
Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens


 , ,
, ,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Microbiological Media
2.2. Bacterial Strains
2.3. Determination the Minimum Inhibitory Concentration (MIC) of Xylitol
2.4. Evaluation of Biofilm Production
2.5. Biofilm Inhibition Assay
2.6. Prodigiosin Inhibition Assay
2.7. Protease Assay
2.8. Assay of Bacterial Motilities
2.9. Oxidative Stress Assay
2.10. Quantification of Virulence Genes’ Expressions
2.11. Mice Survival Test
2.12. In-Silico Analysis of Binding to SmaR Receptor
2.13. Statistical Analysis
3. Results
3.1. Determination of MIC
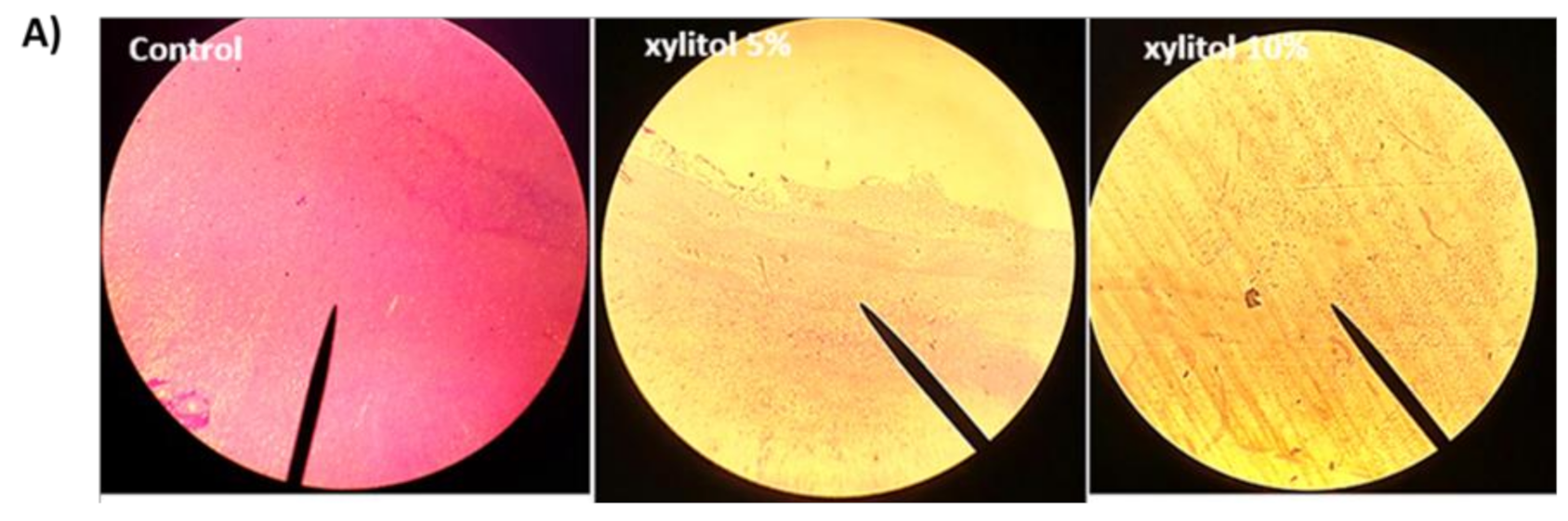
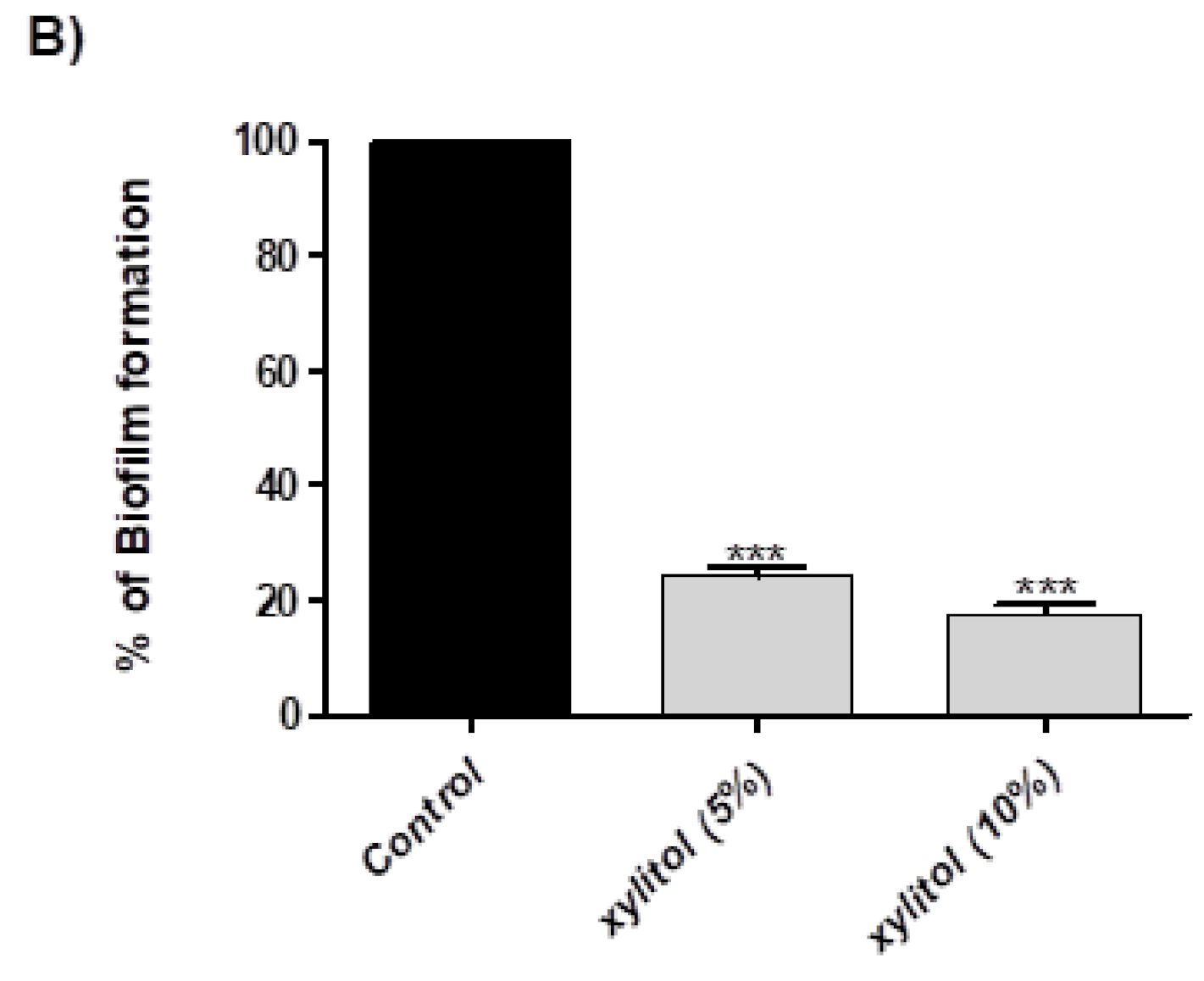
3.2. Xylitol Inhibits Biofilm Formation
3.3. Xylitol Decreases Prodigiosin Production
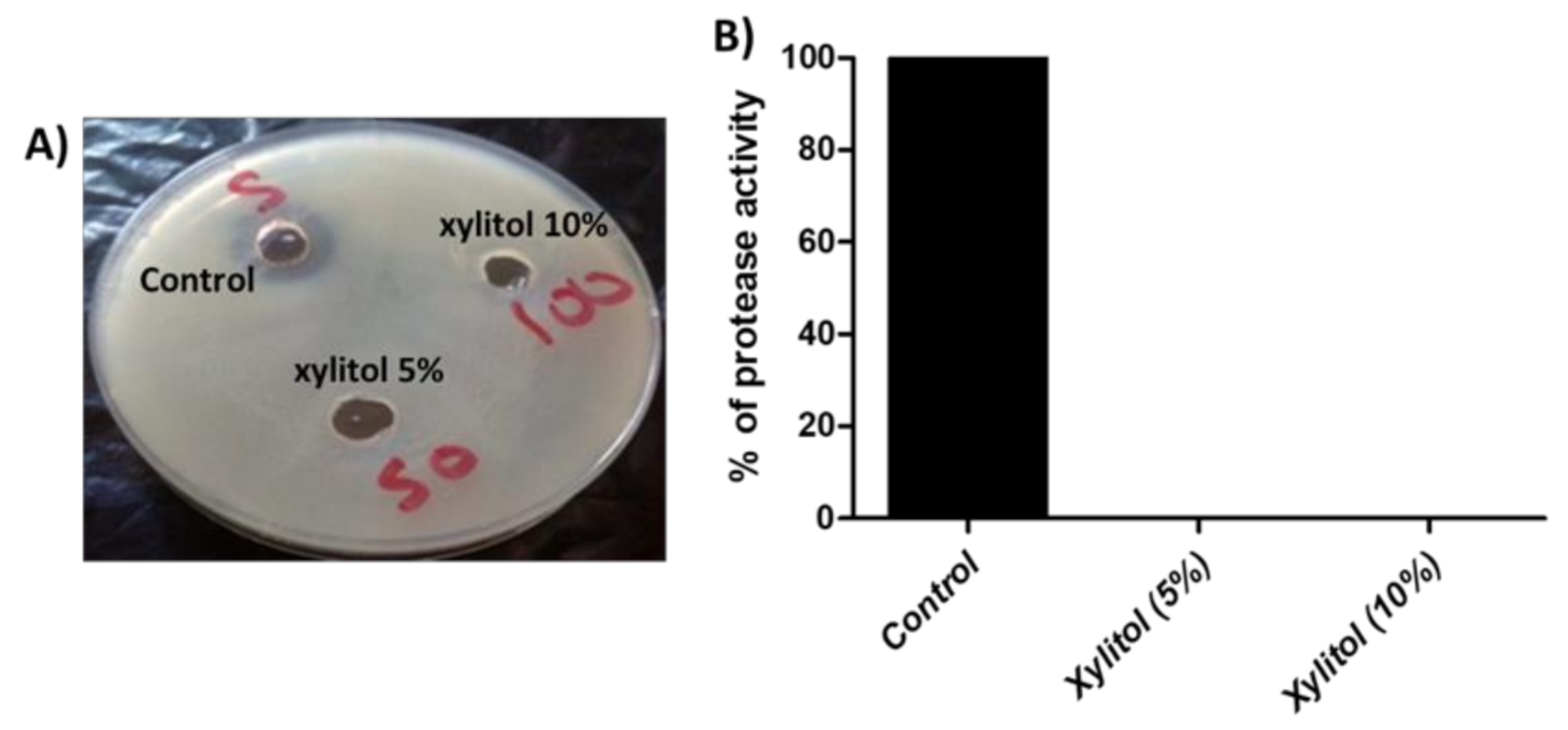
3.4. Xylitol Interferes with Protease Activity
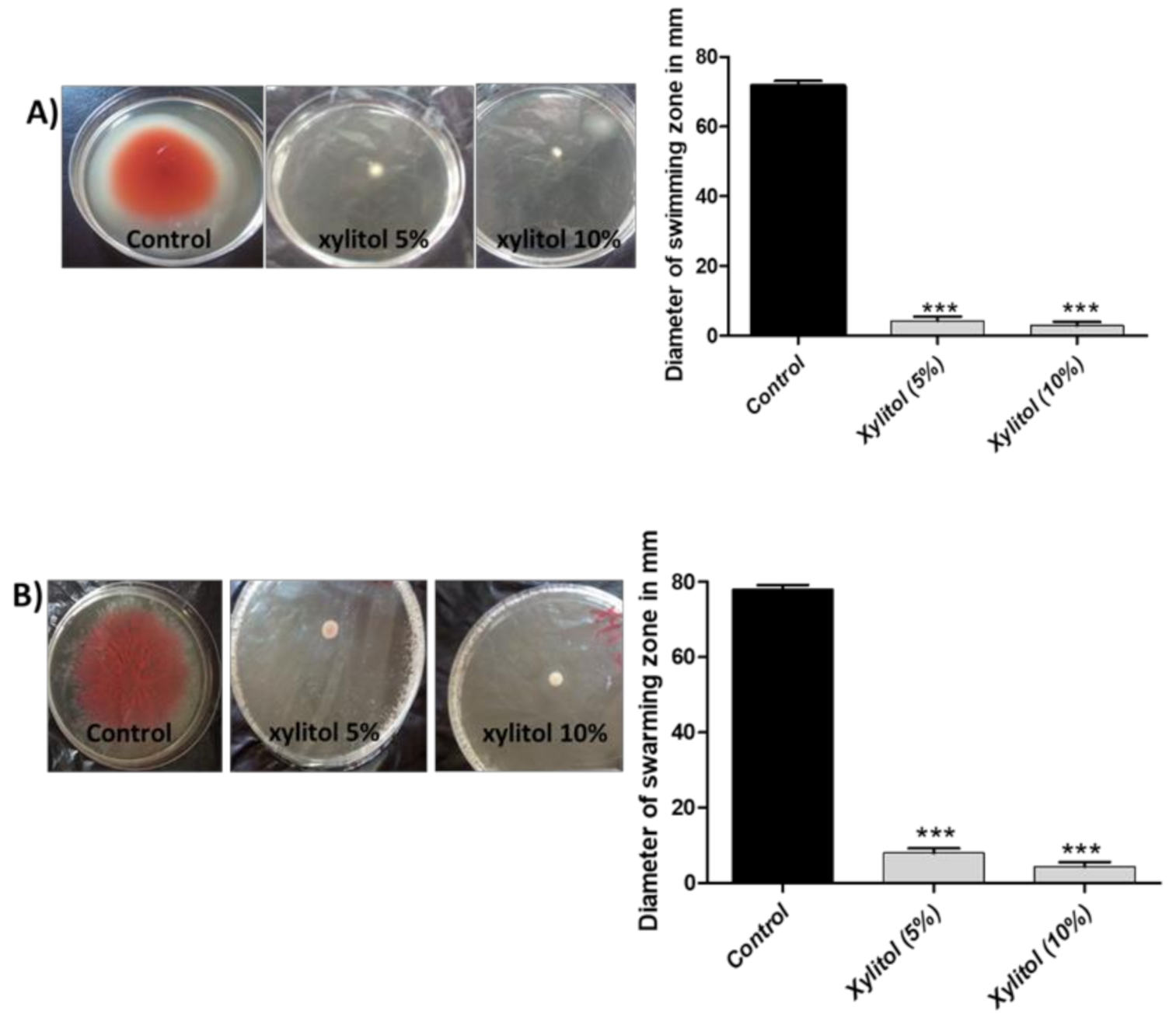
3.5. Xylitol Inhibits Swimming and Swarming Motilities
3.6. Xylitol Increases the Sensitivity to Oxidative Stress
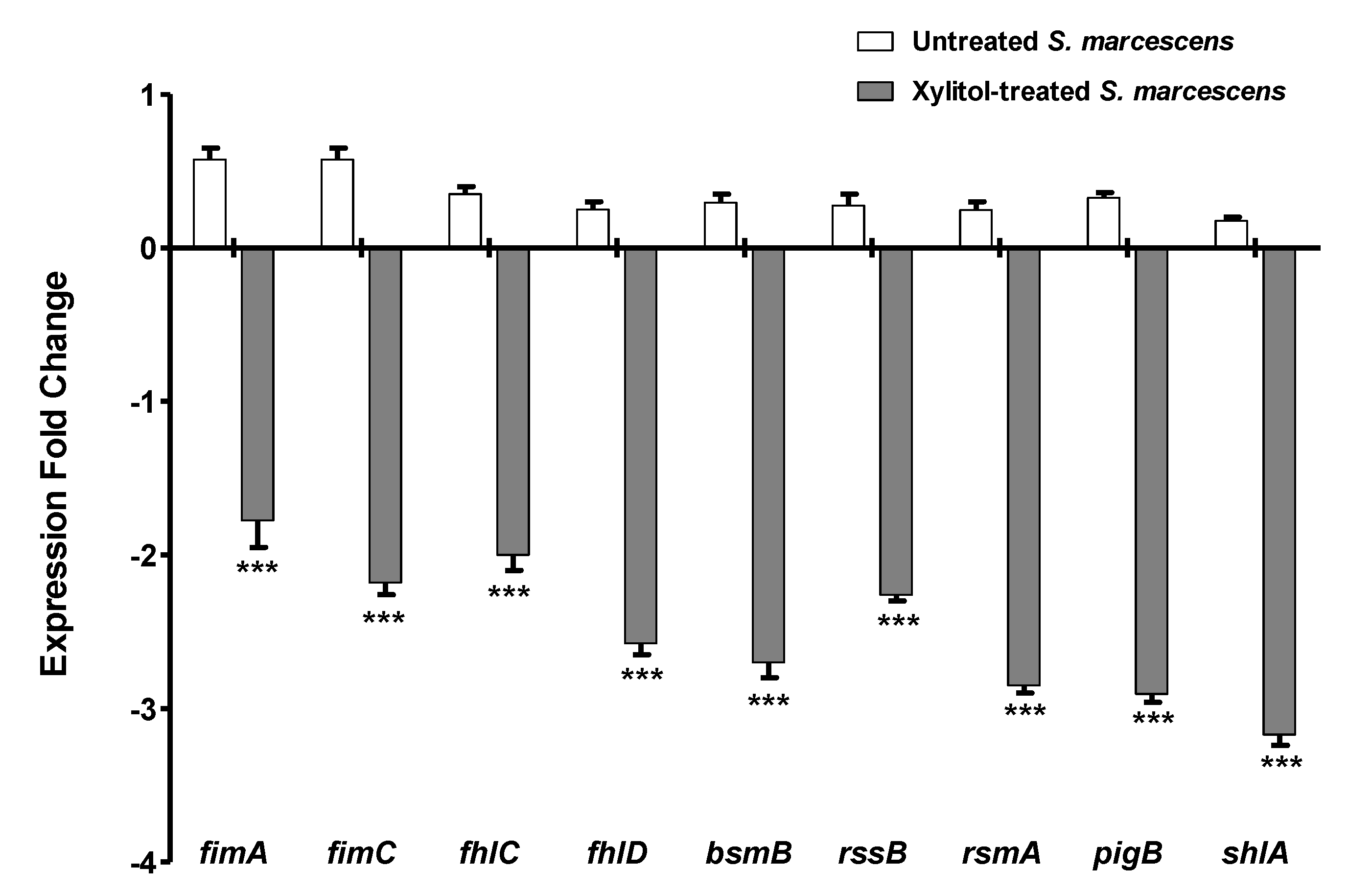
3.7. Xylitol Decreased the Expression of S. marcescens Virulence Genes
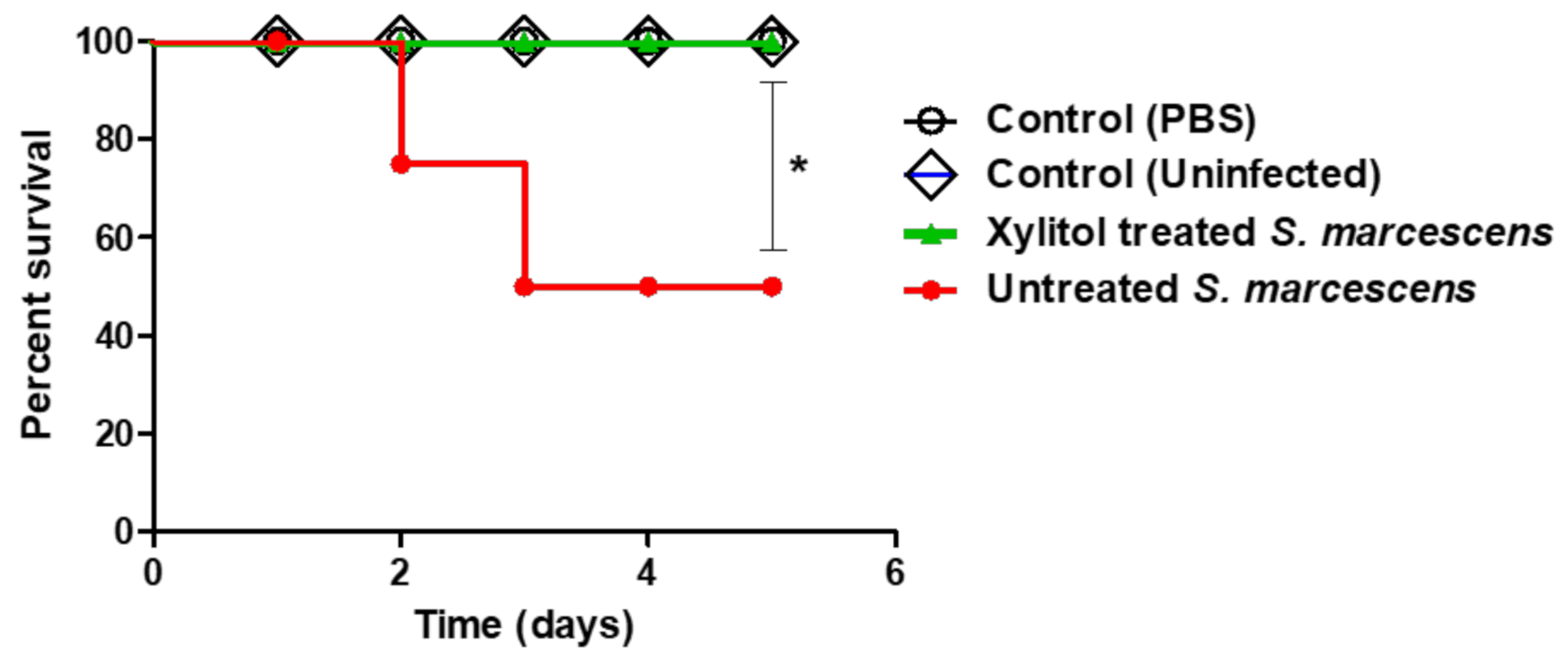
3.8. In-Vivo Protection Activity of Xylitol Against S. marcescens
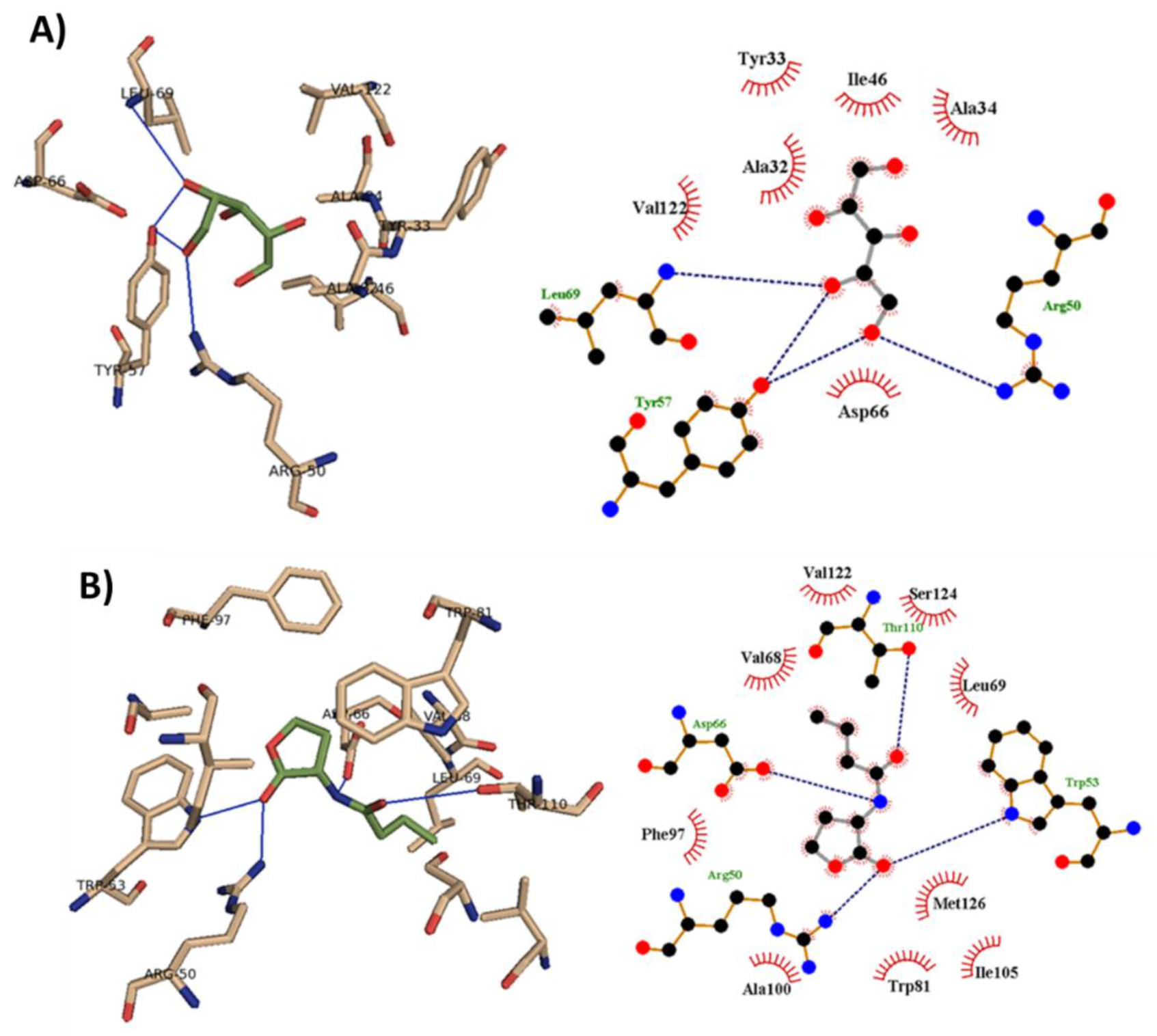
3.9. Xylitol Could Bind to Smar Receptor In-Silico
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ramanathan, S.; Ravindran, D.; Arunachalam, K.; Arumugam, V.R. Inhibition of quorum sensing-dependent biofilm and virulence genes expression in environmental pathogen Serratia marcescens by petroselinic acid. Antonie Van Leeuwenhoek 2018, 111, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Us, E.; Kutlu, H.H.; Tekeli, A.; Ocal, D.; Cirpan, S.; Memikoglu, K.O. Wound and soft tissue infections of Serratia marcescens in patients receiving wound care: A health care-associated outbreak. Am. J. Infect. Control. 2017, 45, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Posluszny, J.A., Jr.; Conrad, P.; Halerz, M.; Shankar, R.; Gamelli, R.L. Surgical burn wound infections and their clinical implications. J. Burn Care Res. 2011, 32, 324–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, D.; Elsayed, S.; Reid, O.; Winston, B.; Lindsay, R. Burn wound infections. Clin. Microbiol. Rev. 2006, 19, 403–434. [Google Scholar] [CrossRef] [Green Version]
- Fekrirad, Z.; Kashef, N.; Arefian, E. Photodynamic inactivation diminishes quorum sensing-mediated virulence factor production and biofilm formation of Serratia marcescens. World J. Microbiol. Biotechnol. 2019, 35, 191. [Google Scholar] [CrossRef]
- Van Houdt, R.; Givskov, M.; Michiels, C.W. Quorum sensing in Serratia. FEMS Microbiol. Rev. 2007, 31, 407–424. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.A.; Hegazy, W.A.H. Repurposing anti-diabetic drug “Sitagliptin” as a novel virulence attenuating agent in Serratia marcescens. PLoS ONE 2020, 15, e0231625. [Google Scholar] [CrossRef] [Green Version]
- Williamson, N.R.; Simonsen, H.T.; Ahmed, R.A.; Goldet, G.; Slater, H.; Woodley, L.; Leeper, F.J.; Salmond, G.P. Biosynthesis of the red antibiotic, prodigiosin, in Serratia: Identification of a novel 2-methyl-3-n-amyl-pyrrole (MAP) assembly pathway, definition of the terminal condensing enzyme, and implications for undecylprodigiosin biosynthesis in Streptomyces. Mol. Microbiol. 2005, 56, 971–989. [Google Scholar] [CrossRef]
- Stock, I.; Burak, S.; Sherwood, K.J.; Gruger, T.; Wiedemann, B. Natural antimicrobial susceptibilities of strains of ’unusual’ Serratia species: S. ficaria, S. fonticola, S. odorifera, S. plymuthica and S. rubidaea. J. Antimicrob. Chemother. 2003, 51, 865–885. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.A.; Hegazy, W.A.H. Targeting the virulence factors of Serratia marcescens by ambroxol. Roum. Arch. Microbiol. Immunol. 2017, 76, 27–32. [Google Scholar]
- Abbas, H.A.; Soliman, W.E.; Shaldam, M.A. Perturbation of Quorum Sensing in Pseudomonas aeruginosa by Febuxostat. Adv. Microbiol. 2018, 650–664. [Google Scholar] [CrossRef] [Green Version]
- Labbate, M.; Zhu, H.; Thung, L.; Bandara, R.; Larsen, M.R.; Willcox, M.D.; Givskov, M.; Rice, S.A.; Kjelleberg, S. Quorum-sensing regulation of adhesion in Serratia marcescens MG1 is surface dependent. J. Bacteriol. 2007, 189, 2702–2711. [Google Scholar] [CrossRef] [Green Version]
- Devi, K.R.; Srinivasan, S.; Ravi, A.V. Inhibition of quorum sensing-mediated virulence in Serratia marcescens by Bacillus subtilis R-18. Microb. Pathog. 2018, 120, 166–175. [Google Scholar] [CrossRef]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Hegazy, W.A.H.; Khayat, M.T.; Ibrahim, T.S.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Repurposing Anti-diabetic Drugs to Cripple Quorum Sensing in Pseudomonas aeruginosa. Microorganisms 2020, 8, 1285. [Google Scholar] [CrossRef]
- Chirife, J.; Scarmato, G.; Herszage, L. Scientific basis for use of granulated sugar in treatment of infected wounds. Lancet 1982, 1, 560–561. [Google Scholar] [CrossRef]
- Zhou, D.P.; Lu, L.Q.; Mao, X.L. Insulin and hyperosmotic glucose solution external used for treating pressure sore. Hunan Yi Ke Da Xue Xue Bao 2001, 26, 475–476. [Google Scholar]
- Rabago, D.; Zgierska, A.; Fortney, L.; Kijowski, R.; Mundt, M.; Ryan, M.; Grettie, J.; Patterson, J.J. Hypertonic dextrose injections (prolotherapy) for knee osteoarthritis: Results of a single-arm uncontrolled study with 1-year follow-up. J. Altern. Complement. Med. 2012, 18, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Tapiainen, T.; Kontiokari, T.; Sammalkivi, L.; Ikaheimo, I.; Koskela, M.; Uhari, M. Effect of xylitol on growth of Streptococcus pneumoniae in the presence of fructose and sorbitol. Antimicrob. Agents Chemother. 2001, 45, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Trahan, L.; Mouton, C. Selection for Streptococcus mutans with an altered xylitol transport capacity in chronic xylitol consumers. J. Dent. Res. 1987, 66, 982–988. [Google Scholar] [CrossRef]
- Dowd, S.E.; Sun, Y.; Smith, E.; Kennedy, J.P.; Jones, C.E.; Wolcott, R. Effects of biofilm treatments on the multi-species Lubbock chronic wound biofilm model. J. Wound Care 2009, 18, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Ammons, M.C.; Ward, L.S.; James, G.A. Anti-biofilm efficacy of a lactoferrin/xylitol wound hydrogel used in combination with silver wound dressings. Int. Wound J. 2011, 8, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, R.; Joshi-Navare, K.; Prabhune, A. Crystalline xylitol production by a novel yeast, Pichia caribbica (HQ222812), and its application for quorum sensing inhibition in gram-negative marker strain Chromobacterium violaceum CV026. Appl. Biochem. Biotechnol. 2013, 169, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.A.; Serry, F.M.; El-Masry, E.M. Combating Pseudomonas aeruginosa biofilms by potential biofilm inhibitors. Asian J. Res. Pharm. Sci. 2012, 2, 66–72. [Google Scholar]
- Hegazy, W.A.H.; Khayat, M.T.; Ibrahim, T.S.; Youns, M.; Mosbah, R.; Soliman, W.E. Repurposing of antidiabetics as Serratia marcescens virulence inhibitors. Braz J. Microbiol. 2021, 52, 627–638. [Google Scholar] [CrossRef]
- Hegazy, W.A.H. Diclofenac inhibits virulence of Proteus mirabilis isolated from diabetic foot ulcer. Afr. J. Microbiol. Res. 2016, 10, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Issac Abraham, S.V.; Palani, A.; Ramaswamy, B.R.; Shunmugiah, K.P.; Arumugam, V.R. Antiquorum sensing and antibiofilm potential of Capparis spinosa. Arch. Med. Res. 2011, 42, 658–668. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; El-Naenaeey, E.-s.Y.; Kandeel, T.M.; Hegazy, W.A.H.; Mosbah, R.A.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 667. [Google Scholar] [CrossRef]
- Sarkar, R.; Chaudhary, S.K.; Sharma, A.; Yadav, K.K.; Nema, N.K.; Sekhoacha, M.; Karmakar, S.; Braga, F.C.; Matsabisa, M.G.; Mukherjee, P.K.; et al. Anti-biofilm activity of Marula—A study with the standardized bark extract. J. Ethnopharmacol. 2014, 154, 170–175. [Google Scholar] [CrossRef]
- Morohoshi, T.; Shiono, T.; Takidouchi, K.; Kato, M.; Kato, N.; Kato, J.; Ikeda, T. Inhibition of quorum sensing in Serratia marcescens AS-1 by synthetic analogs of N-acylhomoserine lactone. Appl. Environ. Microbiol. 2007, 73, 6339–6344. [Google Scholar] [CrossRef] [Green Version]
- Hassett, D.J.; Schweizer, H.P.; Ohman, D.E. Pseudomonas aeruginosa sodA and sodB mutants defective in manganese- and iron-cofactored superoxide dismutase activity demonstrate the importance of the iron-cofactored form in aerobic metabolism. J. Bacteriol. 1995, 177, 6330–6337. [Google Scholar] [CrossRef] [Green Version]
- Salini, R.; Pandian, S.K. Interference of quorum sensing in urinary pathogen Serratia marcescens by Anethum graveolens. Pathog. Dis. 2015, 73, ftv038. [Google Scholar] [CrossRef] [Green Version]
- Askoura, M.; Hegazy, W.A.H. Ciprofloxacin interferes with Salmonella Typhimurium intracellular survival and host virulence through repression of Salmonella pathogenicity island-2 (SPI-2) genes expression. Pathog. Dis. 2020, 78, ftaa011. [Google Scholar] [CrossRef]
- Bendary, M.M.; Ibrahim, D.; Mosbah, R.A.; Mosallam, F.; Hegazy, W.A.H.; Awad, N.F.S.; Alshareef, W.A.; Alomar, S.Y.; Zaitone, S.A.; Abd El-Hamid, M.I. Thymol Nanoemulsion: A New Therapeutic Option for Extensively Drug Resistant Foodborne Pathogens. Antibiotics 2020, 10, 25. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.H.; Byun, Y.; Park, H.D. 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci. Rep. 2015, 5, 8656. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.G.; Pappas, K.M.; Brace, J.L.; Miller, P.C.; Oulmassov, T.; Molyneaux, J.M.; Anderson, J.C.; Bashkin, J.K.; Winans, S.C.; Joachimiak, A. Structure of a bacterial quorum-sensing transcription factor complexed with pheromone and DNA. Nature 2002, 417, 971–974. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Lin, C.S.; Tsai, Y.H.; Chang, C.J.; Tseng, S.F.; Wu, T.R.; Lu, C.C.; Wu, T.S.; Lu, J.J.; Horng, J.T.; Martel, J.; et al. An iron detection system determines bacterial swarming initiation and biofilm formation. Sci. Rep. 2016, 6, 36747. [Google Scholar] [CrossRef]
- Rodrigues, A.P.; Holanda, A.R.; Lustosa, G.P.; Nobrega, S.M.; Santana, W.J.; Souza, L.B.; Coutinho, H.D. Virulence factors and resistance mechanisms of Serratia marcescens. A short review. Acta Microbiol. Immunol. Hung. 2006, 53, 89–93. [Google Scholar] [CrossRef]
- Jones, R.N. Microbial etiologies of hospital-acquired bacterial pneumonia and ventilator-associated bacterial pneumonia. Clin. Infect. Dis. 2010, 51, S81–S87. [Google Scholar] [CrossRef] [Green Version]
- Mangwani, N.; Kumari, S.; Das, S. Bacterial biofilms and quorum sensing: Fidelity in bioremediation technology. Biotechnol. Genet. Eng. Rev. 2016, 32, 43–73. [Google Scholar] [CrossRef]
- Zhou, J.; Lyu, Y.; Richlen, M.; Anderson, D.M.; Cai, Z. Quorum sensing is a language of chemical signals and plays an ecological role in algal-bacterial interactions. CRC Crit. Rev. Plant. Sci. 2016, 35, 81–105. [Google Scholar] [CrossRef]
- Srinivasan, R.; Mohankumar, R.; Kannappan, A.; Karthick Raja, V.; Archunan, G.; Karutha Pandian, S.; Ruckmani, K.; Veera Ravi, A. Exploring the Anti-quorum Sensing and Antibiofilm Efficacy of Phytol against Serratia marcescens Associated Acute Pyelonephritis Infection in Wistar Rats. Front. Cell Infect. Microbiol. 2017, 7, 498. [Google Scholar] [CrossRef] [Green Version]
- Hegazy, W.A.H.; Abbas, H.A. Evaluation of the role of SsaV ‘Salmonella pathogenicity island-2 dependent type III secretion system components on the virulence behavior of Salmonella enterica serovar Typhimurium. Afr. J. Biotechnol. 2017, 16, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Coulthurst, S.J.; Williamson, N.R.; Harris, A.K.P.; Spring, D.R.; Salmond, G.P.C. Metabolic and regulatory engineering of Serratia marcescens: Mimicking phage-mediated horizontal acquisition of antibiotic biosynthesis and quorum-sensing capacities. Microbiology 2006, 152, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.D.F.F.; Caldana, M.L.d.S.T.; de Andrade Moreira Machado, M.A.; Buzalaf, M.A.R. Use of xylitol for prevention of acute otitis media. Arq. Int. Otorrinolaringol. 2009, 13, 87–92. [Google Scholar]
- Da Silva, A.F.S.S.; Oliveira, M.G.; Raposo, N.R.B. Inibição in vitro da aderência de enteropatógenos pelo xilitol. Rev. Ciênc. Méd. Biol. 2010, 9, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.F.D.S.E.; Ferreira, A.S.; Oliveira, M.S.; Silva, S.S.D.; Raposo, N.R.B. In vitro inhibition of adhesion of Escherichia coli strains by Xylitol. Braz. Arch. Biol. Technol. 2011, 54, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Kirov, S.M. Bacteria that express lateral flagella enable dissection of the multifunctional roles of flagella in pathogenesis. Fems Microbiol. Lett. 2003, 224, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Rice, S.A.; Koh, K.S.; Queck, S.Y.; Labbate, M.; Lam, K.W.; Kjelleberg, S. Biofilm formation and sloughing in Serratia marcescens are controlled by quorum sensing and nutrient cues. J. Bacteriol. 2005, 187, 3477–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Kida, Y.; Inoue, H.; Shimizu, T.; Kuwano, K. Serratia marcescens serralysin induces inflammatory responses through protease-activated receptor 2. Infect. Immun. 2007, 75, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, W.L.; Leonard, M.T.; Fajardo-Cavazos, P.; Panayotova, N.; Farmerie, W.G.; Triplett, E.W.; Schuerger, A.C. Complete Genome Sequence of Serratia liquefaciens Strain ATCC 27592. Genome Announc. 2013, 1, e00548-13. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Nagaya, Y.; Pradel, E.; Ooka, T.; Ogura, Y.; Katsura, K.; Kurokawa, K.; Oshima, K.; Hattori, M.; Parkhill, J.; et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol. Evol. 2014, 6, 2096–2110. [Google Scholar] [CrossRef] [Green Version]
- Liaw, S.J.; Lai, H.C.; Ho, S.W.; Luh, K.T.; Wang, W.B. Role of RsmA in the regulation of swarming motility and virulence factor expression in Proteus mirabilis. J. Med. Microbiol. 2003, 52, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Horng, J.T.; Yang, C.H.; Tsai, Y.H.; Su, L.H.; Wei, C.F.; Chen, C.C.; Hsieh, S.C.; Lu, C.C.; Lai, H.C. RssAB-FlhDC-ShlBA as a major pathogenesis pathway in Serratia marcescens. Infect. Immun. 2010, 78, 4870–4881. [Google Scholar] [CrossRef] [Green Version]
- Askoura, M.; Youns, M.; Halim Hegazy, W.A. Investigating the influence of iron on Campylobacter jejuni transcriptome in response to acid stress. Microb. Pathog. 2020, 138, 103777. [Google Scholar] [CrossRef]
- Wei, C.F.; Tsai, Y.H.; Tsai, S.H.; Lin, C.S.; Chang, C.J.; Lu, C.C.; Huang, H.C.; Lai, H.C. Cross-talk between bacterial two-component systems drives stepwise regulation of flagellar biosynthesis in swarming development. Biochem. Biophys. Res. Commun. 2017, 489, 70–75. [Google Scholar] [CrossRef]
- Stella, N.A.; Fender, J.E.; Lahr, R.M.; Kalivoda, E.J.; Shanks, R.M. The LysR Transcription Factor, HexS, Is Required for Glucose Inhibition of Prodigiosin Production by Serratia marcescens. Adv. Microbiol. 2012, 2, 10.4236/aim.2012.24065. [Google Scholar] [CrossRef] [Green Version]
- Brothers, K.M.; Stella, N.A.; Romanowski, E.G.; Kowalski, R.P.; Shanks, R.M. EepR Mediates Secreted-Protein Production, Desiccation Survival, and Proliferation in a Corneal Infection Model. Infect. Immun. 2015, 83, 4373–4382. [Google Scholar] [CrossRef] [Green Version]
- Reboud, E.; Bouillot, S.; Patot, S.; Beganton, B.; Attree, I.; Huber, P. Pseudomonas aeruginosa ExlA and Serratia marcescens ShlA trigger cadherin cleavage by promoting calcium influx and ADAM10 activation. PLoS Pathog. 2017, 13, e1006579. [Google Scholar] [CrossRef]
- Shanks, R.M.; Stella, N.A.; Brothers, K.M.; Polaski, D.M. Exploitation of a “hockey-puck” phenotype to identify pilus and biofilm regulators in Serratia marcescens through genetic analysis. Can. J. Microbiol. 2016, 62, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Shanks, R.M.; Stella, N.A.; Kalivoda, E.J.; Doe, M.R.; O’Dee, D.M.; Lathrop, K.L.; Guo, F.L.; Nau, G.J. A Serratia marcescens OxyR homolog mediates surface attachment and biofilm formation. J. Bacteriol. 2007, 189, 7262–7272. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Xu, Y.; Xu, W.; Liao, W.; Xu, C.; Zhang, X.; Cao, J.; Zhou, T. Hypertonic glucose inhibits growth and attenuates virulence factors of multidrug-resistant Pseudomonas aeruginosa. BMC Microbiol. 2020, 20, 203. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | H-Bonding | Hydrophobic Interaction | Autodock Score |
|---|---|---|---|
| Xylitol | Arg 50, Tyr 57, Leu 69 | Ala 32, Tyr 33, Ala 34, Ile 46, Asp 66, Val 122 | −4.5 |
| C4-HSL | Arg 50, Trp 53, Asp 66, Thr 110 | Val 68, Leu 69, Trp 81, Phe 97, Ala 100, Ile 105, Val 122, Ser 124, Met 126 | −6.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khayyat, A.N.; Hegazy, W.A.H.; Shaldam, M.A.; Mosbah, R.; Almalki, A.J.; Ibrahim, T.S.; Khayat, M.T.; Khafagy, E.-S.; Soliman, W.E.; Abbas, H.A. Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens. Microorganisms 2021, 9, 1083. https://doi.org/10.3390/microorganisms9051083
Khayyat AN, Hegazy WAH, Shaldam MA, Mosbah R, Almalki AJ, Ibrahim TS, Khayat MT, Khafagy E-S, Soliman WE, Abbas HA. Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens. Microorganisms. 2021; 9(5):1083. https://doi.org/10.3390/microorganisms9051083
Chicago/Turabian StyleKhayyat, Ahdab N., Wael A. H. Hegazy, Moataz A. Shaldam, Rasha Mosbah, Ahmad J. Almalki, Tarek S. Ibrahim, Maan T. Khayat, El-Sayed Khafagy, Wafaa E. Soliman, and Hisham A. Abbas. 2021. "Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens" Microorganisms 9, no. 5: 1083. https://doi.org/10.3390/microorganisms9051083
APA StyleKhayyat, A. N., Hegazy, W. A. H., Shaldam, M. A., Mosbah, R., Almalki, A. J., Ibrahim, T. S., Khayat, M. T., Khafagy, E. -S., Soliman, W. E., & Abbas, H. A. (2021). Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens. Microorganisms, 9(5), 1083. https://doi.org/10.3390/microorganisms9051083