
Gut Microbiota Dynamics in Natural Populations of Pintomyia evansi under Experimental Infection with Leishmania infantum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Area and Survey of Sand Flies
2.3. Infections under Experimental Conditions of P. evansi with L. infantum
2.4. Sandfly Washing Procedure and Midgut Dissection
2.5. DNA Extraction of Guts from P. evansi
2.6. Bacterial 16S rRNA Gene Fragment PCR Amplification and Sequencing
2.7. Bioinformatics and Statistical Analysis of the Microbiota Data
3. Results
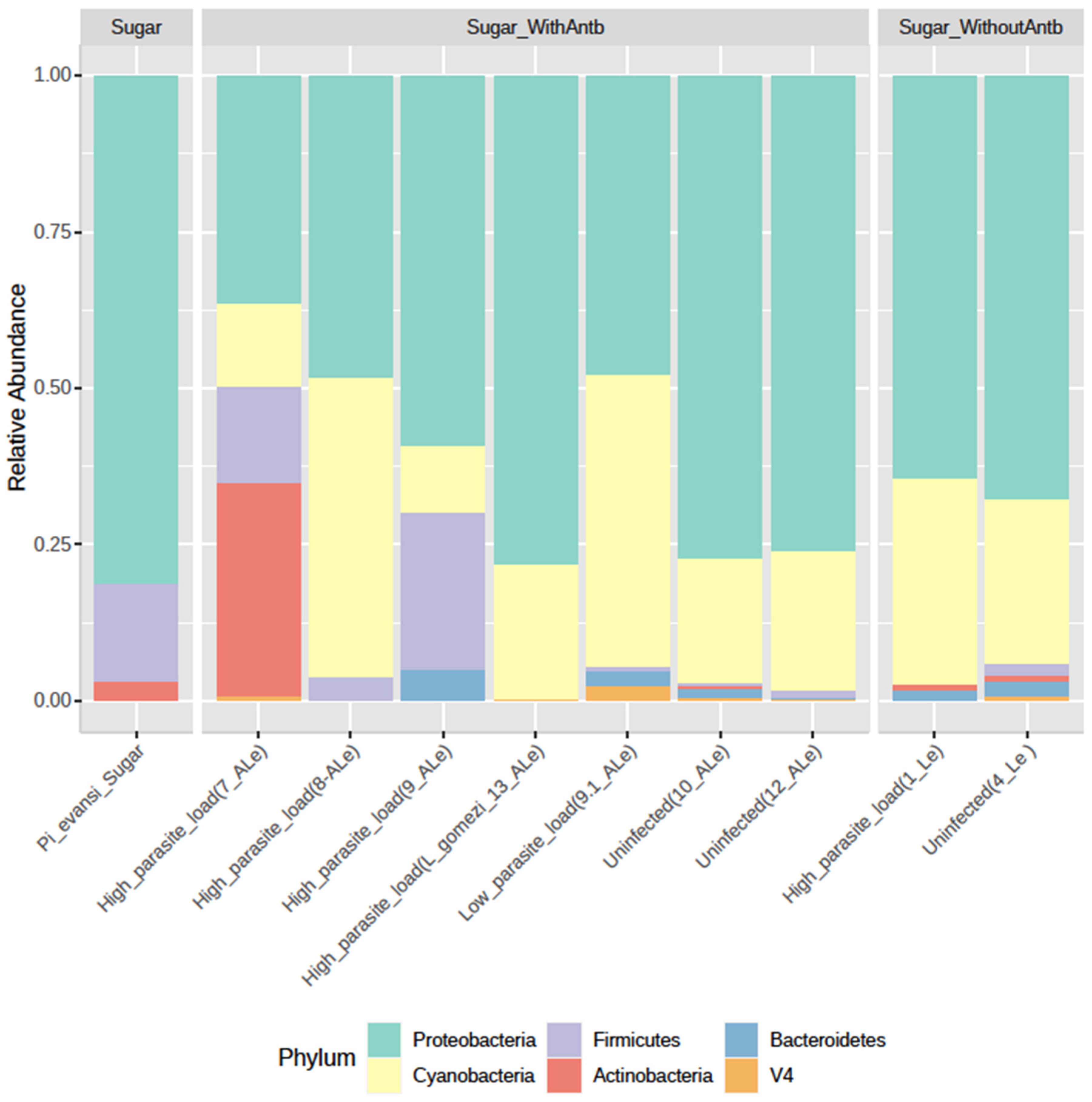
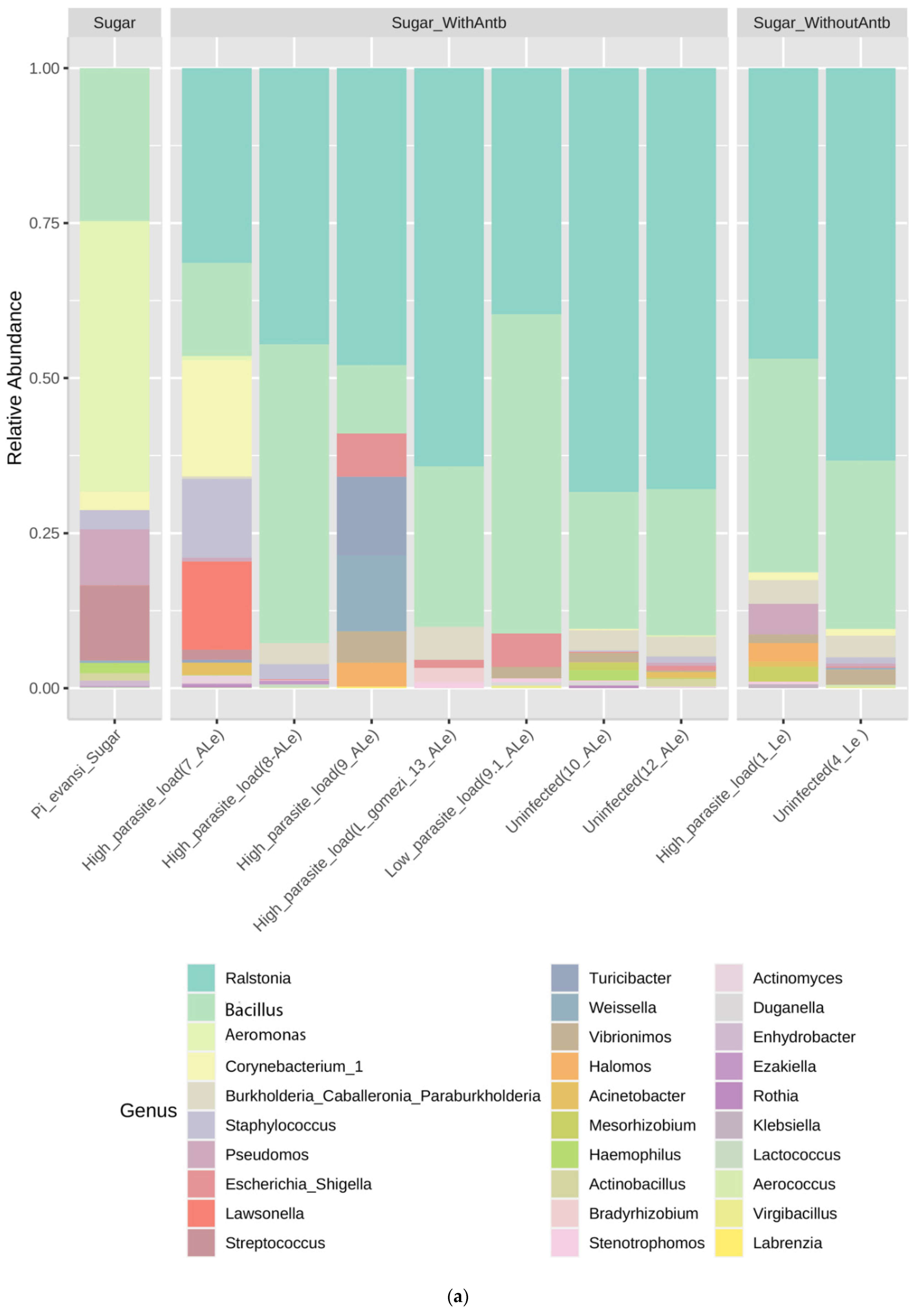
3.1. P. evansi Gut Microbiota Composition
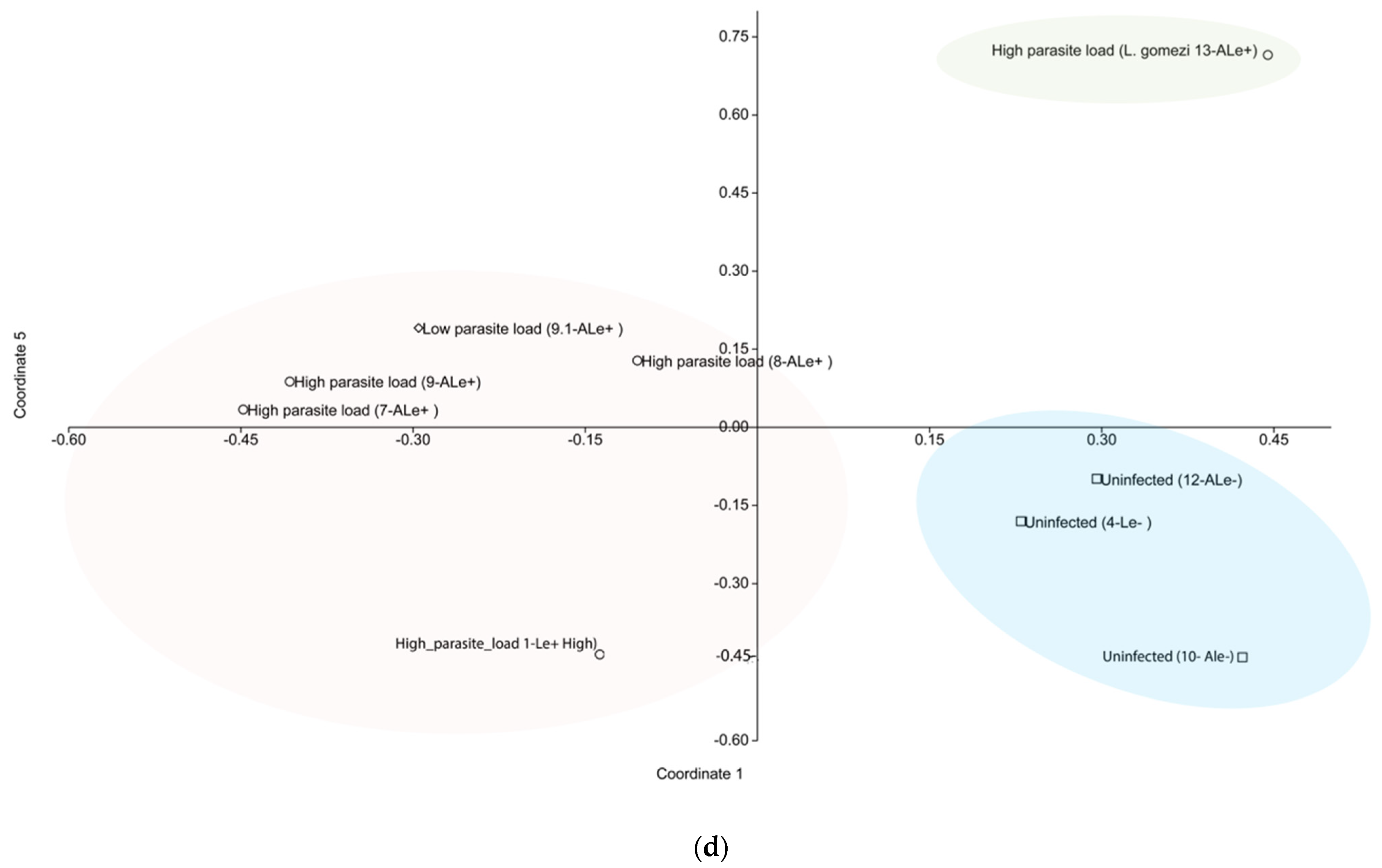
3.2. Influence of Leishmania and Antibiotic Cocktail on Gut Microbiota
3.3. Endosymbiont Detection
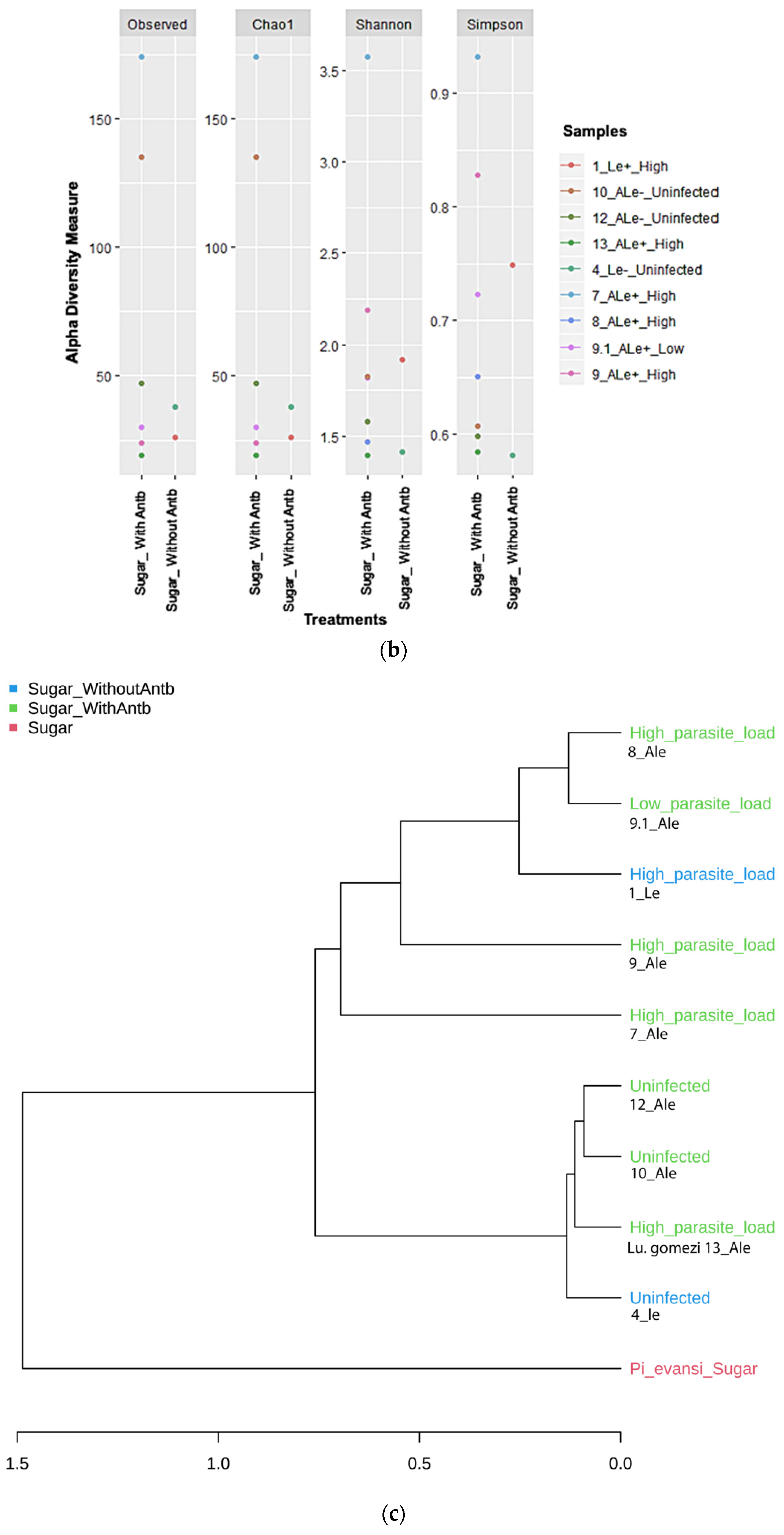
3.4. Diversity of Gut Microbiota in P. evansi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Telleria, E.L.; Martins-Da-Silva, A.; Tempone, A.J.; Traub-Csekö, Y.M. Leishmania, microbiota and sand fly immunity. Parasitology 2018, 145, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omondi, Z.N.; Demir, S. Bacteria composition and diversity in the gut of sand fly: Impact on Leishmania and sand fly development. Int. J. Trop. Insect. Sci. 2021, 41, 25. [Google Scholar] [CrossRef]
- Kelly, P.H.; Bahr, S.M.; Serafim, T.D.; Ajami, N.J.; Petrosino, J.F.; Meneses, C.; Kirby, J.R.; Valenzuela, J.G.; Kamhawi, S.; Wilsone, M.E. The gut microbiome of the vector Lutzomyia longipalpis is essential for survival of Leishmania infantum. mBio 2017, 8, e01121-16. [Google Scholar] [CrossRef] [Green Version]
- Campolina, T.B.; Villegas, L.E.M.; Monteiro, C.C.; Pimenta, P.F.P.; Secundino, N.F.C. Tripartite interactions: Leishmania, microbiota and Lutzomyia longipalpis. PLoS Negl. Trop. Dis. 2020, 14, e0008666. [Google Scholar] [CrossRef]
- Monteiro, C.C.; Villegas, L.E.; Campolina, T.B.; Pires, A.C.; Miranda, J.C.; Pimenta, P.F.; Secundino, N.F. Bacterial diversity of the American sand fly Lutzomyia intermedia using high-throughput metagenomic sequencing. Parasites Vectors 2016, 9, 480. [Google Scholar] [CrossRef] [Green Version]
- Telleria, E.L.; Sant’Anna, M.R.; Alkurbi, M.O.; Pitaluga, A.N.; Dillon, R.J.; Traub-Csekö, Y.M. Bacterial feeding, Leishmania infection and distinct infection routes induce differential defensin expression in Lutzomyia longipalpis. Parasites Vectors 2013, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Telleria, E.L.; de Araújo, A.P.; Secundino, N.F.; d’Avila-Levy, C.M.; Traub-Csekö, Y.M. Trypsin-like serine proteases in Lutzomyia longipalpis—Expression, activity and possible modulation by Leishmania infantum chagasi. PLoS ONE 2010, 5, e10697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sant’Anna, M.R.; Diaz-Albiter, H.; Mubaraki, M.; Dillon, R.J.; Bates, P.A. Inhibition of trypsin expression in Lutzomyia longipalpis using RNAi enhances the survival of Leishmania. Parasites Vectors 2009, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Vivero, R.J.; Cadavid-Restrepo, G.; Herrera, C.X.; Soto, S.I. Molecular detection and identification of Wolbachia in three species of the genus Lutzomyia on the Colombian Caribbean coast. Parasites Vectors 2017, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivero, R.J.; Jaramillo, N.G.; Cadavid-Restrepo, G.; Soto, S.I.; Herrera, C.X. Structural differences in gut bacteria communities in developmental stages of natural populations of Lutzomyia evansi from Colombia’s Caribbean coast. Parasites Vectors 2016, 9, 496. [Google Scholar] [CrossRef] [Green Version]
- Vivero, R.J.; Villegas-Plazas, M.; Cadavid-Restrepo, G.E.; Herrera, C.X.M.; Uribe, S.I.; Junca, H. Wild specimens of sand fly phlebotomine Lutzomyia evansi, vector of leishmaniasis, show high abundance of Methylobacterium and natural carriage of Wolbachia and Cardinium types in the midgut microbiome. Sci. Rep. 2019, 9, 17746. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.B.; Modi, G.B. A simple method for experimental infection of phlebotomine sand flies with Leishmania. Am. J. Trop. Med. Hyg. 1984, 33, 41. [Google Scholar] [CrossRef]
- Santamaría, E.; Ponce, N.; Puerta, C.; Ferro, C. Validación de la PCR en la detección de parásitos de Leishmania (Viannia) spp. en Lutzomyia (Diptera: Psychodidae) como herramienta en la definición de especies vectores. Biomédica 2005, 25, 1349. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, E.; Castillo, M.; Cárdenas, R.; Bello, F.; Ayala, M.; Ferro, C. Competencia vectorial de las especies de Lutzomyia del grupo verrucarum (Diptera, Psychodidae) en un foco endémico de Leishmania braziliensis en Reventones, Cundinamarca. Biomédica 1999, 19, 115. [Google Scholar] [CrossRef] [Green Version]
- Braig, H.R.; Zhou, W.; Dobson, S.L.; O’Neill, S.L. Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis. J. Bacteriol. 1998, 180, 2373. [Google Scholar] [CrossRef] [Green Version]
- Weeks, A.R.; Velten, R.; Stouthamer, R. Incidence of a new sex-ratio-distorting endosymbiotic bacterium among arthropods. Proc. Biol. Sci. 2003, 270, 1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valderrama, A.; Tavares, M.G.; Dilermando Andrade Filho, J. Phylogeography of the Lutzomyia gomezi (Diptera: Phlebotominae) on the Panama Isthmus. Parasites Vectors 2014, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Illumina. 16S Illumina Amplicon Protocol: Earth Microbiome Project; Illumina: San Diego, CA, USA, 2017. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621. [Google Scholar] [CrossRef] [Green Version]
- Rath, S.; Heidrich, B.; Pieper, D.H.; Vital, M. Uncovering the trimethylamine-producing bacteria of the human gut microbiota. Microbiome 2017, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabbabi, A.; Watanabe, S.; Mizushima, D.; Caceres, A.G.; Gomez, E.A.; Yamamoto, D.S.; Cui, L.; Hashiguchi, Y.; Kato, H. (Comparative Analysis of Bacterial Communities in Lutzomyia ayacuchensis Populations with Different Vector Competence to Leishmania Parasites in Ecuador and Peru. Microorganisms 2020, 9, 68. [Google Scholar] [CrossRef]
- Cirimotich, C.M.; Ramirez, J.L.; Dimopoulos, G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 2011, 10, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telleria, E.L.; Sant’Anna, M.R.; Ortigão-Farias, J.R.; Pitaluga, A.N.; Dillon, V.M.; Bates, P.A.; Traub-Cseko, Y.M.; Dillon, R.J. Caspar-like gene depletion reduces leishmania infection in sand fly host Lutzomyia longipalpis. J. Biol. Chem. 2012, 287, 12985. [Google Scholar] [CrossRef] [Green Version]
- Moraes, C.S.; Seabra, S.H.; Castro, D.P.; Brazil, R.P.; de Souza, W.; Garcia, E.S.; Azambuja, P. Leishmania (leishmania) chagasi interactions with Serratia marcescens: Ultrastructural studies, lysis and carbohydrate effects. Exp. Parasitol. 2008, 118, 561. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef] [Green Version]
- Louradour, I.; Monteiro, C.C.; Inbar, E.; Ghosh, K.; Merkhofer, R.; Lawyer, P.; Paun, A.; Smelkinson, M.; Secundino, N.; Lewis, M.; et al. The midgut microbiota plays an essential role in sand fly vector competence for Leishmania major. Cell Microbiol. 2017, 19, 12755. [Google Scholar] [CrossRef] [Green Version]
- Dillon, R.J. A transdisciplinary consideration of sand flies & leishmaniasis. C. R. Biol. 2019, 342, 272. [Google Scholar]
- Sant’Anna, M.R.; Diaz-Albiter, H.; Aguiar-Martins, K.; Al Salem, W.S.; Cavalcante, R.R.; Dillon, V.M.; Bates, P.A.; Genta, F.A.; Dillon, R.J. Colonisation resistance in the sand fly gut: Leishmania protects Lutzomyia longipalpis from bacterial infection. Parasites Vectors 2014, 7, 329. [Google Scholar] [CrossRef] [Green Version]
- Remenant, B.; de Cambiaire, J.C.; Cellier, G.; Jacobs, J.M.; Mangenot, S.; Barbe, V.; Lajus, A.; Vallener, D.; Medigue, C.; Fegan, M.; et al. Ralstonia syzygii, the blood disease bacterium and some Asian R. solanacearum strains form a single genomic species despite divergent lifestyles. PLoS ONE 2011, 6, e24356. [Google Scholar] [CrossRef]
- Yun, J.H.; Roh, S.W.; Whon, T.W.; Jung, M.J.; Kim, M.S.; Park, D.S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montong, V.; Salaki, S. Insects as carriers of Ralstonia solanacearum phylotype IV on kepOK banana flowers in South Minahasa and Minahasa districts. Int. J. ChemTech Res. 2020, 13, 100. [Google Scholar] [CrossRef]
- Sant’Anna, M.R.; Darby, A.C.; Brazil, R.P.; Montoya-Lerma, J.; Dillon, V.M.; Bates, P.A.; Dillon, R.J. Investigation of the bacterial communities associated with females of Lutzomyia sand fly species from South America. PLoS ONE 2012, 7, e42531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, C.B.; Diambra, L.A.; Rivera Pomar, R.V. Metagenomic analysis of taxa associated with Lutzomyia longipalpis, vector of visceral leishmaniasis, using an unbiased high-throughput approach. PLoS Negl. Trop. Dis. 2011, 5, e1304. [Google Scholar] [CrossRef]
- Mukhopadhyay, J.; Braig, H.R.; Rowton, E.D.; Ghosh, K. Naturally occurring culturable aerobic gut flora of adult Phlebotomus papatasi, vector of Leishmania major in the Old World. PLoS ONE 2012, 7, e35748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleki-Ravasan, N.; Oshaghi, M.A.; Hajikhani, S.; Saeidi, Z.; Akhavan, A.A.; Gerami-Shoar, M.; Shirazi, M.H.; Yakhchali, B.; Rassi, Y.; Afshar, D. Aerobic microbial community of insectary population of Phlebotomus papatasi. J. Arthropod. Borne Dis. 2014, 8, 69. [Google Scholar] [PubMed]
- Fraihi, W.; Fares, W.; Perrin, P.; Dorkeld, F.; Sereno, D.; Barhoumi, W.; Sbissi, I.; Cherni, S.; Chelbi, I.; Durvasula, R.; et al. An integrated overview of the midgut bacterial flora composition of Phlebotomus perniciosus, a vector of zoonotic visceral leishmaniasis in the western Mediterranean basin. PLoS Negl. Trop. Dis. 2017, 11, e0005484. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, C.; Asensi, M.D.; Zahner, V.; Rangel, E.F.; Oliveira, S.M. Study on the bacterial midgut microbiota associated to different Brazilian populations of Lutzomyia longipalpis (Lutz & Neiva) (Diptera: Psychodidae). Neotrop. Entomol. 2008, 37, 597. [Google Scholar] [CrossRef] [Green Version]
- Herren, J.K.; Mbaisi, L.; Mararo, E.; Makhulu, E.E.; Mobegi, V.A.; Butungi, H.; Mancini, M.; Oundo, J.W.; Teal, E.T.; Pinaud, S.; et al. A microsporidian impairs Plasmodium falciparum transmission in Anopheles arabiensis mosquitoes. Nat. Commun. 2020, 11, 2187. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The transmission efficiency of tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J. Virol. 2010, 84, 9310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, V.S.; Singh, S.T.; Priya, N.G.; Kumar, J.; Rajagopal, R. Arsenophonus GroEL interacts with CLCuV and is localized in midgut and salivary gland of whitefly B. tabaci. PLoS ONE 2012, 7, e42168. [Google Scholar] [CrossRef] [PubMed]
- Raina, H.S.; Rawal, V.; Singh, S.; Daimei, G.; Shakarad, M.; Rajagopal, R. Elimination of Arsenophonus and decrease in the bacterial symbionts diversity by antibiotic treatment leads to increase in fitness of whitefly, Bemisia tabaci. Infect. Genet. Evol. 2015, 32, 224. [Google Scholar] [CrossRef]
- Nováková, E.; Hypša, V.; Moran, N.A. Arsenophonus, an emerging clade of intracellular symbionts with a broad host distribution. BMC Microbiol. 2009, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, C.; Beeton, M.; Harbison, C.; Jones, T.; Pontes, M. Isolation, pure culture, and characterization of “Candidatus arsenophonus arthropodicus,” an intracellular secondary endosymbiont from the hippoboscid louse fly Pseudolynchia canariensis. Appl. Environ. Microbiol. 2006, 72, 2997. [Google Scholar] [CrossRef] [Green Version]
- Gumiel, M.; da Mota, F.F.; de Sousa Rizzo, V.; Sarquis, O.; de Castro, D.P.; Lima, M.M.; de Souza Garcia, E.; Carels, N.; Azambuja, P. Characterization of the microbiota in the guts of Triatoma brasiliensis and Triatoma pseudomaculata infected by Trypanosoma cruzi in natural conditions using culture independent methods. Parasites Vectors 2015, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Duque, P.; Vélez, I.; Morales, M.; Sierra, D. Sand flies fauna involved in the transmission of cutaneous leishmaniasis in Afro-Colombian and Amerindian communities of Choco, Pacific Coast of Colombia. Neotrop. Entomol. 2004, 33, 255–264. [Google Scholar] [CrossRef]
- Cortés, A.; Pérez-Doria, A.; Bejarano, E. Flebotomíneos (Diptera: Psychodidae) antropofílicos de importancia en salud pública en Los Montes de María, Colombia. Revista Cubana de Medicina Tropical 2009, 61, 220–225. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
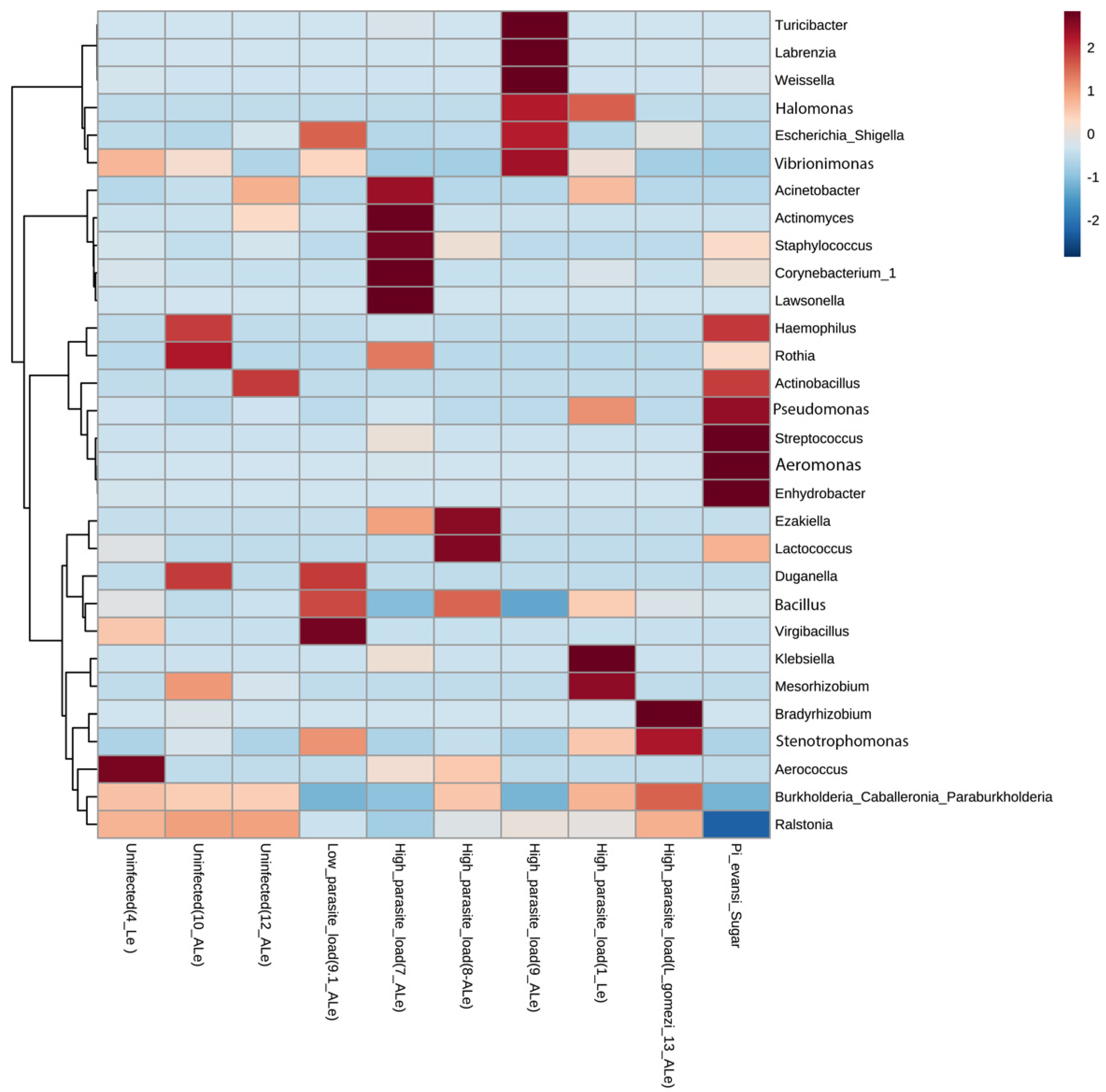{kind=link}
| Treatments | Sandfly | Group Code | No. Guts per Groups | Parasite Load Observed * | DNA Total Concentration (ng/μL) |
|---|---|---|---|---|---|
| Sugar solution (30%) supplemented with an antibiotic cocktail (50 μg/μL) and L. infantum (5 × 106 parasites/mL) | P. evansi | 7-ALe+ | 15 | High | 22.1 |
| 8-ALe+ | 15 | High | 16.3 | ||
| 9-ALe+ | 12 | High | 17.5 | ||
| 9.1-ALe+ | 7 | Low | 23.8 | ||
| 10-ALe− | 10 | Uninfected | 15.1 | ||
| 12-ALe− | 8 | Uninfected | 20.3 | ||
| Sugar solution (30%) and L. infantum (5 × 106 parasites/mL) | 1-Le+ | 5 | High | 19.7 | |
| 4-Le− | 3 | Uninfected | 19.8 | ||
| Sugar solution (30%) supplemented with an antibiotic cocktail (50 μg/μL) and L. infantum (5 × 106 parasites/mL) | L. gomezi | 13-ALe+ | 2 | High | 13.8 |
| P. evansi Coinfection | P. evansi Coinfection | |
|---|---|---|
| Dataset Untreated | Dataset Treated | |
| Total Reads | 1′551.612 | 1′540.250 |
| No. ASVs | 415 | 227 |
| Phyla | (14) | (11) |
| Acidobacteria | Acidobacteria | |
| Actinobacteria | Actinobacteria | |
| Armatimonadetes | Armatimonadetes | |
| Bacteroidetes | Bacteroidetes | |
| Cyanobacteria | Cyanobacteria | |
| Deinococcus–Thermus | Firmicutes | |
| Euglenozoa | Fusobacteria | |
| Firmicutes | Microsporidia | |
| Fusobacteria | Proteobacteria | |
| Microsporidia | Tenericutes | |
| Patescibacteria | Verrucomicrobia | |
| Proteobacteria | ||
| Tenericutes | ||
| Verrucomicrobia | ||
| 5 major families (total counts) | Burkholderiaceae | Burkholderiaceae |
| Bacillaceae | Bacillaceae | |
| Corynebacteriaceae | Corynebacteriaceae | |
| Chitinophagaceae | Chitinophagaceae | |
| Elsteraceae | Elsteraceae | |
| Top 6 most abundant bacterial genera (total counts) | Ralstonia | Ralstonia |
| Bacillus | Bacillus | |
| Burkholderia-Caballeronia-Paraburkholderia | Burkholderia-Caballeronia-Paraburkholderia | |
| Vibrionimonas | Vibrionimonas | |
| CorynebacteriumStaphylococcus | Corynebacterium Staphylococcus | |
| No. taxa summarized to the genus level | 141 | 87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vivero, R.J.; Castañeda-Monsalve, V.A.; Romero, L.R.; D. Hurst, G.; Cadavid-Restrepo, G.; Moreno-Herrera, C.X. Gut Microbiota Dynamics in Natural Populations of Pintomyia evansi under Experimental Infection with Leishmania infantum. Microorganisms 2021, 9, 1214. https://doi.org/10.3390/microorganisms9061214
Vivero RJ, Castañeda-Monsalve VA, Romero LR, D. Hurst G, Cadavid-Restrepo G, Moreno-Herrera CX. Gut Microbiota Dynamics in Natural Populations of Pintomyia evansi under Experimental Infection with Leishmania infantum. Microorganisms. 2021; 9(6):1214. https://doi.org/10.3390/microorganisms9061214
Chicago/Turabian StyleVivero, Rafael José, Victor Alfonso Castañeda-Monsalve, Luis Roberto Romero, Gregory D. Hurst, Gloria Cadavid-Restrepo, and Claudia Ximena Moreno-Herrera. 2021. "Gut Microbiota Dynamics in Natural Populations of Pintomyia evansi under Experimental Infection with Leishmania infantum" Microorganisms 9, no. 6: 1214. https://doi.org/10.3390/microorganisms9061214
APA StyleVivero, R. J., Castañeda-Monsalve, V. A., Romero, L. R., D. Hurst, G., Cadavid-Restrepo, G., & Moreno-Herrera, C. X. (2021). Gut Microbiota Dynamics in Natural Populations of Pintomyia evansi under Experimental Infection with Leishmania infantum. Microorganisms, 9(6), 1214. https://doi.org/10.3390/microorganisms9061214