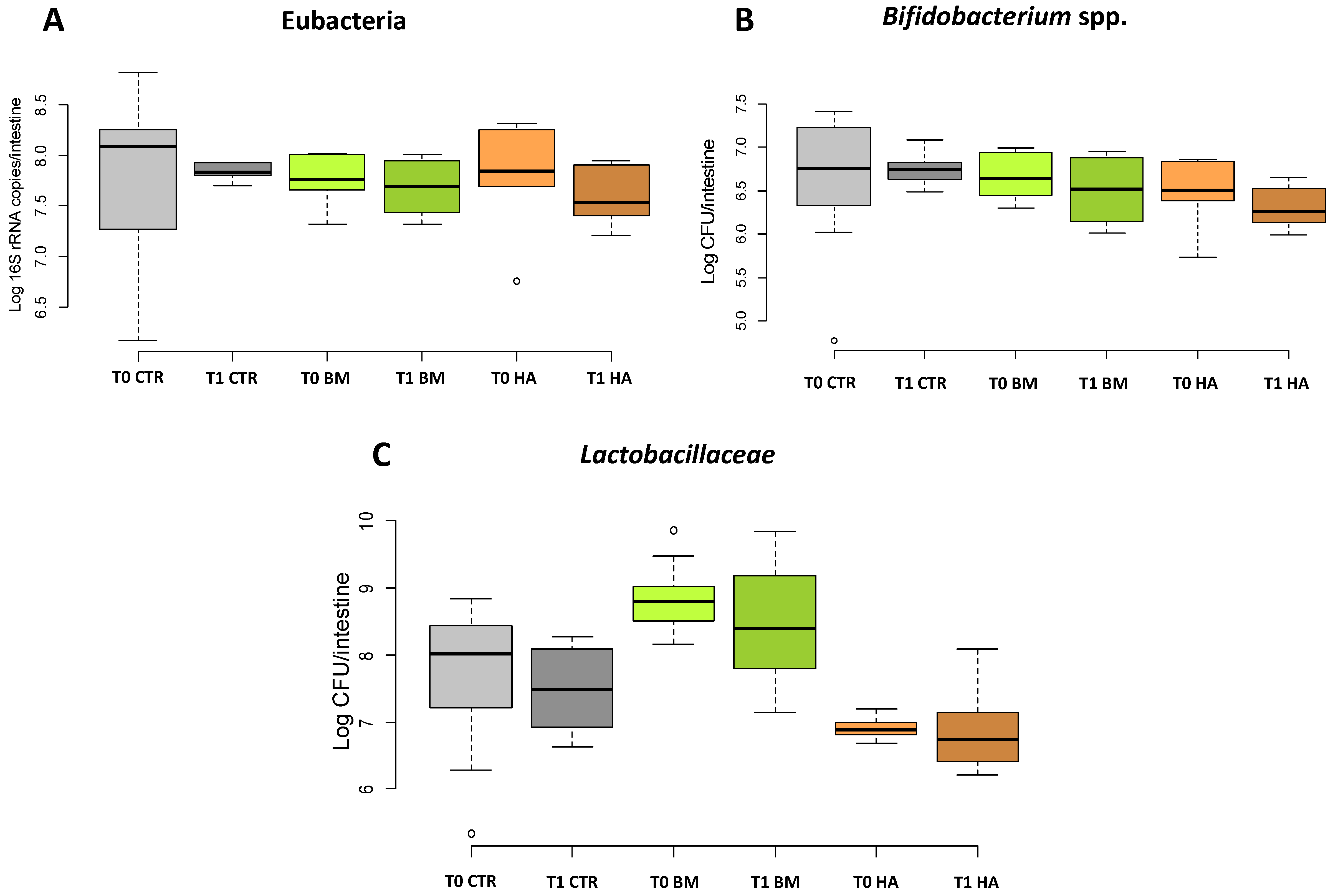
3.2. qPCR Quantification of Total Bacteria, Bifidobacterium and Lactobacillaceae in the Bee Gut
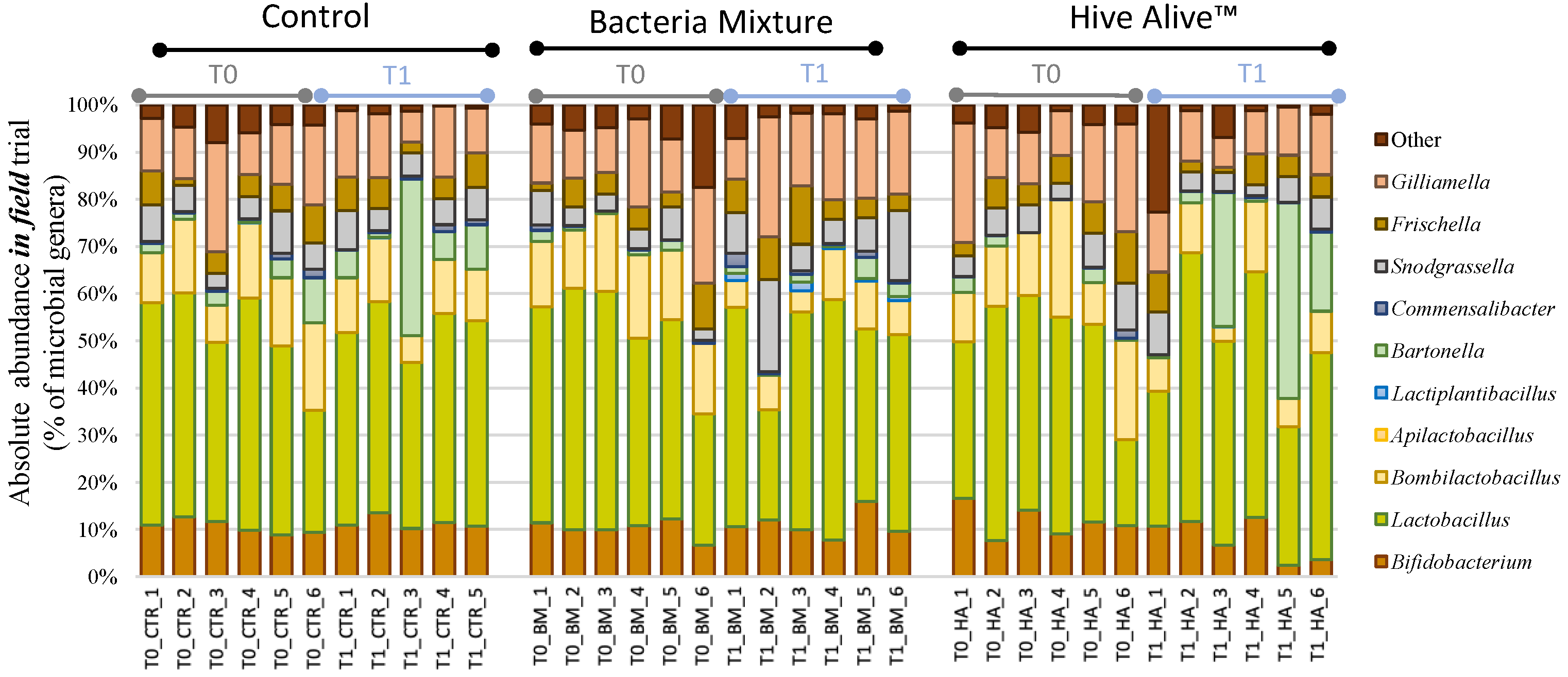
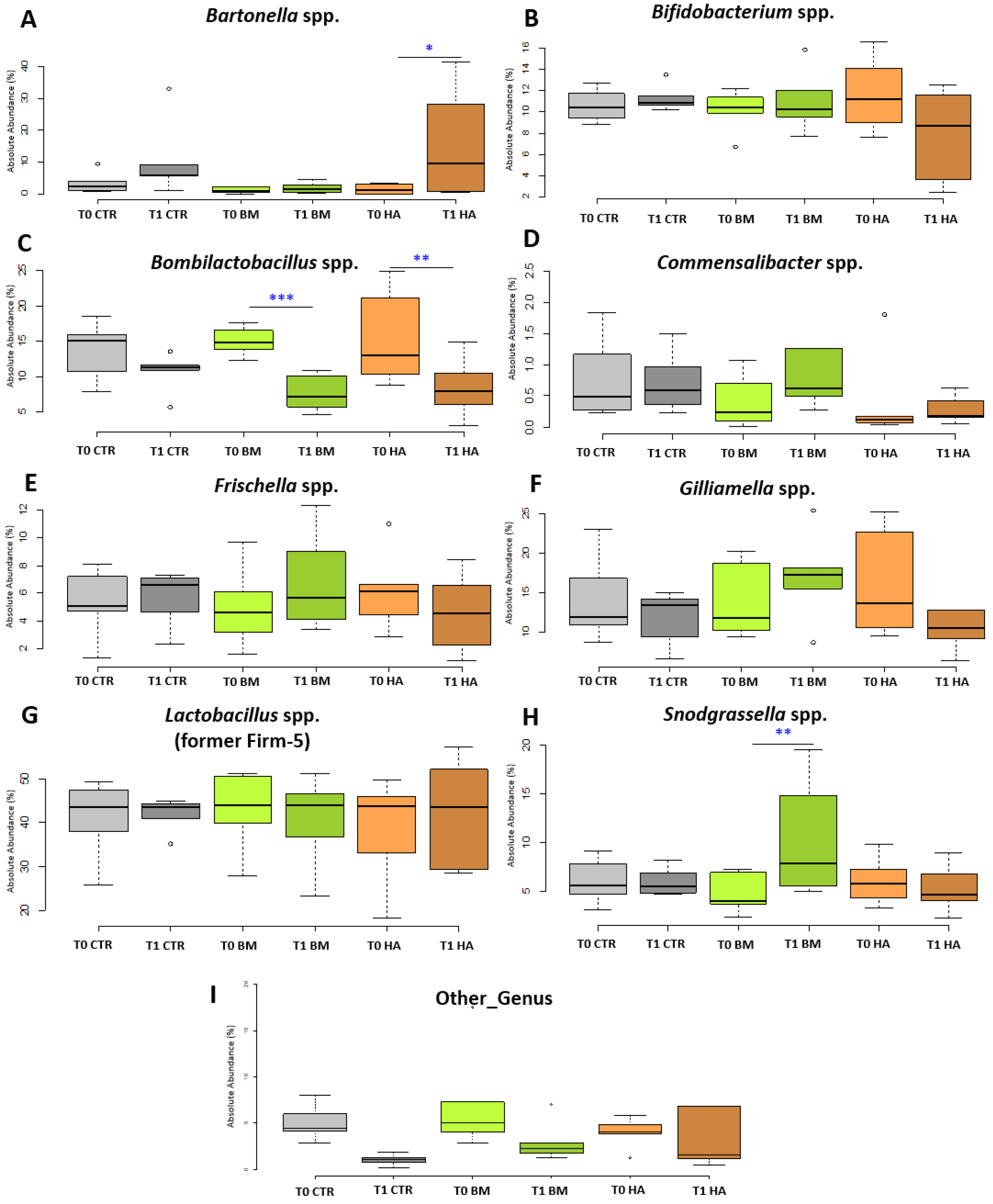
3.3. Bee Gut Microbiota Analysis via NGS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BM | Bacterial mixture |
| CFU | Colony forming unit |
| CTR | Control |
| HA | HiveAliveTM |
| NGS | Next Generation Sequencing |
| OTU | Open taxonomic unit |
| PCA | Principal component analysis |
| PD | Phylogenetic diversity |
| qPCR | Quantitative Polymerise chain reaction |
| T0 | Experiment begins |
| T1 | Experiment end |
References
- Tepedino, V.J. The importance of bees and other insect pollinators in maintaining floral species composition. Great Basin Nat. Mem. 1979, 3, 139–150. [Google Scholar]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Kellar, B.M. Honey Bees and Apple Trees: Hood River, Oregon as a Case Study for the Creation of the Honey Bee Pollination Industry; Oregon State University Archive: Hood River County, OR, USA, 2018. [Google Scholar]
- Sáez, A.; Aizen, M.A.; Medici, S.; Viel, M.; Villalobos, E.; Negri, P. Bees increase crop yield in an alleged pollinator-independent almond variety. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Fontana, P.; Costa, C.; Di Prisco, G.; Ruzzier, E.; Annoscia, D.; Battisti, A.; Caoduro, G.; Carpana, C.; Contessi, A.; Dal Lago, A.; et al. Appeal for biodiversity protection of native honey bee subspecies of Apis mellifera in Italy (San Michele all’Adige declaration). Sci. Bull. Insectol. 2018, 71, 257–271. [Google Scholar]
- Requier, F.; Garnery, L.; Kohl, P.L.; Njovu, H.K.; Pirk, C.W.; Crewe, R.M.; Steffan-Dewenter, I. The conservation of native honey bees is crucial. Sci. Trends Ecol. Evol. 2019, 34, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, J.; Knispel, H.; Hertlein, G.; Fünfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef]
- Forsgren, E.; Locke, B.; Sircoulomb, F.; Schäfer, M.O. Bacterial diseases in honeybees. Curr. Clin. Microbiol. Rep. 2018, 5, 18–25. [Google Scholar] [CrossRef]
- Charbonneau, L.R.; Hillier, N.K.; Rogers, R.E.; Williams, G.R.; Shutler, D. Effects of Nosema apis, N. ceranae, and coinfections on honey bee (Apis mellifera) learning and memory. Sci. Rep. 2016, 6, 22626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinpoo, C.; Paxton, R.J.; Disayathanoowat, T.; Krongdang, S.; Chantawannakul, P. Impact of Nosema ceranae and Nosema apis on individual worker bees of the two host species (Apis cerana and Apis mellifera) and regulation of host immune response. J. Insect. Physiol. 2018, 105, 1–8. [Google Scholar] [CrossRef]
- Moritz, R.F.A.; Erler, S. Lost colonies found in a data mine: Global honey trade but not pests or pesticides as a major cause of regional honeybee colony declines. Agric. Ecosyst. Environ. 2016, 216, 44–50. [Google Scholar] [CrossRef]
- Maini, S.; Medrzycki, P.; Porrini, C. The puzzle of honey bee losses: A brief review. Bull. Insect. 2010, 63, 153–160. [Google Scholar]
- Commission Regulation (EU) No. 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regarding maximum residue limits in foodstuffs of animal origin with honey bee (Apis mellifera) foragers. Off. J. Eur. Union. 2010, 15, 1–72.
- Sneeringer, S.; Bowman, M.; Clancy, M. The US and EU Animal Pharmaceutical Industries in the Age of Antibiotic Resistance. Sci. Trends Ecol. Evolut. 2019, 34, 789–798. [Google Scholar] [CrossRef]
- Underwood, R.M.; Traver, B.E.; López-Uribe, M.M. Beekeeping management practices are associated with operation size and beekeepers’ philosophy towards in-hive chemicals. Insects 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Cilia, L. The plight of the honeybee: A socioecological analysis of large-scale beekeeping in the United States. Sociol. Ruralis. 2019, 59, 831–849. [Google Scholar] [CrossRef]
- Rice, R.N. Nosema Disease in Honeybees: Genetic Variation and Control: A Report for the Rural Industries Research and Development Corporation; Rural Industries Research and Development Corporation: Kingston, Astralia, 2001. [Google Scholar]
- Costa, C.; Lodesani, M.; Maistrello, L. Effect of thymol and resveratrol administered with candy or syrup on the development of Nosema ceranae and on the longevity of honeybees (Apis mellifera L.) in laboratory conditions. Apidologie 2010, 41, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Damiani, N.; Fernández, N.J.; Porrini, M.P.; Gende, L.B.; Álvarez, E.; Buffa, F.; Brasesco, C.; Maggi, M.D.; Marcangeli, J.A.; Eguaras, M.J. Laurel leaf extracts for honeybee pest and disease management: Antimicrobial, microsporicidal, and acaricidal activity. Parasitol. Res. 2014, 113, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Porrini, M.P.; Garrido, P.M.; Gende, L.B.; Rossini, C.; Hermida, L.; Marcángeli, J.A.; Eguaras, M.J. Oral administration of essential oils and main components: Study on honey bee survival and Nosema ceranae development. J. Apic. Res. 2017, 56, 616–624. [Google Scholar] [CrossRef]
- Audisio, M.C.; Sabate, D.C.; Benítez-Ahrendts, M.R. Effect of Lactobacillus johnsonii CRL1647 on different parameters of honeybee colonies and bacterial populations of the bee gut. Benef. Microbes 2015, 6, 687–695. [Google Scholar] [CrossRef]
- Alberoni, D.; Baffoni, L.; Gaggìa, F.; Ryan, P.M.; Murphy, K.; Ross, P.R.; Stanton, C.; Di Gioia, D. Impact of beneficial bacteria supplementation on the gut microbiota, colony development and productivity of Apis mellifera L. Benef. Microbes. 2018, 9, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Rosso, G.; Engel, P. Functional roles and metabolic niches in the honey bee gut microbiota. Curr. Opin. Microbiol. 2018, 43, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A. Genomics of the honey bee microbiome. Curr. Opin. Insect. Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.; Ratnayeke, N.; Moran, N.A. Strain diversity and host specificity in a specialized gut symbiont of honeybees and bumblebees. Mol. Ecol. 2016, 25, 4461–4471. [Google Scholar] [CrossRef] [Green Version]
- Raymann, K.; Shaffer, Z.; Moran, N.A. Antibiotic exposure perturbs the gut microbiota and elevates mortality in honeybees. PLoS Biol. 2017, 15, e2001861. [Google Scholar] [CrossRef]
- Motta, E.V.; Raymann, K.; Moran, N.A. Glyphosate perturbs the gut microbiota of honey bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [Green Version]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut microbiota structure differs between honeybees in winter and summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Zappi-Recordati, A. Facts about beekeeping in Italy. Bee World 1956, 37, 229–237. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggìa, F.; Alberoni, D.; Cabbri, R.; Nanetti, A.; Biavati, B.; Di Gioia, D. Effect of dietary supplementation of Bifidobacterium and Lactobacillus strains in Apis mellifera L. against Nosema ceranae. Benef. Microbes 2016, 7, 45–51. [Google Scholar] [CrossRef] [PubMed]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, G.; de Graaf, D.C. Stress indicator gene expression profiles, colony dynamics and tissue development of honey bees exposed to sub-lethal doses of imidacloprid in laboratory and field experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinson, V.G.; Moy, J.; Moran, N.A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberoni, D.; Favaro, R.; Baffoni, L.; Angeli, S.; Di Gioia, D. Neonicotinoids in the agroecosystem: In-field long-term assessment on honeybee colony strength and microbiome. Sci. Total. Environ. 2021, 144116. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggìa, F.; Di Gioia, D.; Santini, C.; Mogna, L.; Biavati, B. A Bifidobacterium-based synbiotic product to reduce the transmission of C. jejuni along the poultry food chain. Int. J. Food. Microbiol. 2012, 157, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.M.; Bussema, C., III; Schmidt, T.M. rrnDB: Documenting the number of rRNA and tRNA genes in bacteria and archaea. Nucleic. Acids. Res. 2009, 37, D489–D493. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, S.F.; Smith, B.J.; Hein, R.; Roller, B.R.; Schmidt, T.M. rrnDB: Improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic. Acids. Res. 2015, 43, D593–D598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiol. 2002, 148, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 23 March 2021).
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Laurence Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 1.0.7. 2020. Available online: https://rpkgs.datanovia.com/factoextra/ (accessed on 23 March 2021).
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Probert, H.M.; Smejkal, C.W.; Gibson, G.R. Using probiotics and prebiotics to improve gut health. Drug Discov. Today 2003, 8, 692–700. [Google Scholar] [CrossRef]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food. Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef]
- Audisio, M.; Benítez-Ahrendts, M. Lactobacillus johnsonii CRL1647, isolated from Apis mellifera L. bee-gut, exhibited a beneficial effect on honeybee colonies. Benef. Microbes 2011, 2, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.S.; Moran, N.A.; Evans, J.D. Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers. Proc. Natl. Acad. Sci. USA 2016, 113, 9345–9350. [Google Scholar] [CrossRef] [Green Version]
- Kwong, W.K.; Engel, P.; Koch, H.; Moran, N.A. Genomics and host specialization of honey bee and bumble bee gut symbionts. Proc. Natl. Acad. Sci. USA 2014, 111, 11509–11514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The bacterial communities associated with honey bee (Apis mellifera) foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef] [Green Version]
- Tamarit, D.; Ellegaard, K.M.; Wikander, J.; Olofsson, T.; Vasquez, A.; Andersson, S.G. Functionally structured genomes in Lactobacillus kunkeei colonizing the honey crop and food products of honeybees and stingless bees. Genome Boil. Evol. 2015, 7, 1455–1473. [Google Scholar] [CrossRef]
- Raymann, K.; Coon, K.L.; Shaffer, Z.; Salisbury, S.; Moran, N.A. Pathogenicity of Serratia marcescens strains in honey bees. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Modesto, M.M.; Biavati, B.; Di Gioia, D. Bifidobacterium xylocopae sp. nov. and Bifidobacterium aemilianum sp. nov., from the carpenter bee (Xylocopa violacea) digestive tract. Syst. App. Microbiol. 2019, 42, 205–216. [Google Scholar] [CrossRef]
- Lockyer, S.; Aguirre, M.; Durrant, L.; Pot, B.; Suzuki, K. The role of probiotics on the roadmap to a healthy microbiota: A symposium report. Gut Microbiome 2020, 1. [Google Scholar] [CrossRef]
- Flint, J.F.; Garner, M.R. Feeding beneficial bacteria: A natural solution for increasing efficiency and decreasing pathogens in animal agriculture. J. Appl. Poult. Res. 2009, 18, 367–378. [Google Scholar] [CrossRef]
- Schmidt, K.; Engel, P. Probiotic Treatment with a Gut Symbiont Leads to Parasite Susceptibility in Honey Bees. Trends Parasitol. 2016, 32, 914–916. [Google Scholar] [CrossRef]
- El Khoury, S.; Rousseau, A.; Lecoeur, A.; Cheaib, B.; Bouslama, S.; Mercier, P.L.; Demey, V.; Castex, M.; Giovenazzo, P.; Derome, N. Deleterious interaction between honeybees (Apis mellifera) and its microsporidian intracellular parasite N. ceranae was mitigated by administrating either endogenous or allochthonous gut microbiota strains. Front. Ecol. Evol. 2018, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Arredondo, D.; Castelli, L.; Porrini, M.P.; Garrido, P.M.; Eguaras, M.J.; Zunino, P.; Antúnez, K. Lactobacillus kunkeei Strains Decreased the Infection by Honey Bee Pathogens Paenibacillus larvae and N. ceranae. Benef. Microbes 2018, 9, 279–290. [Google Scholar] [CrossRef]
- Shumkova, R.; Balkanska, R.; Hristov, P. The Herbal Supplements NOZEMAT HERB® and NOZEMAT HERB PLUS®: An Alternative Therapy for N. ceranae Infection and Its Effects on Honey Bee Strength and Production Traits. Pathogens 2021, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Cilia, G.; Garrido, C.; Bonetto, M.; Tesoriero, D.; Nanetti, A. Effect of Api-Bioxal® and ApiHerb® Treatments against N. ceranae Infection in Apis mellifera Investigated by Two QPCR Methods. Vet. Sci. 2020, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Charistos, L.; Parashos, N.; Hatjina, F. Long Term Effects of a Food Supplement HiveAliveTM on Honey Bee Colony Strength and N. ceranae Spore Counts. J. Apic. Res. 2015, 54, 420–426. [Google Scholar] [CrossRef]
- Diaz, T.; del-Val, E.; Ayala, R.; Larsen, J. Alterations in honey bee gut microorganisms caused by Nosema spp. and pest control methods. Pest. Manag. Sci. 2019, 75, 835–843. [Google Scholar] [CrossRef]
- Segers, F.H.I.D.; Kesnerova, L.; Kosoy, M.; Engel, P. Genomic changes associated with the evolutionary transition of an insect gut symbiont into a blood-borne pathogen. ISME J. 2017, 11, 1232–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, L.; Branchiccela, B.; Garrido, M.; Invernizzi, C.; Porrini, M.; Romero, H.; Santos, E.; Zunino, P.; Antúnez, K. Impact of Nutritional Stress on Honeybee Gut Microbiota, Immunity, and Nosema ceranae Infection. Microb. Ecol. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lucia, A.; Audino, P.G.; Seccacini, E.; Licastro, S.; Zerba, E.; Masuh, H. Larvicidal effect of Eucalyptus grandis essential oil and turpentine and their major components on Aedes aegypti larvae. J. Am. Mosq. Control. Assoc. 2007, 23, 299–303. [Google Scholar] [CrossRef]
- Motta, E.V.; Moran, N.A. Impact of glyphosate on the honey bee gut microbiota: Effects of intensity, duration, and timing of exposure. Msystems 2020, 5. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Theses | Dose per Treatment * | Reference Article | ||
|---|---|---|---|---|
| Thesis Acronym | Active Ingredient | Commercial Brand | ||
| HA | Thymol + Seaweeds mixture | HiveAliveTM Advance Science | 300 L (2.4 mg of thymol) | Manufacturer instructions |
| BM | Bacteria mixture ** | N.A. | 2.2 × 10 bacterial cells/mL *** | [23,32] |
| CTR | - | - | - | - |
| PHYLA | ||||||||
|---|---|---|---|---|---|---|---|---|
| CTR_T0 | CTR_T1 | BM_T0 | BM_T1 | HA_T0 | HA_T1 | |||
| Actinobacteria | 10.58 ± 1.49 | 11.37 ± 1.42 | 10.16 ± 2.01 | 10.96 ± 3.02 | 11.61 ± 3.23 | 7.92 ± 4.28 | ||
| Firmicutes | 55.12 ± 9.20 | 52.36 ± 6.71 | 57.90 ± 9.48 | 49.36 ± 10.85 | 54.28 ± 11.22 | 50.76 ± 14.62 | ||
| Proteobacteria | 29.30 ± 9.53 | 35.19 ± 7.89 | 24.95 ± 10.24 | 36.75 ± 9.74 | 30.11 ± 10.31 | 35.56 ± 17.26 | ||
| Other_phyla | 5.00 ± 0.74 | 1.08 ± 0.39 | 6.99 ± 0.51 | 2.93 ± 0.14 | 4.00 ± 0.22 | 5.76 ± 0.31 | ||
| FAMILY | ||||||||
| CTR_T0 | CTR_T1 | BM_T0 | BM_T1 | HA_T0 | HA_T1 | |||
| Acetobacteraceae | 0.75 ± 0.33 | 0.73 ± 0.04 | 0.40 ± 1.34 | 1.02 ± 0.63 | 0.39 ± 1.68 | 0.27 ± 9.46 | ||
| Bartonellaceae | 3.38 ± 3.29 | 11.08 ± 12.70 | 1.11 ± 0.93 | 1.86 ± 1.59 | 1.53 ± 1.51 | 15.08 ± 17.04 | ||
| Bifidobacteraceae | 10.58 ± 1.51 | 11.37 ± 1.39 | 10.16 ± 1.97 | 10.96 ± 2.88 | 11.61 ± 3.26 | 7.92 ± 4.27 | ||
| Lactobacillaceae | 55.12 ± 8.43 | 52.36 ± 6.83 | 57.90 ± 8.58 | 49.36 ± 10.99 | 54.28 ± 11.84 | 50.76 ± 14.76 | ||
| Neisseriaceae | 5.99 ± 2.09 | 6.06 ± 1.50 | 4.73 ± 1.98 | 10.13 ± 5.77 | 6.08 ± 2.36 | 5.29 ± 2.22 | ||
| Orbaceae | 19.18 ± 6.16 | 17.32 ± 4.82 | 18.71 ± 6.74 | 23.74 ± 6.54 | 22.12 ± 7.61 | 14.92 ± 4.33 | ||
| Other_families | 5.00 ± 1.55 | 1.08 ± 0.31 | 6.99 ± 5.77 | 2.93 ± 1.29 | 4.00 ± 1.45 | 5.76 ± 1.95 | ||
| GENUS | ||||||||
| CTR_T0 | CTR_T1 | BM_T0 | BM_T1 | HA_T0 | HA_T1 | |||
| Apilactobacillus | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.00 ± 0.00 | ||
| Bartonella | 3.38 ± 3.25 | 11.08 ± 12.70 | 1.11 ± 0.97 | 1.86 ± 1.61 | 1.53 ± 1.52 | 15.08 ± 17.04 | ||
| Bifidobacterium | 10.58 ± 1.48 | 11.37 ± 1.28 | 10.16 ± 1.91 | 10.96 ± 2.79 | 11.61 ± 3.31 | 7.92 ± 4.33 | ||
| Bombilactobacillus | 13.88 ± 3.87 | 10.64 ± 2.94 | 15.00 ± 1.89 | 7.61 ± 2.44 | 15.20 ± 6.36 | 8.39 ± 4.05 | ||
| Commensalibacter | 0.75 ± 0.64 | 0.73 ± 0.51 | 0.40 ± 0.41 | 1.02 ± 0.95 | 0.39 ± 0.70 | 0.27 ± 0.21 | ||
| Frischella | 5.27 ± 2.38 | 5.58 ± 2.11 | 4.97 ± 2.79 | 6.69 ± 3.49 | 6.20 ± 2.76 | 4.59 ± 2.70 | ||
| Gilliamella | 13.91 ± 5.25 | 11.74 ± 3.57 | 13.74 ± 4.60 | 17.06 ± 5.37 | 15.92 ± 6.79 | 10.33 ± 2.41 | ||
| Lactobacillus | 41.24 ± 8.76 | 41.72 ± 3.98 | 42.91 ± 8.64 | 40.88 ± 9.91 | 39.07 ± 11.67 | 42.37 ± 11.59 | ||
| Plantilactobacillus | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.87 ± 0.66 | 0.00 ± 0.00 | 0.01 ± 0.01 | ||
| Snodgrassella | 5.99 ± 2.16 | 6.06 ± 1.50 | 4.73 ± 1.97 | 10.13 ± 5.80 | 6.08 ± 2.28 | 5.29 ± 2.39 | ||
| Other_genus | 5.00 ± 1.79 | 1.08 ± 0.62 | 6.99 ± 5.35 | 2.93 ± 2.10 | 4.00 ± 1.52 | 5.76 ± 8.64 | ||
| Group 1 | Group 2 | p | p |
|---|---|---|---|
| Weighted UniFrac | |||
| CTR_T0 vs. CTR_T0 | CTR_T1 vs. CTR_T1 | 0.085 | 0.935 |
| HA_T0 vs. HA_T0 | HA_T1 vs. HA_T1 | 0.012 | 0.132 |
| BM_T0 vs. BM_T0 | BM_T0 vs. BM_T1 | 0.936 | 1 |
| CTR_T0 vs. CTR_T1 | HA_T0 vs. HA_T1 | 0.002 | 0.022 ** |
| CTR_T0 vs. CTR_T1 | BM_T0 vs. BM_T1 | 0.014 | 0.154 |
| BM | HA | CTR | |
|---|---|---|---|
| Phyla | Other_phyla ↓ | ||
| Family | Neisseraceae ↑ Other_families ↓ | Other_families ↓ | |
| Genus | Snodgrassella↑ Bombilactobacillus ↓ | Bartonella↑ Bombilactobacillus ↓ | |
| Species | S. alvi↑ B. mellifer ↓ L. plantarum↑ B. indicum↑ L. jhonsoniiDan92↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberoni, D.; Baffoni, L.; Braglia, C.; Gaggìa, F.; Di Gioia, D. Honeybees Exposure to Natural Feed Additives: How Is the Gut Microbiota Affected? Microorganisms 2021, 9, 1009. https://doi.org/10.3390/microorganisms9051009
Alberoni D, Baffoni L, Braglia C, Gaggìa F, Di Gioia D. Honeybees Exposure to Natural Feed Additives: How Is the Gut Microbiota Affected? Microorganisms. 2021; 9(5):1009. https://doi.org/10.3390/microorganisms9051009
Chicago/Turabian StyleAlberoni, Daniele, Loredana Baffoni, Chiara Braglia, Francesca Gaggìa, and Diana Di Gioia. 2021. "Honeybees Exposure to Natural Feed Additives: How Is the Gut Microbiota Affected?" Microorganisms 9, no. 5: 1009. https://doi.org/10.3390/microorganisms9051009
APA StyleAlberoni, D., Baffoni, L., Braglia, C., Gaggìa, F., & Di Gioia, D. (2021). Honeybees Exposure to Natural Feed Additives: How Is the Gut Microbiota Affected? Microorganisms, 9(5), 1009. https://doi.org/10.3390/microorganisms9051009