
The Effect of Antimicrobial Treatment upon the Gill Bacteriome of Atlantic Salmon (Salmo salar L.) and Progression of Amoebic Gill Disease (AGD) In Vivo



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Source and Husbandry
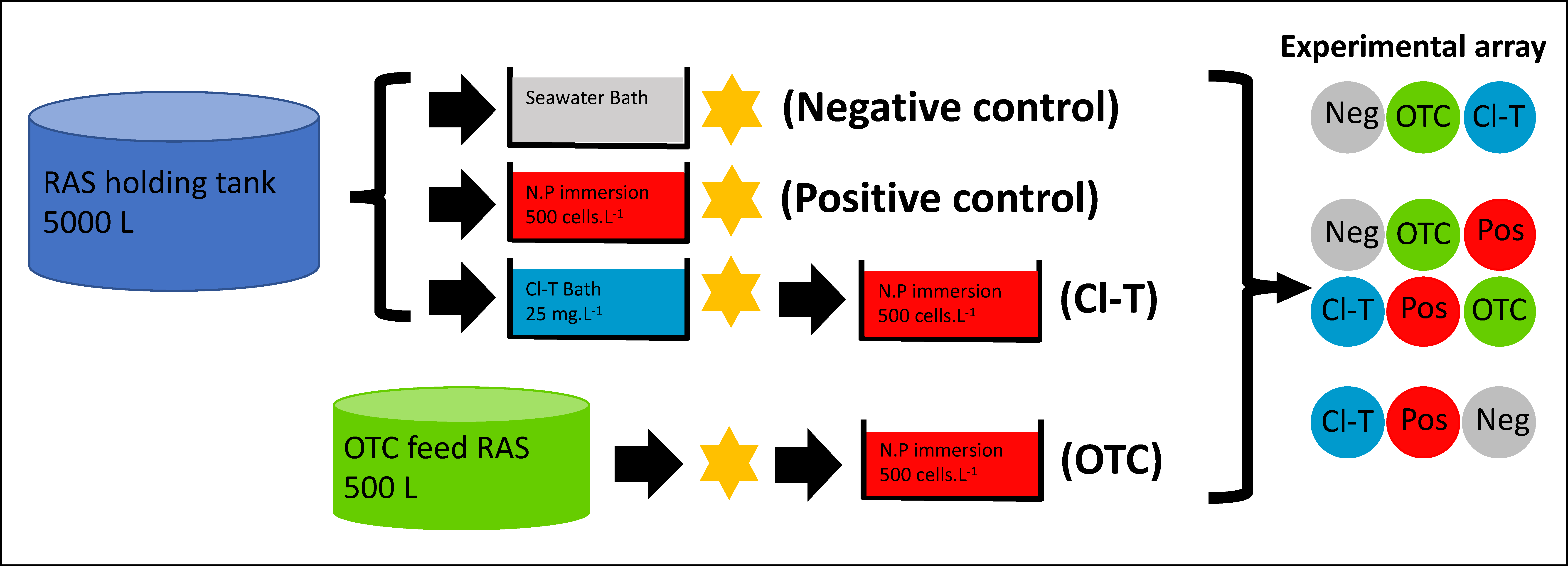
2.2. Experimental Design, Procedures and Maintenance
2.3. Pre-Challenge with Antimicrobial Treatments
2.4. Sampling Strategy
2.4.1. Gill Mucus Collection
2.4.2. Tank Water Samples
2.5. DNA Extraction and Purification
2.5.1. Gill Mucus
2.5.2. Tank Water
2.6. Challenge with Neoparamoeba Perurans and AGD Assessment
2.6.1. Gill Score Assessment
2.6.2. Gill Histopathology
2.6.3. Quantitative PCR Assay
2.7. Gill Bacteriome Assessment
2.7.1. Branchial Bacteria Counts
2.7.2. 16 S rRNA Amplicon Sequencing
2.8. Bioinformatics Pipeline
2.9. Statistical Analysis
3. Results
3.1. Onset and Progression of AGD after Challenge with N. perurans
3.2. Gill Bacteriomic Profiles
3.2.1. Following Antibacterial Treatment
3.2.2. Concurrent to Neoparamoeba Perurans Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef]
- de Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 2018, 94, 1–12. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and ecological roles of external fish mucus: A review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kessel, M.A.H.J.; Mesman, R.J.; Arshad, A.; Metz, J.R.; Spanings, F.A.T.; van Dalen, S.C.M.; van Niftrik, L.; Flik, G.; Wendelaar Bonga, S.E.; Jetten, M.S.M.; et al. Branchial nitrogen cycle symbionts can remove ammonia in fish gills. Environ. Microbiol. Rep. 2016, 8, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Cabillon, N.; Lazado, C. Mucosal barrier functions of fish under changing environmental conditions. Fishes 2019, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Minniti, G.; Hagen, L.H.; Porcellato, D.; Jørgensen, S.M.; Pope, P.B.; Vaaje-Kolstad, G. The skin-mucus microbial community of farmed Atlantic salmon (Salmo salar). Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.; Salinas, I. Under pressure: Interactions between commensal microbiota and the teleost immune system. Front. Immunol. 2017, 8, 559. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, J.; Ringø, E.; Merrifield, D.L. The gut microbiota of fish. In Aquaculture Nutrition; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 75–100. [Google Scholar]
- Moya, A.; Ferrer, M. Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends Microbiol. 2016, 24, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.; Gardiner, M. Microbial dysbiosis: Rethinking disease in marine ecosystems. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Webster, T.M.U.; Consuegra, S.; de Leaniz, C.G. Early life stress causes persistent impacts on the microbiome of Atlantic salmon. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Lokesh, J.; Kiron, V. Transition from freshwater to seawater reshapes the skin-associated microbiota of Atlantic salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llewellyn, M.S.; Leadbeater, S.; Garcia, C.; Sylvain, F.-E.; Custodio, M.; Ang, K.P.; Powell, F.; Carvalho, G.R.; Creer, S.; Elliot, J.; et al. Parasitism perturbs the mucosal microbiome of Atlantic Salmon. Sci. Rep. 2017, 7, 43465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Long, M.; Ji, C.; Shen, Z.; Gatesoupe, F.J.; Zhang, X.; Zhang, Q.; Zhang, L.; Zhao, Y.; Liu, X.; et al. Alterations of the gut microbiome of largemouth bronze gudgeon (Coreius guichenoti) suffering from furunculosis. Sci. Rep. 2016, 6, 30606. [Google Scholar] [CrossRef]
- Bowker, J.D.; Carty, D.G.; Telles, L.; David, B.; Oviedo, D. Efficacy of chloramine-T to control mortality in freshwater-reared salmonids diagnosed with bacterial gill disease. N. Am. J. Aquac. 2008, 70, 20–26. [Google Scholar] [CrossRef]
- Wynne, J.W.; Thakur, K.K.; Slinger, J.; Samsing, F.; Milligan, B.; Powell, J.F.F.; McKinnon, A.; Nekouei, O.; New, D.; Richmond, Z.; et al. Microbiome profiling reveals a microbial dysbiosis during a natural outbreak of Tenacibaculosis (yellow mouth) in atlantic salmon. Front. Microbiol. 2020, 11, 586387. [Google Scholar] [CrossRef]
- Kiemer, M.C.B.; Black, K.D. The effects of hydrogen peroxide on the gill tissues of Atlantic salmon, Salmo salar L. Aquaculture 1997, 153, 181–189. [Google Scholar] [CrossRef]
- Munday, B.L. Disease of salmonids. In Proceedings of the Workshop on Diseases in Australian Fish and Shellfish, Victoria Department of Agriculture, Melbourne, Australia, 27–30 May 1985; Humphrey, J.D., Langdon, J.S., Eds.; Australian Fish Health Reference Laboratory, Regional Veterinary Laboratory: Benalla, Australia, 1986; pp. 127–141. [Google Scholar]
- Adams, M.B.; Nowak, B.F. Pathology of amoebic gill disease in Atlantic salmon (Salmo salar L.). J. Fish Dis. 2003, 26, 601–614. [Google Scholar] [CrossRef]
- Roubal, F.R.; Lester, R.J.G.; Foster, C.K. Studies on cultured and gill-attached Paramoeba sp. (Gymnamoebae: Paramoebidae) and the cytopathology of paramoebic gill disease in Atlantic salmon, Salmo salar L., from Tasmania. J. Fish Dis. 1989, 12, 481–492. [Google Scholar] [CrossRef]
- Crosbie, P.B.B.; Bridle, A.R.; Cadoret, K.; Nowak, B.F. In vitro cultured Neoparamoeba perurans causes amoebic gill disease in Atlantic salmon and fulfils Koch’s postulates. Int. J. Parasitol. 2012, 42, 511–515. [Google Scholar] [CrossRef]
- Young, N.D.; Crosbie, P.B.B.; Adams, M.B.; Nowak, B.F.; Morrison, R.N. Neoparamoeba perurans n. sp., an agent of amoebic gill disease of Atlantic salmon (Salmo salar). Int. J. Parasitol. 2007, 37, 1469–1481. [Google Scholar] [CrossRef]
- Wiik-Nielsen, J.; Mo, T.A.; Kolstad, H.; Mohammad, S.N.; Hytterød, S.; Powell, M.D. Morphological diversity of Paramoeba perurans trophozoites and their interaction with Atlantic salmon, Salmo salar L., gills. J. Fish Dis. 2016, 39, 1113–1123. [Google Scholar] [CrossRef]
- Leef, M.J.; Harris, J.O.; Powell, M.D. Respiratory pathogenesis of amoebic gill disease (AGD) in experimentally infected Atlantic salmon Salmo salar. Dis. Aquat. Organ. 2005, 66, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.O.; Powell, M.D.; Attard, M.G.; DeHayr, L. Clinical assessment of chloramine-T and freshwater as treatments for the control of gill amoebae in Atlantic salmon, Salmo salar L. Aquac. Res. 2005, 36, 776–784. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hamlin-Wright, H.; Monaghan, S.; Herath, T.; Baily, J.; del Pozo, J.; Downes, J.; Preston, A.; Chalmers, L.; Jayasuriya, N.; et al. Changes in distribution, morphology and ultrastructure of chloride cell in Atlantic salmon during an AGD infection. J. Fish Dis 2019, 42, 1433–1446. [Google Scholar] [CrossRef]
- Munday, B.L.; Zilberg, D.; Findlay, V. Gill disease of marine fish caused by infection with Neoparamoeba pemaquidensis. J. Fish Dis. 2001, 24, 497–507. [Google Scholar] [CrossRef]
- Powell, M.D.; Leef, M.J.; Roberts, S.D.; Jones, M.A. Neoparamoebic gill infections: Host response and physiology in salmonids. J. Fish Biol. 2008, 73, 2161–2183. [Google Scholar] [CrossRef]
- Bridle, A.R.; Crosbie, P.B.B.; Cadoret, K.; Nowak, B.F. Rapid detection and quantification of Neoparamoeba perurans in the marine environment. Aquaculture 2010, 309, 56–61. [Google Scholar] [CrossRef]
- Wright, D.W.; Nowak, B.; Oppedal, F.; Bridle, A.; Dempster, T. Depth distribution of the amoebic gill disease agent, Neoparamoeba perurans, in salmon sea-cages. Aquac. Environ. Interact. 2015, 7, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Embar-Gopinath, S.; Butler, R.; Nowak, B. Influence of salmonid gill bacteria on development and severity of amoebic gill disease. Dis. Aquat. Organ. 2005, 67, 55–60. [Google Scholar] [CrossRef]
- Adams, M.B.; Ellard, K.; Nowak, B.F. Gross pathology and its relationship with histopathology of amoebic gill disease (AGD) in farmed Atlantic salmon, Salmo salar L. J. Fish Dis. 2004, 27, 151–161. [Google Scholar] [CrossRef]
- MacPhail, D.; Koppenstein, R.; Longshaw, M.; Henriquez, F. The bacterial microbiome of in vitro cultures of Paramoeba perurans. Access Microbiol. 2019. [Google Scholar] [CrossRef]
- Slinger, J.; Adams, M.B.; Wynne, J.W. Bacteriomic profiling of branchial lesions induced by neoparamoeba perurans challenge reveals commensal dysbiosis and an association with tenacibaculum dicentrarchi in agd-affected atlantic salmon (Salmo salar L.). Microorganisms 2020, 8, 1189. [Google Scholar] [CrossRef]
- Bowman, J.P.; Nowak, B. Salmonid gill bacteria and their relationship to amoebic gill disease. J. Fish Dis. 2004, 27, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Embar-Gopinath, S. Role of Bacteria in Amoebic Gill Disease. Doctoral Dissertation, University of Tasmania, Hobart, TAS, Australia, 2006. [Google Scholar]
- Galván-Moroyoqui, J.M.; del Carmen Domínguez-Robles, M.; Franco, E.; Meza, I. The interplay between Entamoeba and enteropathogenic bacteria modulates epithelial cell damage. PLoS Negl. Trop. Dis. 2008, 2, e266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badenoch, P.R.; Johnson, A.M.; Christy, P.E.; Coster, D.J. Pathogenicity of acanthamoeba and a corynebacterium in the rat cornea. Arch. Ophthalmol. 1990, 108, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.S.; Muller, W.J.; Cook, M.T.; Kube, P.D.; Elliott, N.G. Gill observations in Atlantic salmon (Salmo salar, L.) during repeated amoebic gill disease (AGD) field exposure and survival challenge. Aquaculture 2009, 290, 1–8. [Google Scholar] [CrossRef]
- Bruno, D.; Collet, B.; Turnbull, A.; Kilburn, R.; Walker, A.; Pendrey, D.; McIntosh, A.; Urquhart, K.; Taylor, G. Evaluation and development of diagnostic methods for Renibacterium salmoninarum causing bacterial kidney disease (BKD) in the UK. Aquaculture 2007, 269, 114–122. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857, Correction in 2019, 37, 1091. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quensen, J. QsRutils: R Functions Useful for Community Ecology. 2020. Available online: https://github.com/jfq3/QsRutils (accessed on 15 March 2021).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Arbizu, P.M. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis, R Package Version 0.3; 2019. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 10 March 2021).
- Ssekagiri, A.; TSloan, W.; Zeeshan Ijaz, U. microbiomeSeq: An R package for analysis of microbial communities in an environmental context. In Proceedings of the ISCB Africa ASBCB Conference, Kumasi, Ghana, 9 August 2017. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Embar-Gopinath, S.; Crosbie, P.B.B.; Nowak, B.F. Use of potassium permanganate to disinfect Atlantic salmon (Salmo salar) gills. Bull. Eur. Assoc. Fish Pathol. 2005, 25, 291–296. [Google Scholar]
- Carlson, J.M.; Leonard, A.B.; Hyde, E.R.; Petrosino, J.F.; Primm, T.P. Microbiome disruption and recovery in the fish Gambusia affinis following exposure to broad-spectrum antibiotic. Infect. Drug Resist. 2017, 10, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.D.; Perry, S.F. Cardio-respiratory effects of chloramine-T exposure in rainbow trout. Exp. Biol. Online 1999, 4, 1–59. [Google Scholar] [CrossRef]
- Powell, M.D.; Reynolds, P.; Kristensen, T. Freshwater treatment of amoebic gill disease and sea-lice in seawater salmon production: Considerations of water chemistry and fish welfare in Norway. Aquaculture 2015, 448, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.B.; Crosbie, P.B.B.; Nowak, B.F. Preliminary success using hydrogen peroxide to treat Atlantic salmon, Salmo salar L., affected with experimentally induced amoebic gill disease (AGD). J. Fish Dis. 2012, 35, 839–848. [Google Scholar] [CrossRef]
- Adams, M.B.; Gross, K.A.; Nowak, B.F. Branchial mechanical injury does not accelerate the progression of experimentally induced amoebic gill disease (AGD) in Atlantic salmon Salmo salar L. Aquaculture 2009, 290, 28–36. [Google Scholar] [CrossRef]
- Yavuzcan Yildiz, H.; Meric, İ.; Mehmet, G.; Ergonul, B. Changes of non-specific immune parameters in rainbow trout, Oncorhynchus mykiss, after exposure to antimicrobial agents used in aquaculture. J. Appl. Aquac. 2009, 21, 139–150. [Google Scholar] [CrossRef]
- Soleng, M.; Johansen, L.H.; Johnsen, H.; Johansson, G.S.; Breiland, M.W.; Rørmark, L.; Pittman, K.; Pedersen, L.F.; Lazado, C.C. Atlantic salmon (Salmo salar) mounts systemic and mucosal stress responses to peracetic acid. Fish Shellfish Immunol. 2019, 93, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Vera, L.M.; Migaud, H. Hydrogen peroxide treatment in Atlantic salmon induces stress and detoxification response in a daily manner. Chronobiol. Int. 2016, 33, 530–542. [Google Scholar] [CrossRef] [Green Version]
- Genaro Sanchez, J.; Speare, D.J.; MacNair, N.; Johnson, G. Effects of a prophylactic chloramine-T treatment on growth performance and condition indices of rainbow trout. J. Aquat. Anim. Health 1996, 8, 278–284. [Google Scholar] [CrossRef]
- Downes, J.; Henshilwood, K.; Collins, E.; Ryan, A.; O’Connor, I.; Rodger, H.; MacCarthy, E.; Ruane, N. A longitudinal study of amoebic gill disease on a marine Atlantic salmon farm utilising a real-time PCR assay for the detection of Neoparamoeba perurans. Aquac. Environ. Interact. 2015, 7, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.D.; Harris, J.O.; Carson, J.; Hill, J.V. Effects of gill abrasion and experimental infection with Tenacibaculum maritimum on the respiratory physiology of Atlantic salmon Salmo salar affected by amoebic gill disease. Dis. Aquat. Organ. 2005, 63, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.H.; Pridgeon, J.W.; Klesius, P.H.; Shoemaker, C.A. Parasitism by protozoan Ichthyophthirius multifiliis enhanced invasion of Aeromonas hydrophila in tissues of channel catfish. Vet. Parasitol. 2012, 184, 101–107. [Google Scholar] [CrossRef]
- Liu, Y.J.; Lu, C.P. Role of Ichthyophthirius multifiliis in the Infection of Aeromonas hydrophila. J. Vet. Med. Ser. B 2004, 51, 222–224. [Google Scholar] [CrossRef]
- Bandilla, M.; Valtonen, E.T.; Suomalainen, L.R.; Aphalo, P.J.; Hakalahti, T. A link between ectoparasite infection and susceptibility to bacterial disease in rainbow trout. Int. J. Parasitol. 2006, 36, 987–991. [Google Scholar] [CrossRef]
- Mohammed, H.H.; Arias, C.R. Potassium permanganate elicits a shift of the external fish microbiome and increases host susceptibility to columnaris disease. Vet. Res. 2015, 46, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, V.; Amaral-Zettler, L.; Davidson, J.; Summerfelt, S.; Good, C. Influence of fishmeal-free diets on microbial communities in atlantic salmon (Salmo Salar) recirculation aquaculture systems. Appl. Environ. Microbiol. 2016, 82, 4470–4481. [Google Scholar] [CrossRef] [Green Version]
- Reid, K.M.; Patel, S.; Robinson, A.J.; Bu, L.; Jarungsriapisit, J.; Moore, L.J.; Salinas, I. Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.) post-smolts. PLoS ONE 2017, 12, e0172856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostland, V.; Ferguson, H.; Prescott, J.; Stevenson, R.; Barker, I. Bacterial gill disease of salmonids; relationship between the severity of gill lesions and bacterial recovery. Dis. Aquat. Organ. 1990, 9, 5–14. [Google Scholar] [CrossRef]
- Navarrete, P.; Mardones, P.; Opazo, R.; Espejo, R.; Romero, J. Oxytetracycline treatment reduces bacterial diversity of intestinal microbiota of atlantic salmon. J. Aquat. Anim. Health 2008, 20, 177–183. [Google Scholar] [CrossRef]
- Rosado, D.; Xavier, R.; Severino, R.; Tavares, F.; Cable, J.; Pérez-Losada, M. Effects of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed seabass (Dicentrarchus labrax). Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A.; Catalano, S.R.; Wos-Oxley, M.L.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. Antibiotic-induced alterations and repopulation dynamics of yellowtail kingfish microbiota. Anim. Microbiome 2020, 2, 1–16. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A.; Catalano, S.R.; Wos-Oxley, M.L.; Stephens, F.; Landos, M.; Bansemer, M.S.; Stone, D.A.J.; Qin, J.G.; Oxley, A.P.A. The inner workings of the outer surface: Skin and gill microbiota as indicators of changing gut health in Yellowtail Kingfish. Front. Microbiol. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Slinger, J.; Adams, M.B.; Wynne, J.W. Comparison of bacterial diversity and distribution on the gills of Atlantic salmon (Salmo salar L.): An evaluation of sampling techniques. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zilberg, D.; Munday, B.L. Pathology of experimental amoebic gill disease in Atlantic salmon, Salmo salar L., and the effect of pre-maintenance of fish in sea water on the infection. J. Fish Dis. 2000, 23, 401–407. [Google Scholar] [CrossRef]
- Roberts, S.D.; Powell, M.D. Oral L-cysteine ethyl ester (LCEE) reduces amoebic gill disease (AGD) in Atlantic salmon Salmo salar. Dis. Aquat. Organ. 2005, 66, 21–28. [Google Scholar] [CrossRef]
- Morrison, R.N.; Crosbie, P.B.B.; Nowak, B.F. The induction of laboratory-based amoebic gill disease revisited. J. Fish Dis. 2004, 27, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Pennacchi, Y.; Leef, M.J.; Crosbie, P.B.B.; Nowak, B.F.; Bridle, A.R. Evidence of immune and inflammatory processes in the gills of AGD-affected Atlantic salmon, Salmo salar L. Fish Shellfish Immunol. 2014, 36, 563–570. [Google Scholar] [CrossRef]
- Wynne, J.W.; Stratford, C.; Slinger, J.; Samsing, F.; Rigby, M.; McCulloch, R.; Quezada-Rodriguez, P.; Taylor, R.S. The interaction between temperature and dose on the efficacy and biochemical response of Atlantic salmon to hydrogen peroxide treatment for amoebic gill disease. J. Fish Dis. 2020, 43, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.S.; Slinger, J.; Camargo Lima, P.; English, C.J.; Maynard, B.T.; Samsing, F.; McCulloch, R.; Quezada-Rodriguez, P.R.; Wynne, J.W. Evaluation of sodium percarbonate as a bath treatment for amoebic gill disease in Atlantic salmon. Aquac. Res. 2020, 52, 117–129. [Google Scholar] [CrossRef]
- Morrison, R.N.; Crosbie, P.B.B.; Cook, M.T.; Adams, M.B.; Nowak, B.F. Cultured gill-derived Neoparamoeba pemaquidensis fails to elicit amoebic gill disease (AGD) in Atlantic salmon Salmo salar. Dis. Aquat. Organ. 2005, 66, 135–144. [Google Scholar] [CrossRef]
- Vincent, B.N.; Adams, M.B.; Crosbie, P.B.B.; Nowak, B.F.; Morrison, R.N. Atlantic salmon (Salmo salar L.) exposed to cultured gill-derived Neoparamoeba branchiphila fail to develop amoebic gill disease (AGD). Bull. Eur. Assoc. Fish Pathol. 2007, 27, 112–115. [Google Scholar]
- Kent, M.L.; Sawyer, T.K.; Hedrick, R.P. Paramoeba pemaquidensis (Sarcomastigophora: Paramoebidae) infestation of the gills of coho salmon Oncorhynchus kisutch reared in sea water. Dis. Aquat. Organ. 1988, 5, 163–169. [Google Scholar] [CrossRef]
- Karlsen, C.; Ottem, K.F.; Brevik, Ø.J.; Davey, M.; Sørum, H.; Winther-Larsen, H.C. The environmental and host-associated bacterial microbiota of Arctic seawater-farmed Atlantic salmon with ulcerative disorders. J. Fish Dis. 2017, 40, 1645–1663. [Google Scholar] [CrossRef]
- McIlroy, S.J.; Nielsen, P.H. The family Saprospiraceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 863–889. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slinger, J.; Adams, M.B.; Stratford, C.N.; Rigby, M.; Wynne, J.W. The Effect of Antimicrobial Treatment upon the Gill Bacteriome of Atlantic Salmon (Salmo salar L.) and Progression of Amoebic Gill Disease (AGD) In Vivo. Microorganisms 2021, 9, 987. https://doi.org/10.3390/microorganisms9050987
Slinger J, Adams MB, Stratford CN, Rigby M, Wynne JW. The Effect of Antimicrobial Treatment upon the Gill Bacteriome of Atlantic Salmon (Salmo salar L.) and Progression of Amoebic Gill Disease (AGD) In Vivo. Microorganisms. 2021; 9(5):987. https://doi.org/10.3390/microorganisms9050987
Chicago/Turabian StyleSlinger, Joel, Mark B. Adams, Chris N. Stratford, Megan Rigby, and James W. Wynne. 2021. "The Effect of Antimicrobial Treatment upon the Gill Bacteriome of Atlantic Salmon (Salmo salar L.) and Progression of Amoebic Gill Disease (AGD) In Vivo" Microorganisms 9, no. 5: 987. https://doi.org/10.3390/microorganisms9050987
APA StyleSlinger, J., Adams, M. B., Stratford, C. N., Rigby, M., & Wynne, J. W. (2021). The Effect of Antimicrobial Treatment upon the Gill Bacteriome of Atlantic Salmon (Salmo salar L.) and Progression of Amoebic Gill Disease (AGD) In Vivo. Microorganisms, 9(5), 987. https://doi.org/10.3390/microorganisms9050987