
Isolation and Characterization of Strain Exiguobacterium sp. KRL4, a Producer of Bioactive Secondary Metabolites from a Tibetan Glacier

 ,
,  , ,
, ,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Strain and Characterization
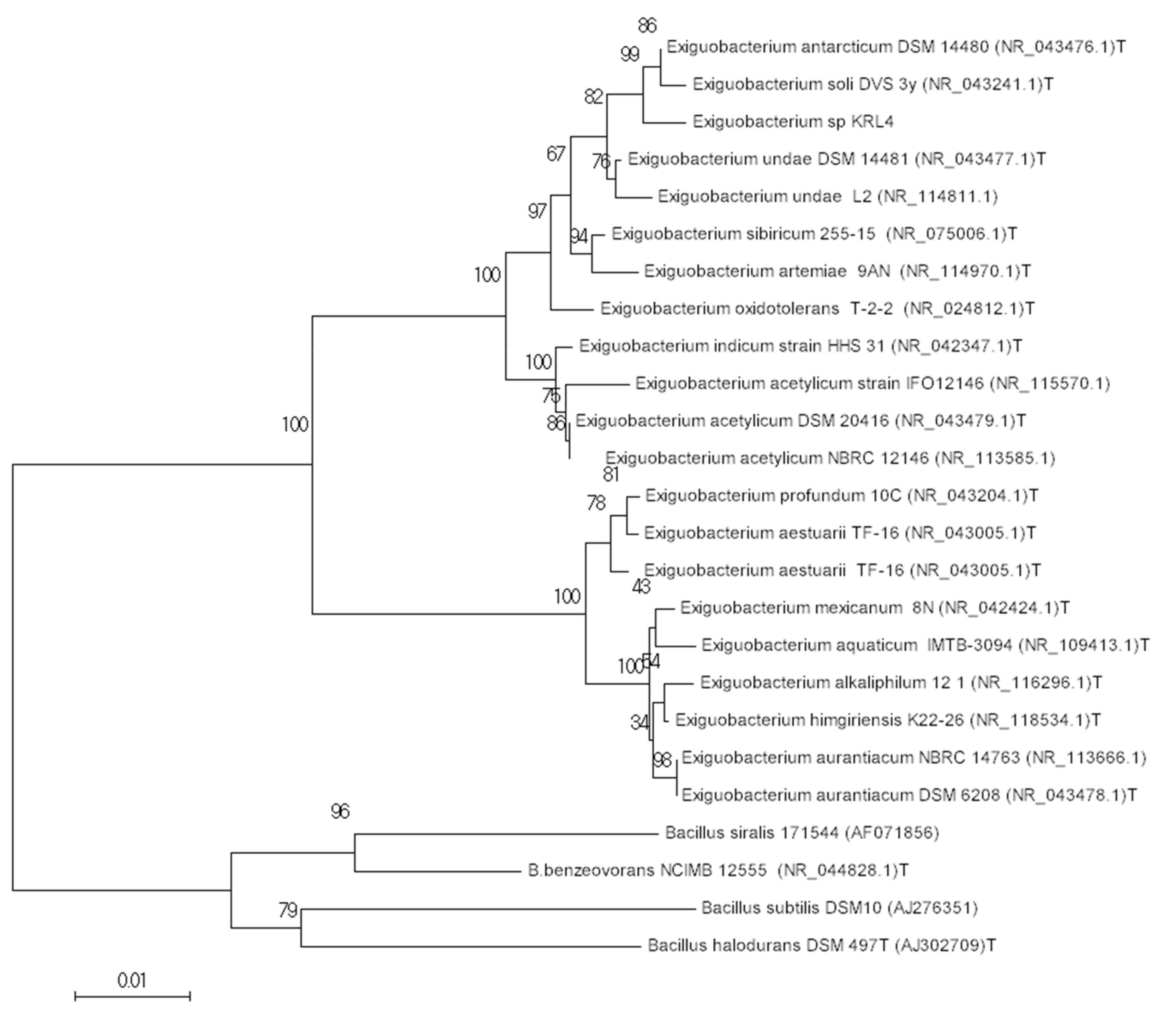
2.2. Bacterial Identification
2.3. Genome Sequencing and Analysis.
2.4. Membrane Composition Analysis
2.5. Bioactivity Assays
2.5.1. Preparation of Crude Extracts
2.5.2. Minimal Inhibitory Concentration Assay (MIC)
2.5.3. Nematode Liquid Toxicity Assay
2.5.4. DPPH Radical Scavenging Assay
2.5.5. Blue Agar CAS Assay for Siderophore Activity
3. Results and Discussion
3.1. Isolation and Characterization of Strain KRL4

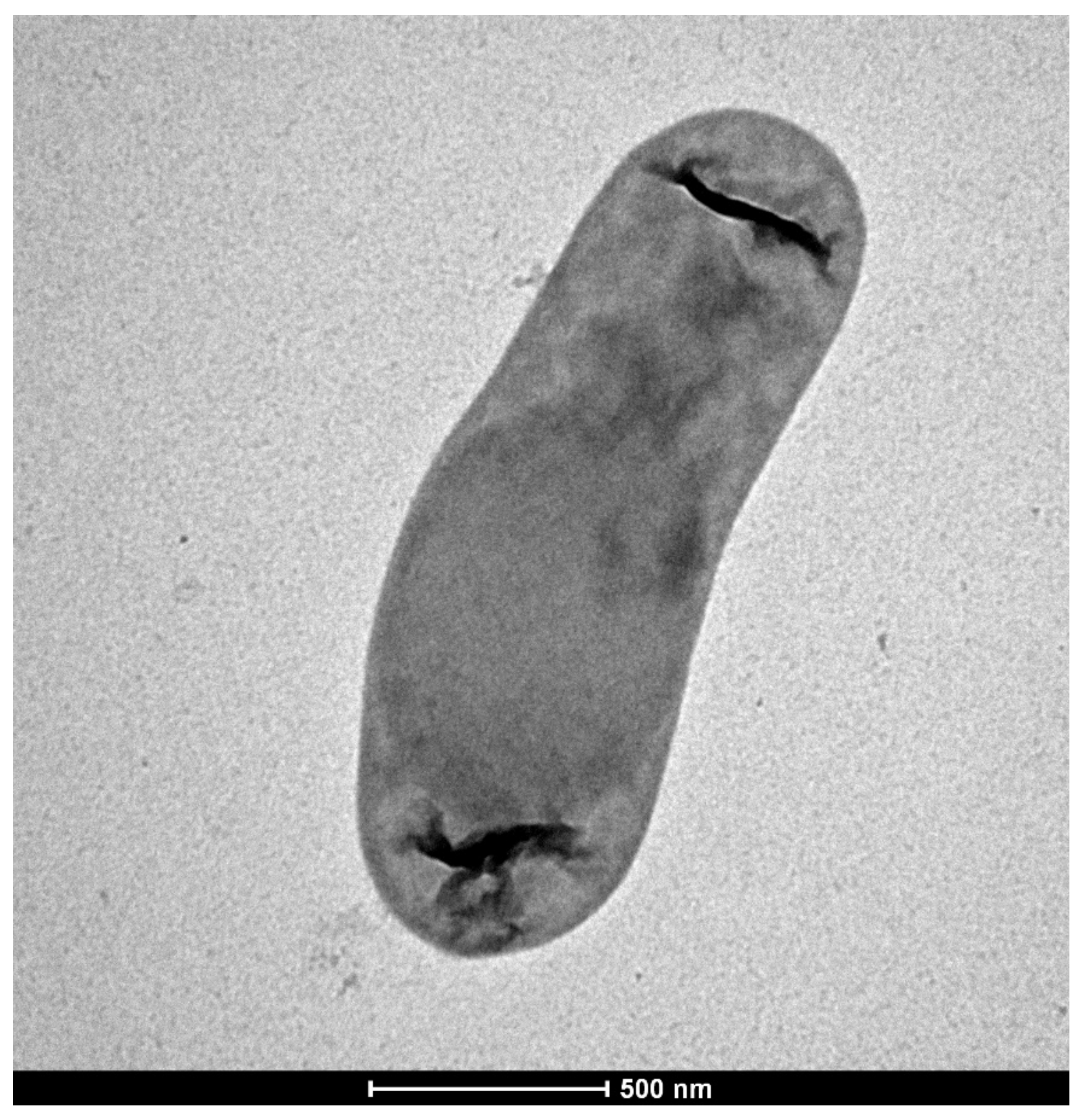{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Term | Evidence Code a |
|---|---|---|
| Classification | Domain Bacteria | TAS [51] |
| Phylum Firmicutes | TAS [52] | |
| Class Bacilli | TAS [48] | |
| Order Bacillales | TAS [53] | |
| Family Bacillales_Incertae Sedis XII | TAS [53] | |
| Genus Exiguobacterium_A | TAS [54] | |
| Positive | IDA | |
| Gram stain | Rod | IDA |
| Cell shape | non-motile | IDA |
| Motility | Not reported | NAS |
| Sporulation | 4–40 °C | IDA |
| Temperature range | 30 °C | IDA |
| Optimum temperature | 5.5–8.5; 6 | IDA |
| pH range; Optimum | D-mannose, Dextrin, D-fructose, sucrose D-maltose, α-D-glucose, D-lactose, D-sorbitol, D-galattosio, Lactulose, Dextrine, glycerol L-hystidine, L-cysteine, Xylan | IDA |
| Carbon source | Soil, sediment | IDA |
| Habitat | 0–15 % NaCl (w/v) | IDA |
| Salinity | Aerobic | IDA |
| Oxygen requirement | free-living | IDA |
| Biotic relationship | non-pathogen | NAS |
| Pathogenicity | China/Tibet | |
| Geographic location | August 2011 | |
| Sample collection | 90°19.23′ E | |
| Latitude | 28°90.71′ N | |
| Longitude | 5200 m | |
| Altitude |
3.2. Genome Properties and Identification of Isolate KRL4
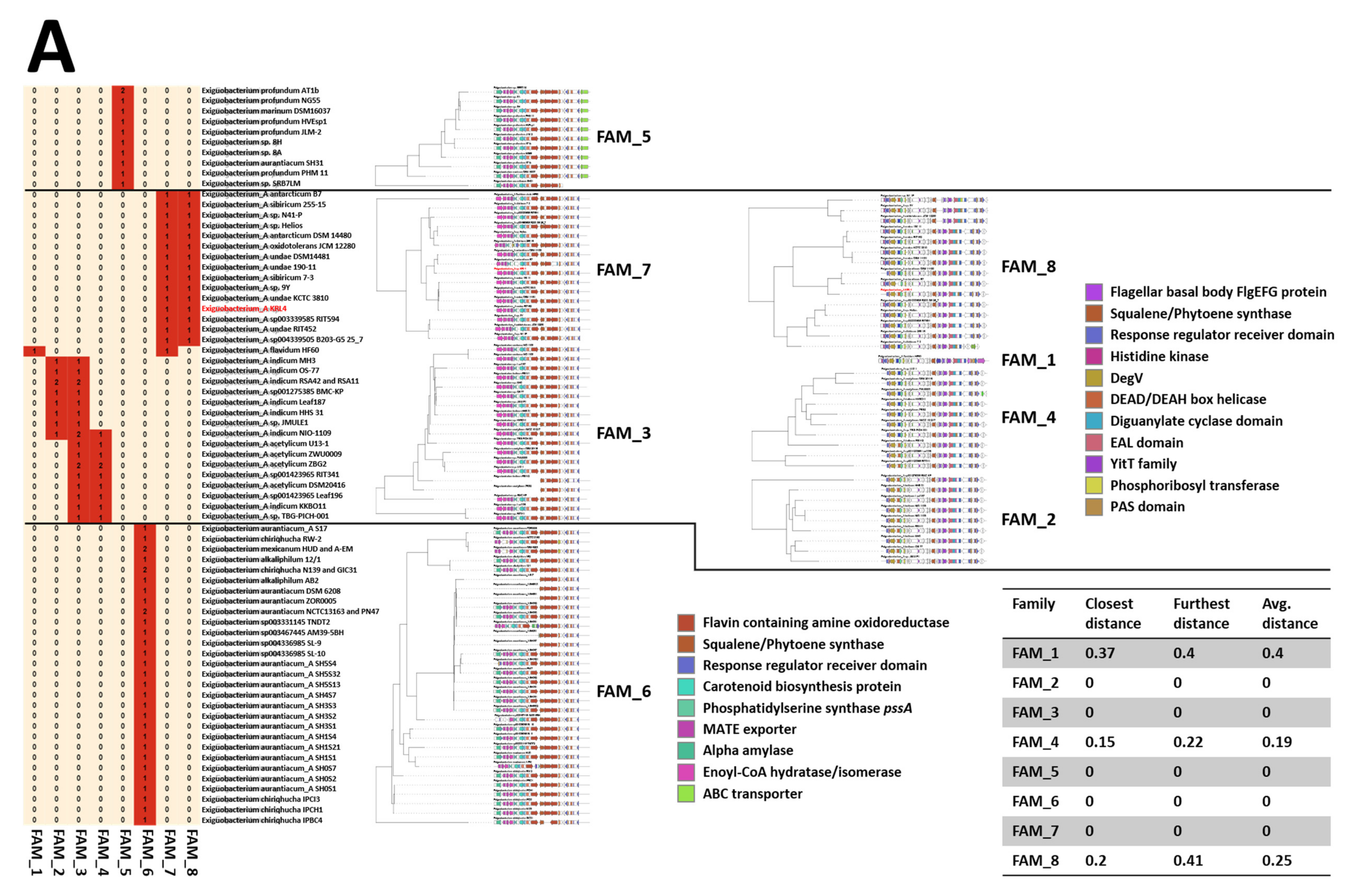
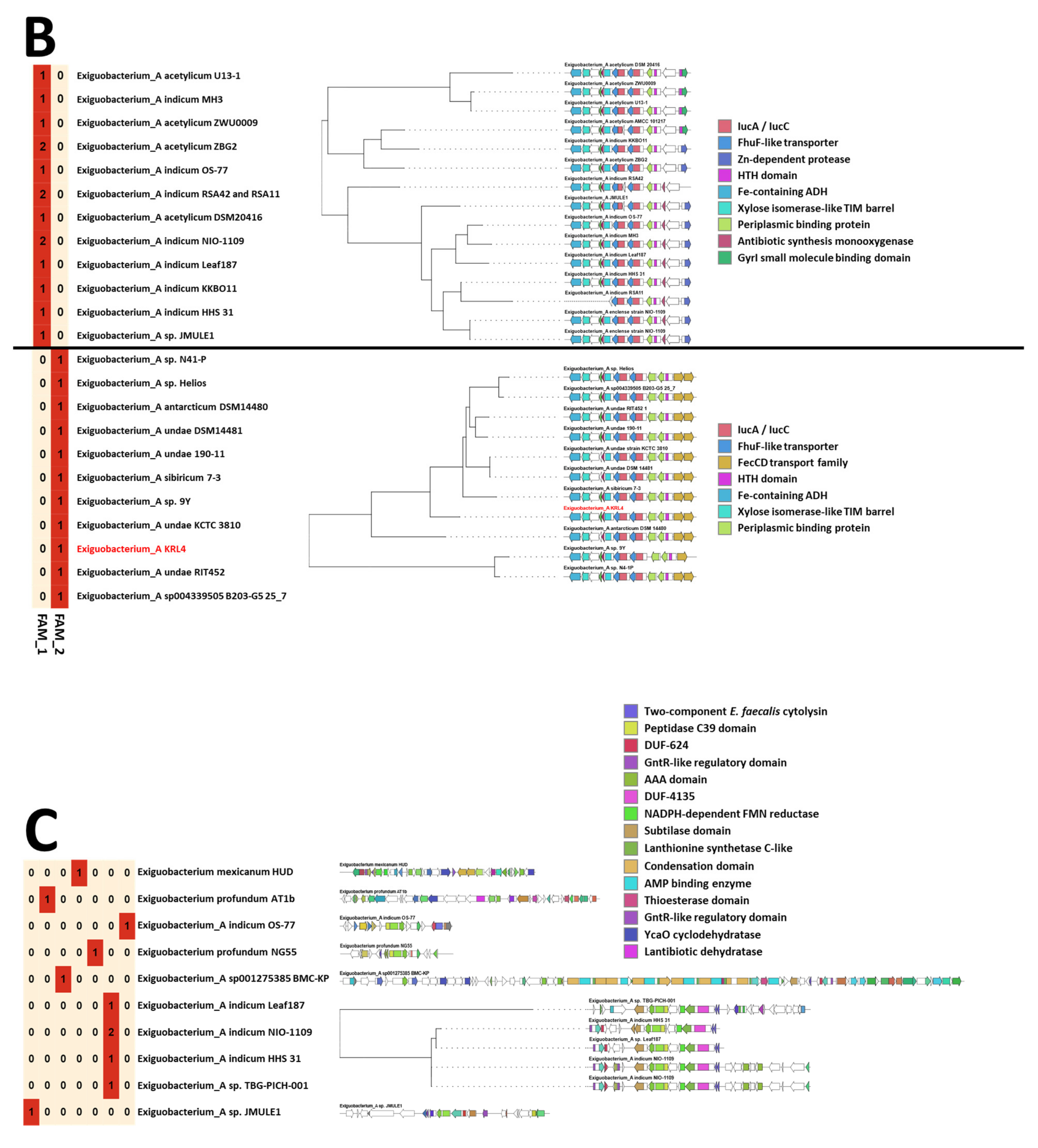
3.3. Genomic Insights of Strain KRL4
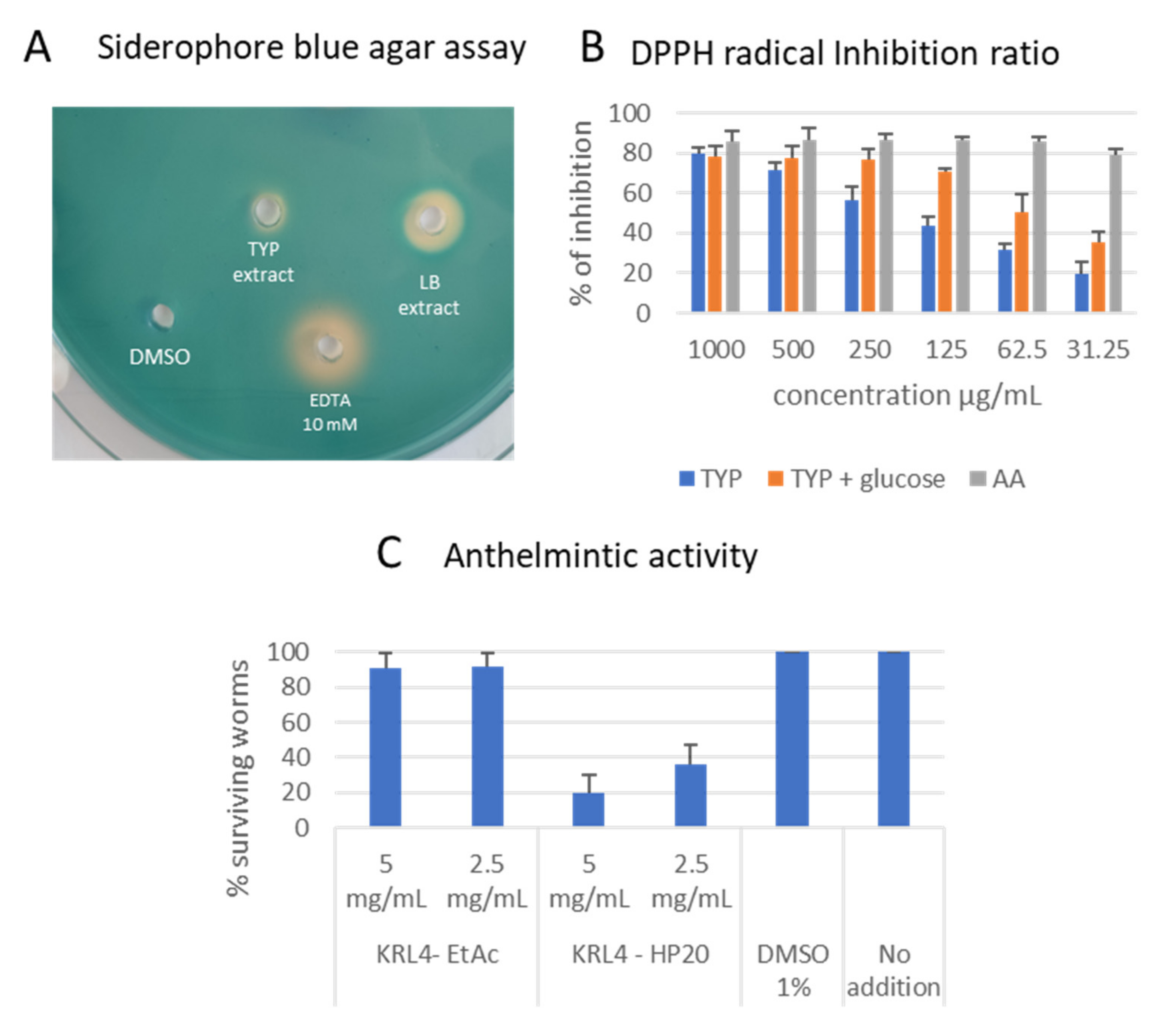
3.4. Bioactivity Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, M.; Lu, J.J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect 2021, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Opportunities for natural products in 21(st) century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine Microbial-Derived Molecules and Their Potential Use in Cosmeceutical and Cosmetic Products. Mar. Drugs 2017, 15, 118. [Google Scholar] [CrossRef]
- Teijaro, C.N.; Adhikari, A.; Shen, B. Challenges and opportunities for natural product discovery, production, and engineering in native producers versus heterologous hosts. J. Ind Microbiol Biotechnol. 2019, 46, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.H.; Kim, J. Cultivation of unculturable soil bacteria. Trends Biotechnol. 2012, 30, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Omar, S.M.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pascale, D.; De Santi, C.; Fu, J.; Landfald, B. The microbial diversity of Polar environments is a fertile ground for bioprospecting. Mar. Genom. 2012, 8, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Romoli, R.; Bartolucci, G.; Maida, I.; Perrin, E.; Fondi, M.; Orlandini, V.; Mengoni, A.; Emiliani, G.; Tutino, M.L.; et al. Bioactive volatile organic compounds from Antarctic (sponges) bacteria. N Biotechnol 2013, 30, 824–838. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Vrenna, G.; Lauro, C.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E.; Selan, L. Cold-adapted bacterial extracts as a source of anti-infective and antimicrobial compounds against Staphylococcus aureus. Future Microbiol. 2019, 14, 1369–1382. [Google Scholar] [CrossRef]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Tortorella, E.; Tedesco, P.; Palma Esposito, F.; January, G.G.; Fani, R.; Jaspars, M.; De Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. [Google Scholar] [CrossRef] [Green Version]
- Zain ul Arifeen, M.; Ma, Y.-N.; Xue, Y.-R.; Liu, C.-H. Deep-Sea Fungi Could Be the New Arsenal for Bioactive Molecules. Mar. Drugs 2020, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-N.; Meng, L.-H.; Wang, B.-G. Progress in Research on Bioactive Secondary Metabolites from Deep-Sea Derived Microorganisms. Mar. Drugs 2020, 18, 614. [Google Scholar] [CrossRef]
- Hemala, L.; Zhang, D.; Margesin, R. Cold-active antibacterial and antifungal activities and antibiotic resistance of bacteria isolated from an alpine hydrocarbon-contaminated industrial site. Res. Microbiol 2014, 165, 447–456. [Google Scholar] [CrossRef]
- Lv, Y.L.; Zhang, F.S.; Chen, J.; Cui, J.L.; Xing, Y.M.; Li, X.D.; Guo, S.X. Diversity and antimicrobial activity of endophytic fungi associated with the alpine plant Saussurea involucrata. Biol Pharm Bull. 2010, 33, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Azzoni, R.S.; Tagliaferri, I.; Franzetti, A.; Mayer, C.; Lambrecht, A.; Compostella, C.; Caccianiga, M.; Minora, U.F.; Garzonio, C.A.; Meraldi, E.; et al. Bacterial diversity in snow from mid-latitude mountain areas: Alps, Eastern Anatolia, Karakoram and Himalaya. Ann. Glaciol. 2018, 59, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Berger, T.; Poyntner, C.; Margesin, R. Culturable bacteria from an Alpine coniferous forest site: Biodegradation potential of organic polymers and pollutants. Folia Microbiol. 2021, 66, 87–98. [Google Scholar] [CrossRef]
- Rehakova, K.; Capkova, K.; Hrouzek, P.; Koblizek, M.; Dolezal, J. Microbial photosynthetic and photoprotective pigments in Himalayan soils originating from different elevations and successional stages. Soil Biol. Biochem. 2019, 132, 153–164. [Google Scholar] [CrossRef]
- Hei, Y.; Zhang, H.; Tan, N.; Zhou, Y.; Wei, X.; Hu, C.; Liu, Y.; Wang, L.; Qi, J.; Gao, J.M. Antimicrobial activity and biosynthetic potential of cultivable actinomycetes associated with Lichen symbiosis from Qinghai-Tibet Plateau. Microbiol Res. 2021, 244, 126652. [Google Scholar] [CrossRef]
- Chen, L. Complete genome sequence of Bacillus velezensis LM2303, a biocontrol strain isolated from the dung of wild yak inhabited Qinghai-Tibet plateau. J. Biotechnol 2017, 251, 124–127. [Google Scholar] [CrossRef]
- Wei, S.; Cui, H.; He, H.; Hu, F.; Su, X.; Zhu, Y. Diversity and Distribution of Archaea Community along a Stratigraphic Permafrost Profile from Qinghai-Tibetan Plateau, China. Archaea 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Li, Z.; Li, H.; Zhang, Z.; Xu, L.; Yue, X. Glaciers in Xinjiang, China: Past Changes and Current Status. 2020, 12, 2367. Water 2020, 12, 2367. [Google Scholar]
- Grifoni, A.; Bazzicalupo, M.; Di Serio, C.; Fancelli, S.; Fani, R. Identification of Azospirillum strains by restriction fragment length polymorphism of the 16S rDNA and of the histidine operon. FEMS Microbiol. Lett. 1995, 127, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Van Domselaar, G.H.; Stothard, P.; Shrivastava, S.; Cruz, J.A.; Guo, A.; Dong, X.; Lu, P.; Szafron, D.; Greiner, R.; Wishart, D.S. BASys: A web server for automated bacterial genome annotation. Nucleic Acids Res. 2005, 33, W455–W459. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting Secretory Proteins with SignalP. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.-A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A complete domain-to-species taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1079–1086. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework to explore large-scale biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Rodriguez-R, L.; Konstantinidis, K. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. Peer J. Prepr 2016, 4, e1900v1. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A fast phage search tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- De Jong, A.; van Hijum, S.A.; Bijlsma, J.J.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef]
- Soriano, F.; Huelves, L.; Naves, P.; Rodríguez-Cerrato, V.; del Prado, G.; Ruiz, V.; Ponte, C. In vitro activity of ciprofloxacin, moxifloxacin, vancomycin and erythromycin against planktonic and biofilm forms of Corynebacterium urealyticum. J. Antimicrob. Chemother. 2009, 63, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Shahi, S.K.; Kumar, A. Isolation and Genetic Analysis of Multidrug Resistant Bacteria from Diabetic Foot Ulcers. Front. Microbiol. 2016, 6, 1464. [Google Scholar] [CrossRef] [Green Version]
- Alanazi, S.A.; Qureshi, K.A.; Elhassan, G.O.; E, I.E.-A. Isolation, Purification and Characterization of Antimicrobial Agent Antagonistic to Escherichia coli ATCC 10536 Produced by Bacillus pumilus SAFR-032 Isolated from the Soil of Unaizah, Al Qassim Province of Saudi Arabia. Pak. J. Biol. Sci. : PJBS 2016, 19, 191–201. [Google Scholar] [CrossRef]
- Werbrouck, H.; Vermeulen, A.; Van Coillie, E.; Messens, W.; Herman, L.; Devlieghere, F.; Uyttendaele, M. Influence of acid stress on survival, expression of virulence genes and invasion capacity into Caco-2 cells of Listeria monocytogenes strains of different origins. Int J. Food Microbiol. 2009, 134, 140–146. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Lima, D.B.; Torres, A.F.; Mello, C.P.; de Menezes, R.R.; Sampaio, T.L.; Canuto, J.A.; da Silva, J.J.; Freire, V.N.; Quinet, Y.P.; Havt, A.; et al. Antimicrobial effect of Dinoponera quadriceps (Hymenoptera: Formicidae) venom against Staphylococcus aureus strains. J. Applied Microbiol. 2014, 117, 390–396. [Google Scholar] [CrossRef]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; Deboy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ebada, S.S.; Edrada, R.A.; Lin, W.; Proksch, P. Methods for isolation, purification and structural elucidation of bioactive secondary metabolites from marine invertebrates. Nat. Protoc. 2008, 3, 1820–1831. [Google Scholar] [CrossRef] [PubMed]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Kasana, R.C.; Pandey, C.B. Exiguobacterium: An overview of a versatile genus with potential in industry and agriculture. Crit Rev. Biotechnol. 2018, 38, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Remonsellez, F.; Castro-Severyn, J.; Pardo-Esté, C.; Aguilar, P.; Fortt, J.; Salinas, C.; Barahona, S.; León, J.; Fuentes, B.; Areche, C.; et al. Characterization and Salt Response in Recurrent Halotolerant Exiguobacterium sp. SH31 Isolated From Sediments of Salar de Huasco, Chilean Altiplano. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- White, R.A.; Soles, S.A.; Gavelis, G.; Gosselin, E.; Slater, G.F.; Lim, D.S.S.; Leander, B.; Suttle, C.A. The Complete Genome and Physiological Analysis of the Eurythermal Firmicute Exiguobacterium chiriqhucha Strain RW2 Isolated From a Freshwater Microbialite, Widely Adaptable to Broad Thermal, pH, and Salinity Ranges. Front. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W. Bergey’s Manual of Systematic Bacteriology - Volume 3: The Firmicutes; The Firmicutes; Springer: New York, NY, USA, 2009. [Google Scholar]
- Mykytczuk, N.C.S.; Foote, S.J.; Omelon, C.R.; Southam, G.; Greer, C.W.; Whyte, L.G. Bacterial growth at −15 °C; molecular insights from the permafrost bacterium Planococcus halocryophilus Or1. The ISME J. 2013, 7, 1211–1226. [Google Scholar] [CrossRef] [Green Version]
- Sleight, S.C.; Orlic, C.; Schneider, D.; Lenski, R.E. Genetic basis of evolutionary adaptation by Escherichia coli to stressful cycles of freezing, thawing and growth. Genetics 2008, 180, 431–443. [Google Scholar] [CrossRef]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [Green Version]
- Gibbonst, N.E.; Murray, R.G.E. Proposals Concerning the Higher Taxa of Bacteria. Int. J. Syst. Bacteriol. 1978, 28, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Skerman, V.B.D.; McGowan, V.; Sneath, P.H.A. (Eds.) Approved Lists of Bacterial Names (Amended); ASM Press Copyright © 2021, ASM Press.: Washington, DC, USA, 1989. [Google Scholar]
- Collins, M.D.; Lund, B.M.; Farrow, J.A.E.; Schleifer, K.H. Chemotaxonomic Study of an Alkalophilic Bacterium, Exiguobacterium aurantiacum gen. nov., sp. nov. J. General Microbiol. 1983, 129, 2037–2043. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nature Genetics 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishnivetskaya, T.A.; Kathariou, S.; Tiedje, J.M. The Exiguobacterium genus: Biodiversity and biogeography. Extremophiles. 2009, 13, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. CONFIDENCE LIMITS ON PHYLOGENIES: AN APPROACH USING THE BOOTSTRAP. Evolution. 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Iwata-Reuyl, D.; Math, S.K.; Desai, S.B.; Poulter, C.D. Bacterial Phytoene Synthase: Molecular Cloning, Expression, and Characterization of Erwinia herbicola Phytoene Synthase. Biochemistry 2003, 42, 3359–3365. [Google Scholar] [CrossRef]
- Ueki, N.; Ide, T.; Mochiji, S.; Kobayashi, Y.; Tokutsu, R.; Ohnishi, N.; Yamaguchi, K.; Shigenobu, S.; Tanaka, K.; Minagawa, J.; et al. Eyespot-dependent determination of the phototactic sign in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 5299–5304. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Tsukamoto, T.; Sudo, Y. Molecular and evolutionary aspects of microbial sensory rhodopsins. Biochimica et Biophysica Acta (BBA) - Bioenerg. 2014, 1837, 562–577. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Chen, Z.; Hou, J.; Wang, Y.; Guo, M.; Cao, J.; Wang, L.; Xu, H.; Tian, B.; Zhao, Y. MazEF Toxin-Antitoxin System-Mediated DNA Damage Stress Response in Deinococcus radiodurans. Front. Genet. 2021, 12. [Google Scholar] [CrossRef]
- Heath, L.S.; Heath, H.E.; LeBlanc, P.A.; Smithberg, S.R.; Dufour, M.; Simmonds, R.S.; Sloan, G.L. The streptococcolytic enzyme zoocin A is a penicillin-binding protein. FEMS Microbiol. Letters 2004, 236, 205–211. [Google Scholar] [CrossRef]
- Albarracín, V.H.; Kraiselburd, I.; Bamann, C.; Wood, P.G.; Bamberg, E.; Farias, M.E.; Gärtner, W. Functional Green-Tuned Proteorhodopsin from Modern Stromatolites. PLOS ONE 2016, 11, e0154962. [Google Scholar] [CrossRef]
- Gushchin, I.; Chervakov, P.; Kuzmichev, P.; Popov, A.N.; Round, E.; Borshchevskiy, V.; Ishchenko, A.; Petrovskaya, L.; Chupin, V.; Dolgikh, D.A.; et al. Structural insights into the proton pumping by unusual proteorhodopsin from nonmarine bacteria. Proc. Natl. Acad. Sci. USA 2013, 110, 12631–12636. [Google Scholar] [CrossRef] [Green Version]
- Castro-Severyn, J.; Remonsellez, F.; Valenzuela, S.L.; Salinas, C.; Fortt, J.; Aguilar, P.; Pardo-Esté, C.; Dorador, C.; Quatrini, R.; Molina, F.; et al. Comparative Genomics Analysis of a New Exiguobacterium Strain from Salar de Huasco Reveals a Repertoire of Stress-Related Genes and Arsenic Resistance. Front. Microbiol 2017, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Li, J.; Zhang, C.; Tian, Z.; Liu, X.; Tang, C.; Han, Y.; Liu, W. Assessment of heavy metal contents in surface soil in the Lhasa-Shigatse-Nam Co area of the Tibetan Plateau, China. Bull. Environ. Contam. Toxicol. 2014, 93, 192–198. [Google Scholar] [CrossRef]
- Paez-Espino, D.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Thomas, A.D.; Huntemann, M.; Mikhailova, N.; Rubin, E.; Ivanova, N.N.; Kyrpides, N.C. Uncovering Earth’s virome. Nature 2016, 536, 425–430. [Google Scholar] [CrossRef]
- Pace, S.; Ceballos, S.J.; Harrold, D.; Stannard, W.; Simmons, B.A.; Singer, S.W.; Thelen, M.P.; VanderGheynst, J.S. Enrichment of microbial communities tolerant to the ionic liquids tetrabutylphosphonium chloride and tributylethylphosphonium diethylphosphate. Applied Microbiol. Biotechnol. 2016, 100, 5639–5652. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.; Holmström, S.J. Siderophores in environmental research: Roles and applications. Microb Biotechnol 2014, 7, 196–208. [Google Scholar] [CrossRef]
- Rajendran, G.; Patel, M.H.; Joshi, S.J. Isolation and Characterization of Nodule-Associated Exiguobacterium sp. from the Root Nodules of Fenugreek (Trigonella foenum-graecum) and Their Possible Role in Plant Growth Promotion. Int. J. Microbiol. 2012, 2012, 693982. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Bioprospecting of plant growth promoting psychrotrophic Bacilli from the cold desert of north western Indian Himalayas. Indian J. Exp. Boil. 2016, 54, 142–150. [Google Scholar]
- Jinendiran, S.; Dahms, H.-U.; Dileep Kumar, B.S.; Kumar Ponnusamy, V.; Sivakumar, N. Diapolycopenedioic-acid-diglucosyl ester and keto-myxocoxanthin glucoside ester: Novel carotenoids derived from Exiguobacterium acetylicum S01 and evaluation of their anticancer and anti-inflammatory activities. . Bioorgan. Chem. 2020, 103, 104149. [Google Scholar] [CrossRef]
- Chellaram, C.; Praveen, M.M.; John, A.A. Antimicrobial activity of an epibiotic Exiguobacterium strain SGAA-2 from Gorgonian coral, Subergorgia suberosa (Pallas, 1766). Indian J. Geo-Mar. Sci. 2015, 43, 750–755. [Google Scholar]
- Shanthakumar, S.P.; Duraisamy, P.; Vishwanath, G.; Selvanesan, B.C.; Ramaraj, V.; Vasantharaj David, B. Broad spectrum antimicrobial compounds from the bacterium Exiguobacterium mexicanum MSSRFS9. Microbiol Res. 2015, 178, 59–65. [Google Scholar] [CrossRef]
- Mirza, Z.; Soto, E.R.; Hu, Y.; Nguyen, T.-T.; Koch, D.; Aroian, R.V.; Ostroff, G.R. Anthelmintic Activity of Yeast Particle-Encapsulated Terpenes. Molecules 2020, 25, 2958. [Google Scholar] [CrossRef]
- Holden-Dye, L.; Walker, R.J. Anthelmintic drugs. WormBook 2007, 1–13. [Google Scholar] [CrossRef]
- Garcia-Bustos, J.F.; Sleebs, B.E.; Gasser, R.B. An appraisal of natural products active against parasitic nematodes of animals. Parasites Vectors 2019, 12, 306. [Google Scholar] [CrossRef] [Green Version]
- Xiang, W.-S.; Wang, J.-D.; Wang, M.; Wang, X.-J. New nemadectin congener from Streptomyces microflavus neau3: Fermentation, isolation, structure elucidation and biological activities. J. Antibiot. 2010, 63, 171–175. [Google Scholar] [CrossRef]





| Attribute | Value | |
|---|---|---|
| Genome size (bp) | 3,118,075 | |
| DNA coding (bp) | 2,823,885 | |
| DNA G+C (bp) | 1,462,377 | |
| DNA scaffolds | 48 | |
| Total genes | 3201 | |
| Protein coding genes | 3079 | |
| RNA genes | 79 | |
| Pseudo genes | 43 | |
| Genes in internal clusters | ||
| Genes with function prediction | 1522 | |
| Genes assigned to COGs | 2052 | |
| Genes with Pfam domains | 2597 | |
| Genes with signal peptides | 146 | |
| Genes with transmembrane helices | 1207 | |
| CRISPR repeats | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedesco, P.; Palma Esposito, F.; Masino, A.; Vitale, G.A.; Tortorella, E.; Poli, A.; Nicolaus, B.; van Zyl, L.J.; Trindade, M.; de Pascale, D. Isolation and Characterization of Strain Exiguobacterium sp. KRL4, a Producer of Bioactive Secondary Metabolites from a Tibetan Glacier. Microorganisms 2021, 9, 890. https://doi.org/10.3390/microorganisms9050890
Tedesco P, Palma Esposito F, Masino A, Vitale GA, Tortorella E, Poli A, Nicolaus B, van Zyl LJ, Trindade M, de Pascale D. Isolation and Characterization of Strain Exiguobacterium sp. KRL4, a Producer of Bioactive Secondary Metabolites from a Tibetan Glacier. Microorganisms. 2021; 9(5):890. https://doi.org/10.3390/microorganisms9050890
Chicago/Turabian StyleTedesco, Pietro, Fortunato Palma Esposito, Antonio Masino, Giovanni Andrea Vitale, Emiliana Tortorella, Annarita Poli, Barbara Nicolaus, Leonardo Joaquim van Zyl, Marla Trindade, and Donatella de Pascale. 2021. "Isolation and Characterization of Strain Exiguobacterium sp. KRL4, a Producer of Bioactive Secondary Metabolites from a Tibetan Glacier" Microorganisms 9, no. 5: 890. https://doi.org/10.3390/microorganisms9050890
APA StyleTedesco, P., Palma Esposito, F., Masino, A., Vitale, G. A., Tortorella, E., Poli, A., Nicolaus, B., van Zyl, L. J., Trindade, M., & de Pascale, D. (2021). Isolation and Characterization of Strain Exiguobacterium sp. KRL4, a Producer of Bioactive Secondary Metabolites from a Tibetan Glacier. Microorganisms, 9(5), 890. https://doi.org/10.3390/microorganisms9050890