
Persistence of Coxsackievirus B4 in Pancreatic β Cells Disturbs Insulin Maturation, Pattern of Cellular Proteins, and DNA Methylation

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Virus
2.2. Chronic Cell Infections and Viral Progeny in Supernatants
2.3. Cell Viability Assay
2.4. DNA and RNA Extraction
2.5. One-Step RT-PCR and Agarose Gel Electrophoresis
2.6. Real-Time Quantitative PCR
2.7. Quantification of Insulin, Proinsulin, and C-Peptide
2.8. Immunofluorescence Assay
2.9. Quantification of DNA Methylation
2.10. Protein Extraction and Digestion
2.11. Mass Spectrometry Data Acquisition UPLC-MS/MS
2.12. Data and Statistical Analyses
3. Results
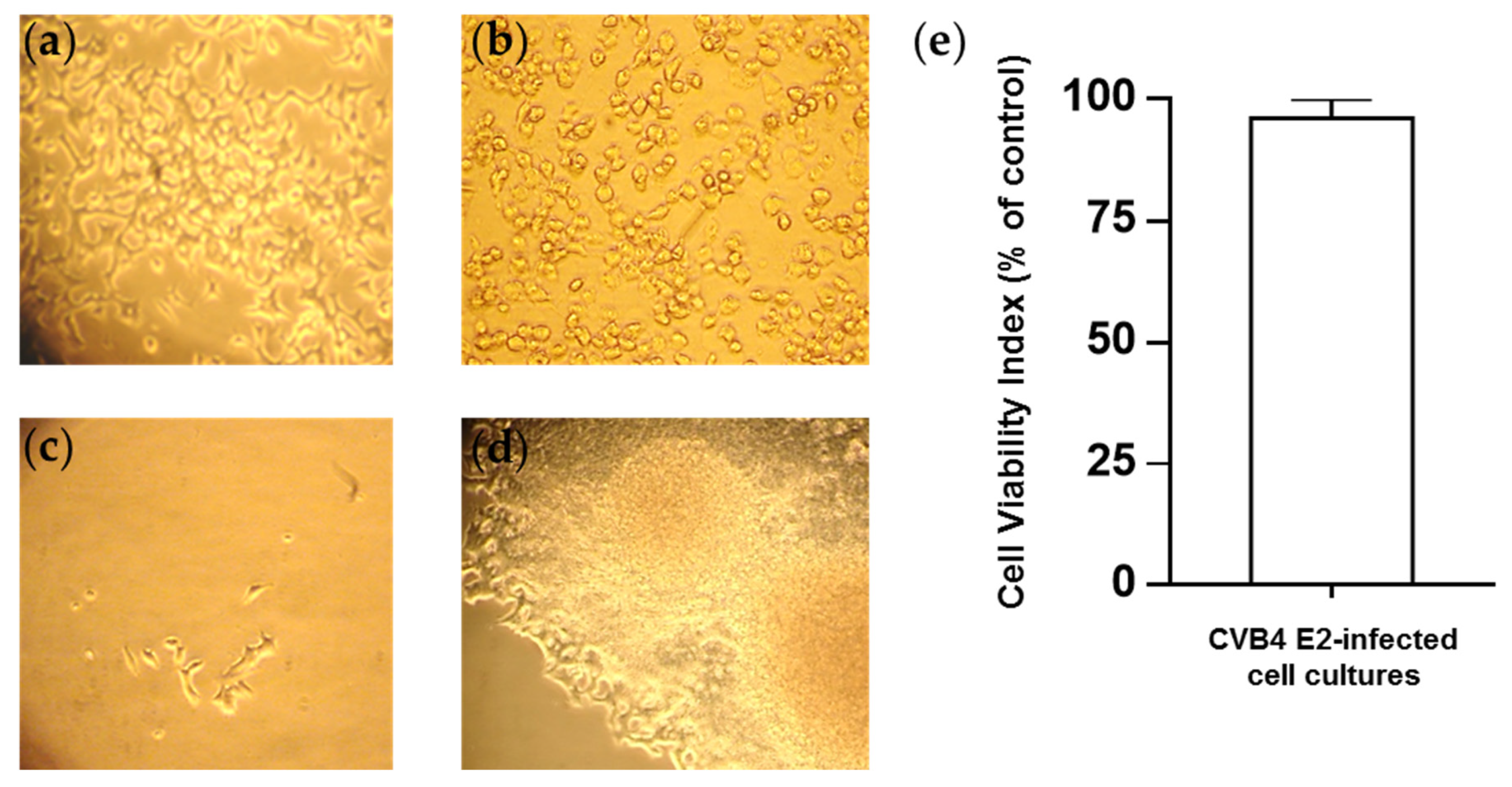
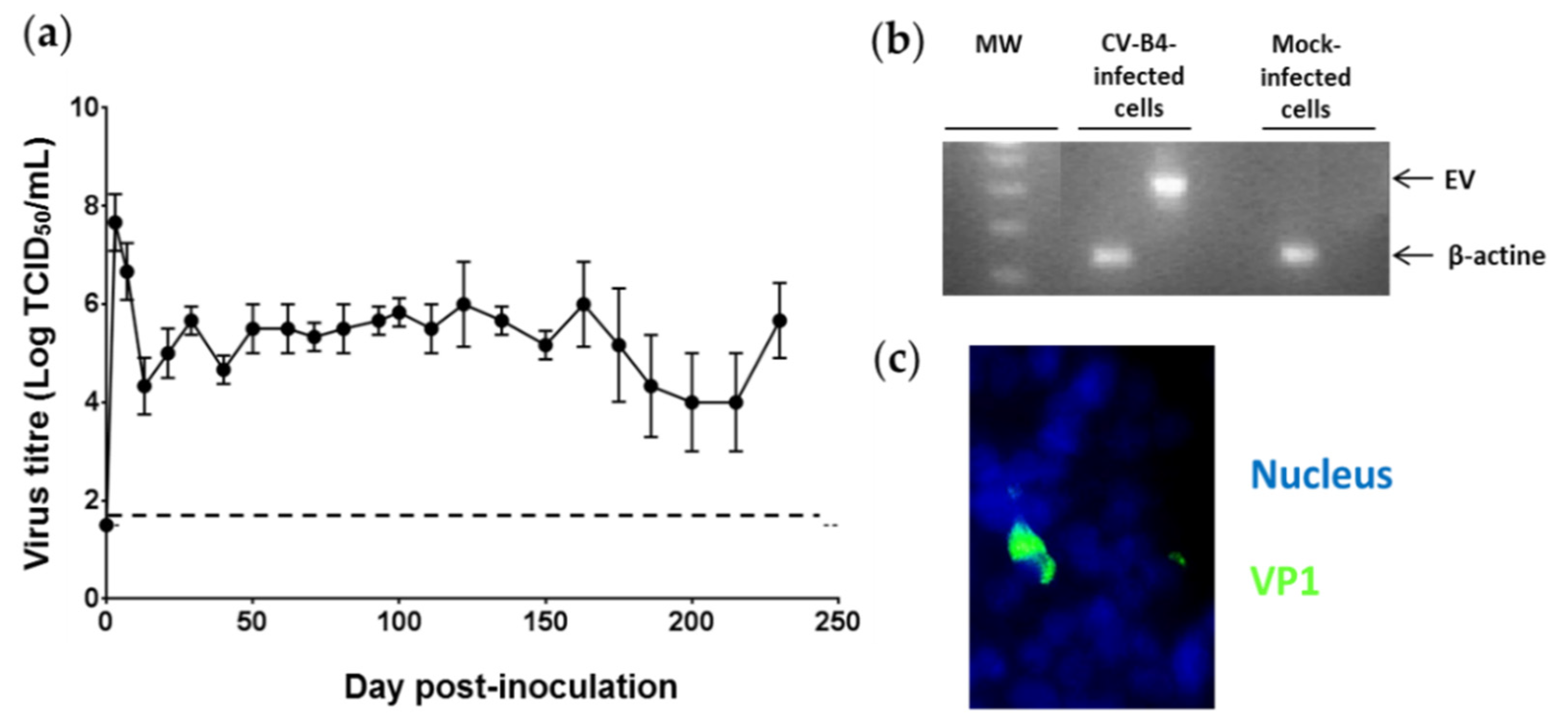
3.1. Persistent CV-B4 E2 Infection of Pancreatic β Cells
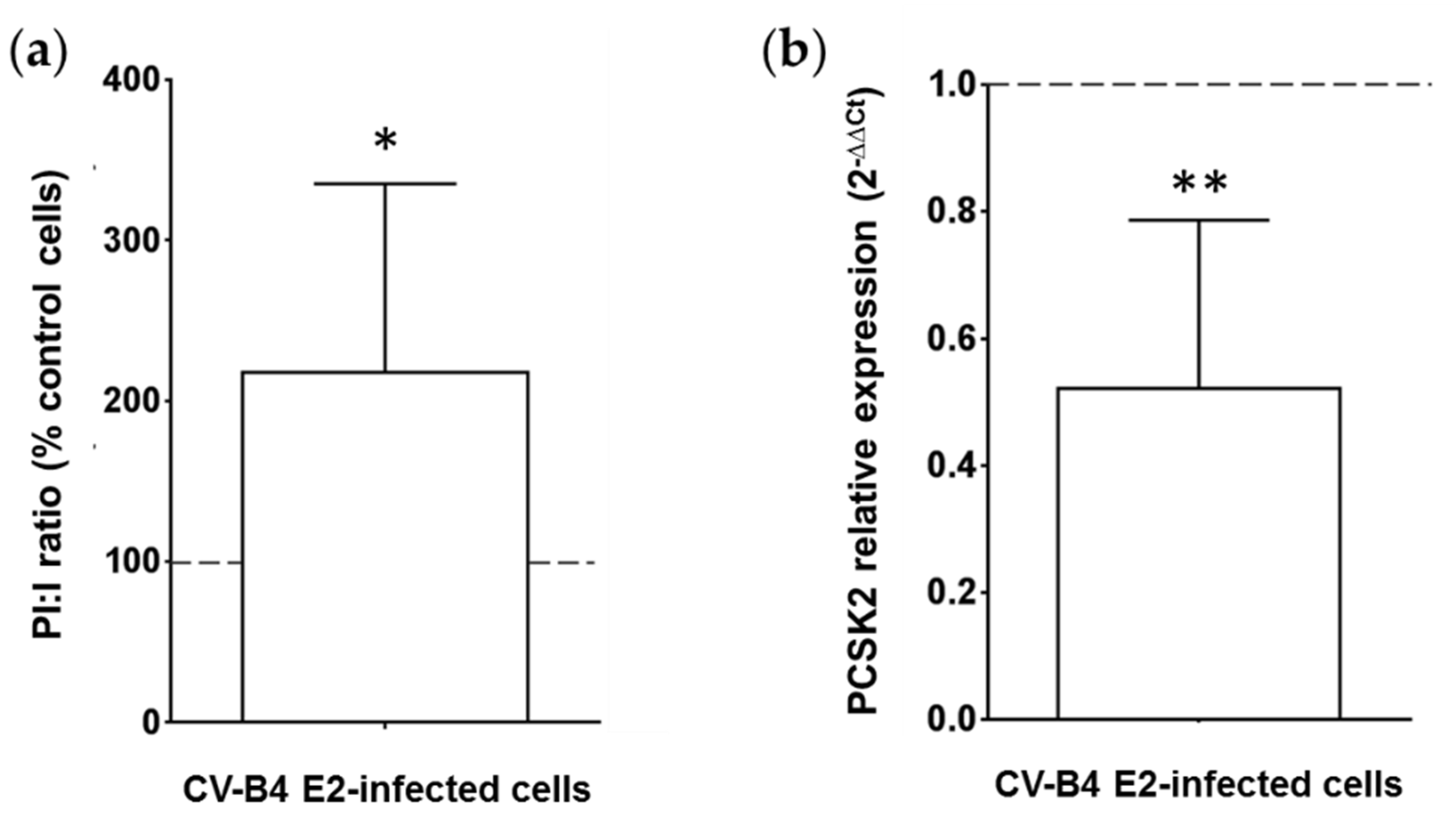
3.2. Persistent CV-B4 E2 Infection of Pancreatic β Cells Results in Change in Insulin Metabolism
3.3. Persistent CV-B4 E2 Infection of Pancreatic β Cells Changes the Expression of Cellular Proteins
3.4. Persistent CV-B4 E2 Infection of Pancreatic β Cells Causes Changes in DNA Methylation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018, 217, 2273–2289. [Google Scholar] [CrossRef] [Green Version]
- Smeekens, S.P.; Montag, A.G.; Thomas, G.; Albiges-Rizo, C.; Carroll, R.; Benig, M.; Phillips, L.A.; Martin, S.; Ohagi, S.; Gardner, P.; et al. Proinsulin processing by the subtilisin-related proprotein convertases furin, PC2, and PC3. Proc. Natl. Acad. Sci. USA 1992, 89, 8822–8826. [Google Scholar] [CrossRef] [Green Version]
- Davidson, H.W.; Rhodes, C.J.; Hutton, J.C. Intraorganellar calcium and pH control proinsulin cleavage in the pancreatic β cell via two distinct site-specific endopeptidases. Nature 1988, 333, 93–96. [Google Scholar] [CrossRef]
- Wasserfall, C.; Nick, H.S.; Campbell-Thompson, M.; Beachy, D.; Haataja, L.; Kusmartseva, I.; Posgai, A.; Beery, M.; Rhodes, C.; Bonifacio, E.; et al. Persistence of pancreatic insulin mRNA expression and proinsulin protein in type 1 diabetes pancreata. Cell Metab. 2017, 26, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Weiss, M.A.; Arunagiri, A.; Yong, J.; Rege, N.; Sun, J.; Haataja, L.; Kaufman, R.J.; Arvan, P. Biosynthesis, structure, and folding of the insulin precursor protein. Diabetes Obes. Metab. 2018, 20, 28–50. [Google Scholar] [CrossRef]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef]
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV virus taxonomy profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef]
- Romero, J.R. Pediatric group B coxsackievirus infections. In Group B Coxsackieviruses; Springer: Berlin/Heidelberg, Germany, 2008; Volume 323, pp. 223–239. [Google Scholar]
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Alidjinou, E.K. Enteroviral pathogenesis of type 1 diabetes: Queries and answers. Curr. Opin. Infect. Dis. 2013, 26, 263–269. [Google Scholar] [CrossRef]
- Ylipaasto, P.; Klingel, K.; Lindberg, A.M.; Otonkoski, T.; Kandolf, R.; Hovi, T.; Roivainen, M. Enterovirus infection in human pancreatic islet cells, islet tropism in vivo and receptor involvement in cultured islet beta cells. Diabetologia 2004, 47, 225–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotta, F.; Censini, S.; Van Halteren, A.G.S.; Marselli, L.; Masini, M.; Dionisi, S.; Mosca, F.; Boggi, U.; Muda, A.O.; Del Prato, S.; et al. Coxsackie B4 virus infection of β cells and natural killer cell insulitis in recent-onset type 1 diabetic patients. Proc. Natl. Acad. Sci. USA 2007, 104, 5115–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.J.; Willcox, A.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. The prevalence of enteroviral capsid protein vp1 immunostaining in pancreatic islets in human type 1 diabetes. Diabetologia 2009, 52, 1143–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.J.; Leete, P.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Expression of the enteroviral capsid protein VP1 in the islet cells of patients with type 1 diabetes is associated with induction of protein kinase R and downregulation of Mcl-1. Diabetologia 2013, 56, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willcox, A.; Richardson, S.J.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Immunohistochemical analysis of the relationship between islet cell proliferation and the production of the enteroviral capsid protein, VP1, in the islets of patients with recent-onset type 1 diabetes. Diabetologia 2011, 54, 2417–2420. [Google Scholar] [CrossRef]
- Yeung, W.-C.G.; Rawlinson, W.D.; Craig, M.E. Enterovirus infection and type 1 diabetes mellitus: Systematic review and meta-analysis of observational molecular studies. BMJ 2011, 342, d35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshebani, A.; Olsson, A.; Westman, J.; Tuvemo, T.; Korsgren, O.; Frisk, G. Effects on isolated human pancreatic islet cells after infection with strains of enterovirus isolated at clinical presentation of type 1 diabetes. Virus Res. 2007, 124, 193–203. [Google Scholar] [CrossRef]
- Hodik, M.; Lukinius, A.; Korsgren, O.; Frisk, G. Tropism analysis of two coxsackie B5 strains reveals virus growth in human primary pancreatic islets but not in exocrine cell clusters in vitro. Open Virol. J. 2013, 7, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Anagandula, M.; Richardson, S.J.; Oberste, M.S.; Sioofy-Khojine, A.B.; Hyöty, H.; Morgan, N.G.; Korsgren, O.; Frisk, G. Infection of human islets of langerhans with two strains of coxsackie B virus serotype 1: Assessment of virus replication, degree of cell death and induction of genes involved in the innate immunity pathway. J. Med. Virol. 2014, 86, 1402–1411. [Google Scholar] [CrossRef]
- Alidjinou, E.K.; Sané, F.; Engelmann, I.; Geenen, V.; Hober, D. Enterovirus persistence as a mechanism in the pathogenesis of type 1 diabetes. Discov. Med. 2014, 18, 273–282. [Google Scholar]
- Pinkert, S.; Klingel, K.; Lindig, V.; Dörner, A.; Zeichhardt, H.; Spiller, O.B.; Fechner, H. Virus-host coevolution in a persistently coxsackievirus B3-infected cardiomyocyte cell line. J. Virol. 2011, 85, 13409–13419. [Google Scholar] [CrossRef] [Green Version]
- Schmidtke, M.; Selinka, H.C.; Heim, A.; Jahn, B.; Tonew, M.; Kandolf, R.; Stelzner, A.; Zell, R. Attachment of coxsackievirus B3 variants to various cell lines: Mapping of phenotypic differences to capsid protein VP1. Virology 2000, 275, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alidjinou, E.K.; Engelmann, I.; Bossu, J.; Villenet, C.; Figeac, M.; Romond, M.B.; Sané, F.; Hober, D. Persistence of coxsackievirus B4 in pancreatic ductal-like cells results in cellular and viral changes. Virulence 2017, 8, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Tam, P.E.; Messner, R.P. Molecular mechanisms of coxsackievirus persistence in chronic inflammatory myopathy: Viral RNA persists through formation of a double-stranded complex without associated genomic mutations or evolution. J. Virol. 1999, 73, 10113–10121. [Google Scholar] [CrossRef] [Green Version]
- Chapman, N.M.; Kim, K.-S.; Drescher, K.M.; Oka, K.; Tracy, S. 5’ terminal deletions in the genome of a coxsackievirus B2 strain occurred naturally in human heart. Virology 2008, 375, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Tracy, S.; Smithee, S.; Alhazmi, A.; Chapman, N. Coxsackievirus can persist in murine pancreas by deletion of 5’ terminal genomic sequences. J. Med. Virol. 2015, 87, 240–247. [Google Scholar] [CrossRef]
- Kuo, H.H.; Ahmad, R.; Lee, G.Q.; Gao, C.; Chen, H.R.; Ouyang, Z.; Szucs, M.J.; Kim, D.; Tsibris, A.; Chun, T.W.; et al. Anti-apoptotic protein BIRC5 maintains survival of HIV-1-infected CD4+ T cells. Immunity 2018, 48, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chehadeh, W.; Kerr-Conte, J.; Pattou, F.; Alm, G.; Lefebvre, J.; Wattré, P.; Hober, D. Persistent infection of human pancreatic islets by coxsackievirus B is associated with alpha interferon synthesis in β cells. J. Virol. 2000, 74, 10153–10164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekoua, M.P.; Bertin, A.; Sane, F.; Alidjinou, E.K.; Lobert, D.; Trauet, J.; Hober, C.; Engelmann, I.; Moutairou, K.; Yessoufou, A.; et al. Pancreatic beta cells persistently infected with coxsackievirus B4 are targets of NK cell-mediated cytolytic activity. Cell. Mol. Life Sci. 2020, 77, 179–194. [Google Scholar] [CrossRef]
- De Beeck, A.O.; Eizirik, D.L. Viral infections in type 1 diabetes mellitus—Why the β cells? Nat. Rev. Endocrinol. 2016, 12, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Lincez, P.J.; Shanina, I.; Horwitz, M.S. Reduced expression of the MDA5 Gene IFIH1 prevents autoimmune diabetes. Diabetes 2015, 64, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, I.; Alidjinou, E.K.; Bertin, A.; Bossu, J.; Villenet, C.; Figeac, M.; Sane, F.; Hober, D. Persistent coxsackievirus B4 infection induces microRNA dysregulation in human pancreatic cells. Cell. Mol. Life Sci. 2017, 74, 3851–3861. [Google Scholar] [CrossRef]
- Sane, F.; Caloone, D.; Gmyr, V.; Engelmann, I.; Belaich, S.; Kerr-Conte, J.; Pattou, F.; Desailloud, R.; Hober, D. Coxsackievirus B4 can infect human pancreas ductal cells and persist in ductal-like cell cultures which results in inhibition of Pdx1 expression and disturbed formation of islet-like cell aggregates. Cell. Mol. Life Sci. 2013, 70, 4169–4180. [Google Scholar] [CrossRef] [PubMed]
- Chick, W.L.; Warren, S.; Chute, R.N.; Like, A.A.; Lauris, V.; Kitchen, K.C. A transplantable insulinoma in the rat. Proc. Natl. Acad. Sci. USA 1977, 74, 628–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.; Halban, P.A.; Wollheim, C.B. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Dechaumes, A.; Nekoua, M.P.; Belouzard, S.; Sane, F.; Engelmann, I.; Dubuisson, J.; Alidjinou, E.K.; Hober, D. Fluoxetine can inhibit SARS-CoV-2 in vitro. Microorganisms 2021, 9, 339. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bertin, A.; Sane, F.; Gmyr, V.; Lobert, D.; Dechaumes, A.; Kerr-Conte, J.; Pattou, F.; Hober, D. Coxsackievirus-b4 infection of human primary pancreatic ductal cell cultures results in impairment of differentiation into insulin-producing cells. Viruses 2019, 11, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, L.; Medina, A.; Aziz, K.; Anagandula, M.; Cabrera-Rode, E.; Fex, M.; Frisk, G.; Cilio, C.M. Differential effects of three echovirus strains on cell lysis and insulin secretion in beta cell derived lines. J. Med. Virol. 2016, 88, 971–978. [Google Scholar] [CrossRef]
- Yin, H.; Berg, A.-K.; Westman, J.; Hellerström, C.; Frisk, G. Complete nucleotide sequence of a Coxsackievirus B-4 strain capable of establishing persistent infection in human pancreatic islet cells: Effects on insulin release, proinsulin synthesis, and cell morphology. J. Med. Virol. 2002, 68, 544–557. [Google Scholar] [CrossRef]
- Lombardi, A.; Tomer, Y. Interferon alpha impairs insulin production in human beta cells via endoplasmic reticulum stress. J. Autoimmun. 2017, 80, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Denning, S.M.; Kurtzberg, J.; Le, P.T.; Tuck, D.T.; Singer, K.H.; Haynes, B.F. Human thymic epithelial cells directly induce activation of autologous immature thymocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 3125–3129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Leung, K.C.; Rawlinson, W.D.; Naing, Z.; Craig, M.E. Enterovirus infection induces cytokine and chemokine expression in insulin-producing cells. J. Med. Virol. 2010, 82, 1950–1957. [Google Scholar] [CrossRef] [PubMed]
- Goffard, A.; Alidjinou, E.K.; Sané, F.; Choteau, L.; Bouquillon, C.; Caloone, D.; Lobert, P.E.; Hober, D. Antibodies enhance the infection of phorbol-ester-differentiated human monocyte-like cells with coxsackievirus B4. Microbes Infect. 2013, 15, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Steenkamp, D.W.; Cacicedo, J.M.; Sahin-Efe, A.; Sullivan, C.; Sternthal, E. Preserved proinsulin secretion in long-standing type 1 diabetes. Endocr. Pract. 2017, 23, 1387–1393. [Google Scholar] [CrossRef]
- Cianciaruso, C.; Phelps, E.A.; Pasquier, M.; Hamelin, R.; Demurtas, D.; Ahmed, M.A.; Piemonti, L.; Hirosue, S.; Swartz, M.A.; De Palma, M.; et al. Primary human and rat β-Cells release the intracellular autoantigens GAD65, IA-2, and proinsulin in exosomes together with cytokine-induced enhancers of immunity. Diabetes 2017, 66, 460–473. [Google Scholar] [CrossRef] [Green Version]
- So, M.; Elso, C.M.; Tresoldi, E.; Pakusch, M.; Pathiraj, V.; Wentworth, J.M.; Harrison, L.C.; Krishnamurthy, B.; Thomas, H.E.; Rodda, C.; et al. Proinsulin C-peptide is an autoantigen in people with type 1 diabetes. Proc. Natl. Acad. Sci. USA 2018, 115, 10732–10737. [Google Scholar] [CrossRef] [Green Version]
- Michels, A.W.; Landry, L.G.; McDaniel, K.A.; Yu, L.; Campbell-Thompson, M.; Kwok, W.W.; Jones, K.L.; Gottlieb, P.A.; Kappler, J.W.; Tang, Q.; et al. Islet-derived CD4 T cells targeting proinsulin in human autoimmune diabetes. Diabetes 2017, 66, 722–734. [Google Scholar] [CrossRef] [Green Version]
- Leong, W.F.; Chow, V.T.K. Transcriptomic and proteomic analyses of rhabdomyosarcoma cells reveal differential cellular gene expression in response to enterovirus 71 infection. Cell. Microbiol. 2006, 8, 565–580. [Google Scholar] [CrossRef]
- Rassmann, A.; Martin, U.; Saluz, H.P.; Peter, S.; Munder, T.; Henke, A. Identification of gene expression profiles in HeLa cells and HepG2 cells infected with Coxsackievirus B3. J. Virol. Methods 2013, 187, 190–194. [Google Scholar] [CrossRef]
- Grubman, M.J.; Moraes, M.P.; Diaz-San Segundo, F.; Pena, L.; De Los Santos, T. Evading the host immune response: How foot-and-mouth disease virus has become an effective pathogen. FEMS Immunol. Med. Microbiol. 2008, 53, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Yu, B.; Lin, L.; Zhai, X.; Han, Y.; Qin, Y.; Guo, Z.; Wu, S.; Zhong, X.; Wang, Y.; et al. A functional nuclear localization sequence in the VP1 capsid protein of coxsackievirus B3. Virology 2012, 433, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Falk, M.M.; Grigera, P.R.; Bergmann, I.E.; Zibert, A.; Multhaup, G.; Beck, E. Foot-and-mouth disease virus protease 3C induces specific proteolytic cleavage of host cell histone H3. J. Virol. 1990, 64, 748–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griger, P.R.; Tisminetzky, S.G. Histone H3 modification in BHK cells infected with foot-and-mouth disease virus. Virology 1984, 136, 10–19. [Google Scholar] [CrossRef]
- Nguyen, E.V.; Gharib, S.A.; Crothers, K.; Chow, Y.H.; Park, D.R.; Goodlett, D.R.; Schnapp, L.M. Proteomic landscape of bronchoalveolar lavage fluid in human immunodeficiency virus infection. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306. [Google Scholar] [CrossRef] [Green Version]
- Hammer, E.; Goritzka, M.; Ameling, S.; Darm, K.; Steil, L.; Klingel, K.; Trimpert, C.; Herda, L.R.; Dörr, M.; Kroemer, H.K.; et al. Characterization of the human myocardial proteome in inflammatory dilated cardiomyopathy by label-free quantitative shotgun proteomics of heart biopsies. J. Proteome Res. 2011, 10, 2161–2171. [Google Scholar] [CrossRef]
- Hsu, Y.Y.; Liu, Y.N.; Lu, W.W.; Kung, S.H. Visualizing and quantifying the differential cleavages of the eukaryotic translation initiation factors eIF4GI and eIF4GII in the enterovirus-infected cell. Biotechnol. Bioeng. 2009, 104, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.S.W.; Moran-Jones, K.; Shan, J.; Munro, T.P.; Snee, M.J.; Hoek, K.S.; Smith, R. Heterogeneous nuclear ribonucleoprotein A3, a novel RNA trafficking response element-binding protein. J. Biol. Chem. 2002, 277, 18010–18020. [Google Scholar] [CrossRef] [Green Version]
- Jagdeo, J.M.; Dufour, A.; Fung, G.; Luo, H.; Kleifeld, O.; Overall, C.M.; Jan, E. Heterogeneous nuclear ribonucleoprotein M facilitates enterovirus infection. J. Virol. 2015, 89, 7064–7078. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, E. ZnT8 and type 1 diabetes. Endocr. J. 2012, 59, 531–537. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence | |
|---|---|---|
| β-actin | Forward | 5′-GGCACTCTTCCAGCCTTCCT-3′ |
| Reverse | 5′-GCAATGCCAGGGTACATGGT-3′ | |
| CV-B4 E2 | Forward | 5′-CAAGCACTTCTGTTTCCCCGG-3′ |
| Reverse | 5′-ATTGTCACCATAAGCAGCCA-3′ | |
| PCSK2 | Forward | 5′-CGAAACCAGCTTCACGATGAG-3′ |
| Reverse | 5′-ACGCCGGCTTAGCAAAATGGA-3′ | |
| Accession | Protein | Infected Cells Score | Controls Score | Score Difference | p-Value |
|---|---|---|---|---|---|
| P21807 | Peripherin | 0.00 ± 0.00 | 12.56 ± 4.10 | −12.56 | *** |
| Q07266-2 | Isoform E1 of Drebrin | 6.29 ± 3.00 | 17.56 ± 2.90 | −11.27 | *** |
| Q4QQS7 | Protein Umps | 0.49 ± 0.99 | 5.40 ± 1.14 | −4.91 | *** |
| P34926 | Microtubule-associated protein 1A | 2.00 ± 1.37 | 14.30 ± 5.17 | −12.3 | ** |
| Q5XIM9 | T-complex protein 1 subunit beta | 7.86 ± 3.07 | 15.90 ± 2.48 | −8.04 | ** |
| P23514 | Coatomer subunit beta | 1.05 ± 2.10 | 5.01 ± 1.13 | −3.96 | ** |
| D4A2G9 | Protein Ranbp1 | 0.00 ± 0.00 | 3.60 ± 1.79 | −3.6 | ** |
| G3V7N5 | Carnitine O-palmitoyltransferase 2, mitochondrial | 0.00 ± 0.00 | 3.38 ± 1.66 | −3.38 | ** |
| D3ZMY7 | Protein Nt5c2 | 0.43 ± 0.87 | 3.75 ± 1.12 | −3.32 | ** |
| F1LMC7 | Septin-7 | 0.44 ± 0.87 | 2.60 ± 1.37 | −2.16 | ** |
| A0A0G2K1J5 | Plectin | 4.04 ± 2.38 | 21.11 ± 13.47 | −17.07 | * |
| Q6URK4 | Heterogeneous nuclear ribonucleoprotein A3 (HNRPA3) | 0.00 ± 0.00 | 16.28 ± 13.66 | −16.28 | * |
| P12785 | Fatty acid synthase | 6.78 ± 3.89 | 22.06 ± 8.44 | −15.28 | * |
| A0A0G2JU82 | Microtubule-actin cross-linking factor 1 | 7.59 ± 5.23 | 22.14 ± 9.29 | −14.55 | * |
| Q05982 | Nucleoside diphosphate kinase A | 6.26 ± 8.23 | 20.65 ± 6.46 | −14.39 | * |
| Q6URK4-2 | Heterogeneous nuclear ribonucleoprotein A3 (HNRPA3) isoform 2 | 18.03 ± 4.53 | 4.98 ± 8.08 | +13.05 | * |
| A0A0G2K013 | Alpha-actinin-4 | 9.04 ± 1.59 | 21.08 ± 7.58 | −12.04 | * |
| P06687 | Sodium/potassium-transporting ATPase subunit alpha-3 | 0.00 ± 0.00 | 11.90 ± 7.78 | −11.9 | * |
| D4AD15 | Protein Eif4g1 | 4.66 ± 2.13 | 16.16 ± 6.89 | −11.5 | * |
| A0A0G2K0Q7 | Protein Mylk | 3.17 ± 2.13 | 11.74 ± 4.47 | −8.57 | * |
| O35314 | Secretogranin-1 | 5.04 ± 1.57 | 13.22 ± 4.74 | −8.18 | * |
| Q62667 | Major vault protein | 6.15 ± 2.75 | 14.06 ± 4.71 | −7.91 | * |
| D4AC23 | Protein Cct7 | 6.14 ± 1.77 | 13.98 ± 4.59 | −7.84 | * |
| P04692-5 | Tropomyosin alpha-1 chain isoform 5 | 1.62 ± 3.23 | 8.57 ± 5.17 | −6.95 | * |
| Q3MIE4 | Synaptic vesicle membrane protein VAT-1 homolog | 2.34 ± 2.20 | 9.00 ± 3.48 | −6.66 | * |
| D3ZRM9 | Uncharacterized protein | 0.00 ± 0.00 | 6.54 ± 3.76 | −6.54 | * |
| D3ZRM9 | Uncharacterized protein | 0.00 ± 0.00 | 6.54 ± 3.76 | −6.54 | * |
| Q62950 | Dihydropyrimidinase-related protein 1 | 2.41 ± 3.70 | 8.67 ± 4.13 | −6.26 | * |
| F1MAA1 | Ubiquitin-specific peptidase 47 | 2.26 ± 2.61 | 8.50 ± 4.06 | −6.24 | * |
| O35303-6 | Dynamin-1-like protein isoform 6 | 1.91 ± 3.83 | 7.53 ± 2.90 | −5.62 | * |
| P41562 | Isocitrate dehydrogenase [NADP] cytoplasmic | 2.57 ± 3.14 | 8.07 ± 2.98 | −5.5 | * |
| D4A0C3 | Protein Hid1 | 1.38 ± 1.78 | 6.44 ± 2.39 | −5.06 | * |
| A0A0G2JZ60 | Protein Fsd1l | 3.86 ± 3.10 | 8.89 ± 2.01 | −5.03 | * |
| A0A0G2JUN7 | Thioredoxin reductase 1, cytoplasmic | 2.77 ± 1.97 | 7.38 ± 2.33 | −4.61 | * |
| D3ZVQ0 | Protein LOC100911959 | 2.72 ± 2.66 | 7.14 ± 1.73 | −4.42 | * |
| O70593 | Small glutamine-rich tetratricopeptide repeat-containing protein alpha | 1.03 ± 1.21 | 5.43 ± 2.75 | −4.4 | * |
| P50475 | Alanine--tRNA ligase, cytoplasmic | 0.44 ± 0.88 | 4.76 ± 2.41 | −4.32 | * |
| P27008 | Poly [ADP-ribose] polymerase 1 | 1.90 ± 2.77 | 6.15 ± 2.62 | −4.25 | * |
| B2RYI2 | Signal recognition particle subunit SRP68 | 2.03 ± 1.60 | 6.04 ± 1.87 | −4.01 | * |
| Q05096-3 | Unconventional myosin-Ib isoform 3 | 1.96 ± 2.70 | 5.89 ± 1.41 | −3.93 | * |
| O88321 | Antisecretory factor | 1.47 ± 1.86 | 5.33 ± 2.30 | −3.86 | * |
| B2GV74 | Kinesin light chain 2 | 1.08 ± 2.16 | 4.89 ± 2.36 | −3.81 | * |
| Q9Z1W6-4 | Protein LYRIC isoform 4 | 0.54 ± 1.08 | 3.58 ± 2.25 | −3.04 | * |
| A0A0H2UHW4 | PEST proteolytic signal-containing nuclear protein | 0.42 ± 0.85 | 3.29 ± 2.02 | −2.87 | * |
| E9PT23 | Putative sodium-coupled neutral amino acid transporter 10 | 0.46 ± 0.93 | 3.00 ± 1.92 | −2.54 | * |
| D3Z8U5 | Metalloendopeptidase | 0.87 ± 1.73 | 3.10 ± 1.25 | −2.23 | * |
| Q5M7W6 | Protein FAM234A | 5.24 ± 0.98 | 1.88 ± 2.18 | +3.36 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekoua, M.P.; Bertin, A.; Sane, F.; Gimeno, J.-P.; Fournier, I.; Salzet, M.; Engelmann, I.; Alidjinou, E.K.; Hober, D. Persistence of Coxsackievirus B4 in Pancreatic β Cells Disturbs Insulin Maturation, Pattern of Cellular Proteins, and DNA Methylation. Microorganisms 2021, 9, 1125. https://doi.org/10.3390/microorganisms9061125
Nekoua MP, Bertin A, Sane F, Gimeno J-P, Fournier I, Salzet M, Engelmann I, Alidjinou EK, Hober D. Persistence of Coxsackievirus B4 in Pancreatic β Cells Disturbs Insulin Maturation, Pattern of Cellular Proteins, and DNA Methylation. Microorganisms. 2021; 9(6):1125. https://doi.org/10.3390/microorganisms9061125
Chicago/Turabian StyleNekoua, Magloire Pandoua, Antoine Bertin, Famara Sane, Jean-Pascal Gimeno, Isabelle Fournier, Michel Salzet, Ilka Engelmann, Enagnon Kazali Alidjinou, and Didier Hober. 2021. "Persistence of Coxsackievirus B4 in Pancreatic β Cells Disturbs Insulin Maturation, Pattern of Cellular Proteins, and DNA Methylation" Microorganisms 9, no. 6: 1125. https://doi.org/10.3390/microorganisms9061125
APA StyleNekoua, M. P., Bertin, A., Sane, F., Gimeno, J.-P., Fournier, I., Salzet, M., Engelmann, I., Alidjinou, E. K., & Hober, D. (2021). Persistence of Coxsackievirus B4 in Pancreatic β Cells Disturbs Insulin Maturation, Pattern of Cellular Proteins, and DNA Methylation. Microorganisms, 9(6), 1125. https://doi.org/10.3390/microorganisms9061125