
First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
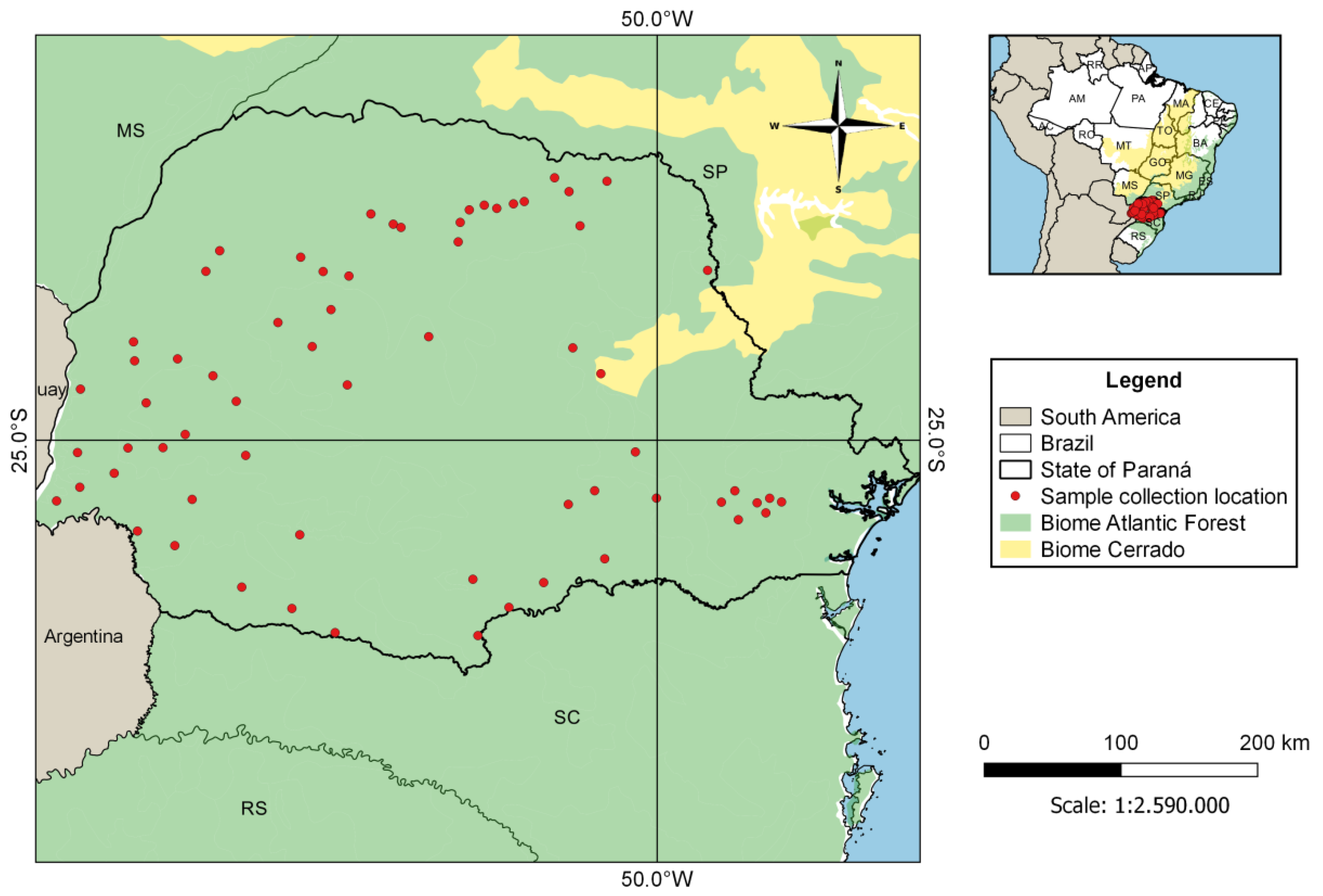
2.1. Sampling
2.2. Polychromophilus Detection
2.3. Host Species Identification
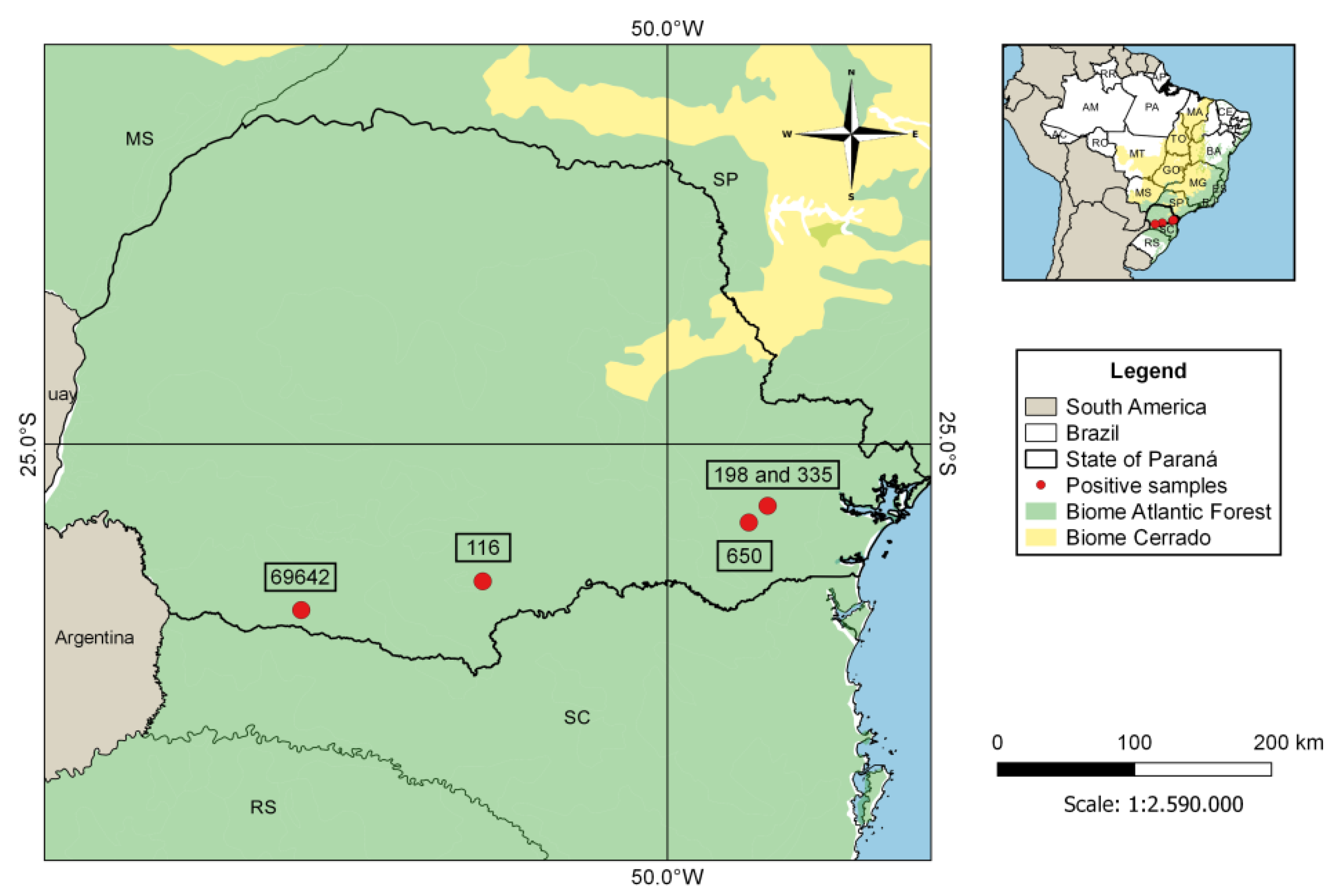
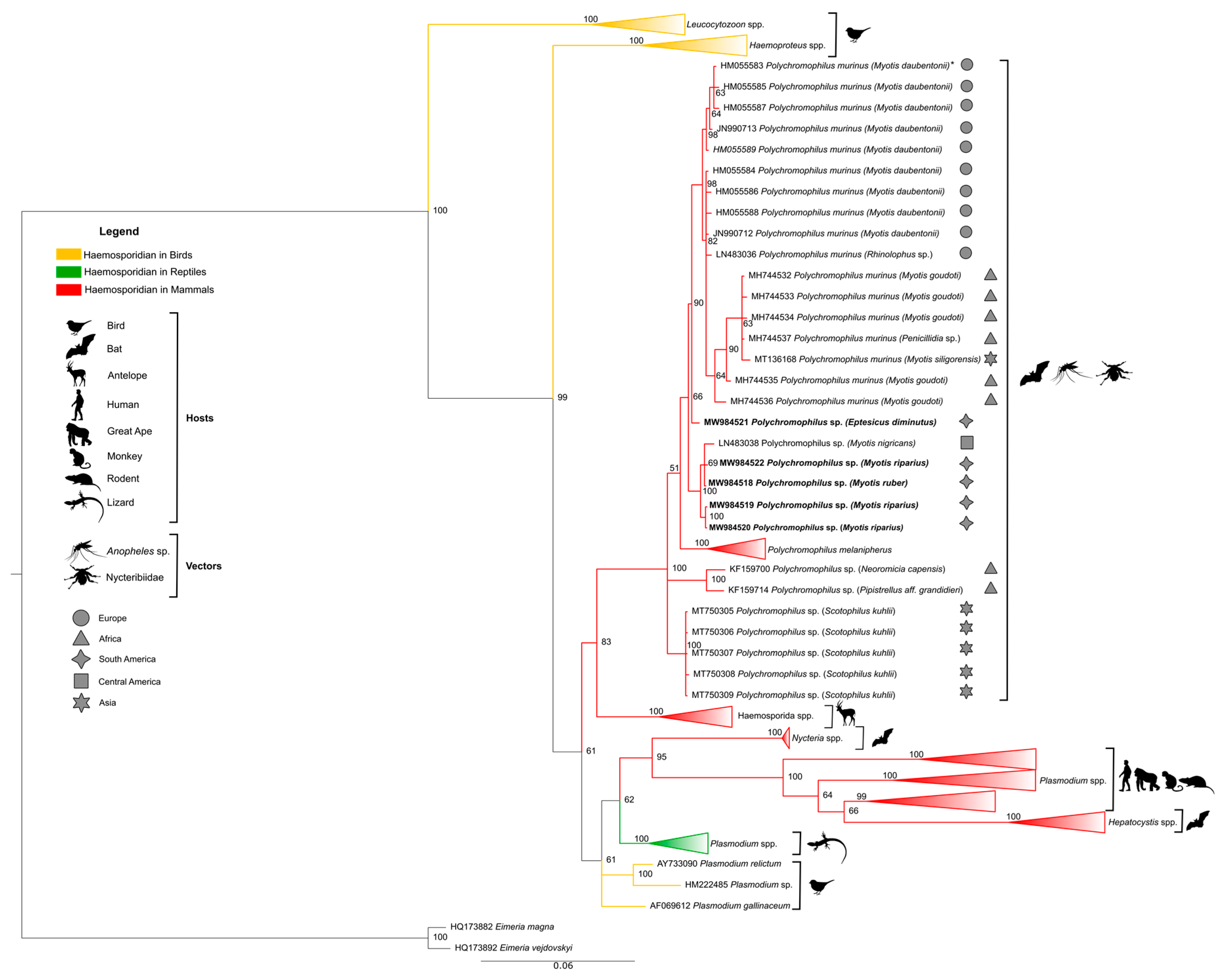
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| GenBank® Accession Number | Parasite Species | Host |
|---|---|---|
| HQ173882 | Eimeria magna | Rabbit |
| HQ173892 | Eimeria vejdovskyi | Rabbit |
| AY099045 | Haemoproteus majoris | Bird |
| HM222472 | Haemoproteus sp. | Bird |
| KT367832, KT367833, KT367822, KT367828 | Haemosporida sp. | Antelope |
| KT367830, KT367819, KT367837 | Haemosporida sp. | Antelope |
| FJ168565 | Hepatocystis sp. | Bat |
| JQ070951, JQ070956 | Hepatocystis sp. | Monkey |
| AY099063 | Leucocytozoon dubreuli | Bird |
| NC_012450, FJ168563 | Leucocytozoon majoris | Bird |
| KF159690, KF159720, MK098843-MK098847 | Nycteria sp. | Bat |
| GQ141581, GQ141585, KT367845, KM598212 | Parahaemoproteus sp. | Bird |
| NC_012447, FJ168561 | Parahaemoproteus vireonis | Bird |
| HM235081 | Plasmodium adleri | Gorilla |
| AY099054, HQ712051 | Plasmodium atheruri | Rodent |
| AY099055 | Plasmodium azurophilum | Lizard |
| KP875474 | Plasmodium billcollinsi | Chimpanzee |
| HM235065 | Plasmodium blacklocki | Gorilla |
| KF159674 | Plasmodium cyclopsi | Bat |
| AB444126 | Plasmodium cynomolgi | Monkey |
| FJ895307 | Plasmodium gaboni | Chimpanzee |
| AF069612 | Plasmodium gallinaceum | Bird |
| AY099053 | Plasmodium giganteum | Lizard |
| JF923751 | Plasmodium gonderi | Mandrill |
| JQ345504 | Plasmodium knowlesi | Human |
| HM000110 | Plasmodium malariae | Chimpanzee |
| GU723548 | Plasmodium ovale | Human |
| JF923762 | Plasmodium praefalciparum | Monkey |
| KP875479 | Plasmodium reichenowi | Chimpanzee |
| AY733090 | Plasmodium relictum | Bird |
| HM222485 | Plasmodium sp. | Bird |
| JF923753 | Plasmodium sp. | Mandrill |
| KJ700853, KJ700854 | Plasmodium vinckei | Rodent |
| KF591834 | Plasmodium vivax | Human |
| KF159671 | Plasmodium voltaicum | Bat |
| DQ414658 | Plasmodium yoelii killicki | Rodent |
| GenBank Accession Number | Parasite Species | Host | Origin |
|---|---|---|---|
| KU318045 | P. melanipherus | Anopheles marshallii | Gabon |
| HM055583 | P. murinus | Myotis daubentonii | Switzerland |
| HM055583 | P. murinus | Eptesicus serotinus | Switzerland |
| HM055583 | P. murinus | Nyctalus noctula | Switzerland |
| HM055583 | P. murinus | Myotis myotis | Switzerland |
| HM055584–HM055589 | P. murinus | Myotis daubentonii | Switzerland |
| MW984521 | Polychromophilus sp. | Eptesicus diminutus | Brazil (this study) |
| KT750375 | Polychromophilus sp. | Miniopterus africanus | Kenya |
| MH744509–MH744511, MH744518, MH744521 | P. melanipherus | Miniopterus gleni | Madagascar |
| MH744506, MH744519 | P. melanipherus | Miniopterus griffithsi | Madagascar |
| MH744514–MH744516 | P. melanipherus | Miniopterus griveaudi | Madagascar |
| MH744508, MH744522–MH744525 | P. melanipherus | Miniopterus griveaudi | Madagascar |
| JQ995284–JQ995288 | Polychromophilus sp. | Miniopterus inflatus | Gabon |
| MH744504, MH744505 | P. melanipherus | Miniopterus mahafaliensis | Madagascar |
| MH744512, MH744526 | P. melanipherus | Miniopterus manavi | Madagascar |
| KT750430 | Polychromophilus sp. | Miniopterus minor | Tanzania |
| MK098848, MK098849 | Polychromophilus sp. | Miniopterus minor | Gabon |
| MW007677 | P. melanipherus | Miniopterus natalensis | South Africa |
| KT750376-KT750382, KT750401, KT750402 | Polychromophilus sp. | Miniopterus natalensis | Kenya |
| KT750406, KT750408, KT750409 | Polychromophilus sp. | Miniopterus natalensis | Kenya |
| MK088162–MK088168 | P. melanipherus | Miniopterus orianae | Australia |
| KT750383-KT750386, KT750415, KT750418 | Polychromophilus sp. | Miniopterus rufus | Kenya |
| JN990708–JN990711 | P. melanipherus | Miniopterus schreibersii | Switzerland |
| KJ131270–KJ131277 | P. melanipherus | Miniopterus schreibersii | Southern and Central Europe |
| MW007689 | P. melanipherus | Miniopterus schreibersii | Spain |
| KT750389 | Polychromophilus sp. | Miniopterus sp. | Tanzania |
| KT750387 | Polychromophilus sp. | Miniopterus sp. | Kenya |
| KF159675, KF159681, KF159699 | Polychromophilus sp. | Miniopterus villiersi | Guinea |
| JN990712, JN990713 | P. murinus | Myotis daubentonii | Switzerland |
| MH744532–MH744536 | P. murinus | Myotis goudoti | Madagascar |
| LN483038 | Polychromophilus sp. | Myotis nigricans | Panamá |
| MW984519, MW984520, MW984522 | Polychromophilus sp. | Myotis riparius | Brazil (this study) |
| MW984518 | Polychromophilus sp. | Myotis ruber | Brazil (this study) |
| MT136168 | P. murinus | Myotis siligorensis | Thailand |
| KF159700 | Polychromophilus sp. | Neoromicia capensis | Guinea |
| MW007685 | P. melanipherus | Nycteribia schmidlii | Spain |
| MW007680, MW007681 | P. melanipherus | Nycteribia schmidlii | Hungary |
| MW007682 | P. melanipherus | Nycteribia schmidlii | Italy |
| MW007671–MW007674, MW007676 | P. melanipherus | Nycteribia schmidlii scotti | South Africa |
| KU182361–KU182367 | P. melanipherus | Nycteribia schmidlii scotti | Gabon |
| MH744527 | P. melanipherus | Nycteribia stylidiopsis | Madagascar |
| MH744520 | P. melanipherus | Paratriaenops furculus | Madagascar |
| KU182368 | P. melanipherus | Penicillidia fulvida | Gabon |
| MH744528–MH744531 | P. melanipherus | Penicillidia leptothrinax | Madagascar |
| MH744537 | P. murinus | Penicillidia sp. | Madagascar |
| KF159714 | Polychromophilus sp. | Pipistrellus aff. grandidieri | Guinea |
| LN483036 | P. murinus | Rhinolophus sp. | Bulgaria |
| MT750305–MT750309 | Polychromophilus sp. | Scotophilus kuhlii | Thailand |
| MT136167 | P. melanipherus | Taphozous melanopogon | Thailand |
References
- Morrison, D.A.; Bornstein, S.; Thebo, P.; Wernery, U.; Kinne, J.; Mattsson, J.G. The current status of the small subunit rRNA phylogeny of the coccidia (Sporozoa). Int. J. Parasitol. 2004, 34, 501–514. [Google Scholar] [CrossRef]
- Gould, S.B.; Tham, W.H.; Cowman, A.F.; Mcfadden, G.I.; Waller, R.F. Alveolins, a new family of cortical proteins that define the protist infrakingdom Alveolata. Mol. Biol. Evol. 2008, 25, 1219–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.S.; Grant, J.; Tekle, Y.I.; Wu, M.; Chaon, B.C.; Cole, J.C.; Logsdon, J.M., Jr.; Patterson, D.J.; Bhattacharya, D.; Katz, L.A. Broadly sampled multigene trees of eukaryotes. BMC Evol. Biol. 2008, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donoghue, P. Haemoprotozoa: Making biological sense of molecular phylogenies. Int. J. Parasitol. Parasites Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef]
- Landau, I.; Chavatte, J.M.; Karadjian, G.; Chabaud, A.; Beveridge, I. The Haemosporidian parasites of bats with description of Sprattiella alecto gen. nov., sp. nov. Parasite 2012, 19, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, S.L.; Schaer, J. A modern menagerie of mammalian malaria. Trends Parasitol. 2016, 32, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Landau, I.; Baccam, D.; Ratanaworabhan, N.; Yenbutra, S.; Boulard, Y.; Chabaud, A.G. Nouveaux Haemoproteidae parasites de Chiroptères en Thailande [New Haemoproteidae parasites of Chiroptera in Thailand]. Ann. Parasitol. Hum. Comp. 1984, 59, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Witsenburg, F.; Salamin, N.; Christe, P. The evolutionary host switches of Polychromophilus: A multi-gene phylogeny of the bat malaria genus suggests a second invasion of mammals by a haemosporidian parasite. Malar. J. 2012, 11, 53. [Google Scholar] [CrossRef]
- Dionisi, A. Les parasites endoglobulaires des chauves-souris. Atti Reale Acad. Lincei 1898, 7, 153–156. [Google Scholar]
- Dionisi, A. Un parassita del globulo rosso in una specie di pipistrello (Miniopterus Schreibersii Kuhl). Atti Reale Acad. Lincei 1898, 7, 214–215. [Google Scholar]
- Garnham, P.C.C. Polychromophilus species in insectivorous bats. Trans. R. Soc. Trop. Med. Hyg. 1973, 67, 2–3. [Google Scholar] [CrossRef]
- Garnham, P.C.C. The zoogeography of Polychromophilus and description of a new species of a gregarine (Lankesteria galliardi). Ann. Parasitol. 1973, 48, 231–242. [Google Scholar] [CrossRef]
- Landau, I.; Rosin, G.; Miltgen, F. The genus Polychromophilus (Haemoproteidae, parasite of Microchiroptera). Ann. Parasitol. Hum. Comp. 1980, 55, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Garnham, P.C.C. Malaria Parasites and Other Haemosporidia; Blackwell: Oxford, UK, 1966. [Google Scholar]
- Schaer, J.; Perkins, S.L.; Decher, J.; Leendertz, F.H.; Fahr, J.; Weber, N.; Matuschewski, K. High diversity of West African bat malaria parasites and a tight link with rodent Plasmodium taxa. Proc. Natl. Acad. Sci. USA 2013, 43, 17415–17419. [Google Scholar] [CrossRef] [Green Version]
- Schaer, J.; Perkins, S.L.; Ejotre, I.; Vodzak, M.E.; Matuschewski, K.; Reeder, D.M. Epauletted fruit bats display exceptionally high infections with a Hepatocystis species complex in South Sudan. Sci. Rep. 2017, 7, 6928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, H.L.; Patterson, B.D.; Kerbis Peterhans, J.C.; Stanley, W.T.; Webala, P.W.; Gnoske, T.P.; Hackett, S.J.; Stanhope, M.J. Diverse sampling of East African haemosporidians reveals chiropteran origin of malaria parasites in primates and rodents. Mol. Phylogenet. Evol. 2016, 99, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Garnham, P.C.C.; Lainson, R.; Shaw, J.J. A malaria-like parasite of a bat from Brazil. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 13. [Google Scholar] [PubMed]
- Garnham, P.C.C.; Lainson, R.; Shaw, J.J. A contribution to the study of the haematozoon parasites of bats. A new mammalian haemoproteid, Polychromophilus deanei n. sp. Mem. Inst. Oswaldo Cruz 1971, 69, 119–127. [Google Scholar] [CrossRef]
- Borner, J.; Pick, C.; Thiede, J.; Kolawole, O.M.; Kingsley, M.T.; Schulze, J.; Cottontail, V.M.; Wellinghausen, N.; Schmidt-Chanasit, J.; Bruchhaus, I.; et al. Phylogeny of haemosporidian blood parasites revealed by a multi-gene approach. Mol. Phylogenet. Evol. 2016, 94, 221–231. [Google Scholar] [CrossRef]
- Perkins, S.L.; Schall, J.J. A molecular phylogeny of malarial parasites recovered from cytochrome b gene sequences. J. Parasitol. 2002, 88, 972–978. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree: Tree Figure Drawing Tool Version 1.4.0; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, N.V.; de Waard, J.R.; Hebert, P.D.N. An inexpensive, automation-friendly protocol for recovering high-quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, N.; Sharma, A. DNA barcoding of the Indian blackbuck (Antilope cervicapra) and their correlation with other closely related species. Egypt J. Forensic. Sci. 2017, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Ruiz, F.; Linton, Y.M.; Ponsonby, D.J.; Conn, J.E.; Herrera, M.; Quiñones, M.L.; Vélez, I.D.; Wilkerson, R.C. Molecular comparison of topotypic specimens confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem. Inst. Oswaldo Cruz 2010, 105, 899–903. [Google Scholar] [CrossRef]
- Miretzki, M. Morcegos do Estado do Paraná, Brasil (Mammalia, Chiroptera): Riqueza de espécies, distribuição e síntese do conhecimento atual. Pap. Avulsos Zool. 2003, 43, 101–138. [Google Scholar] [CrossRef]
- Holz, P.H.; Lumsden, L.F.; Legione, A.R.; Hufschmid, J. Polychromophilus melanipherus and haemoplasma infections not associated with clinical signs in southern bent-winged bats (Miniopterus orianae bassanii) and eastern bent-winged bats (Miniopterus orianae oceanensis). Int. J. Parasitol. Parasites Wildl. 2018, 8, 10–18. [Google Scholar] [CrossRef]
- Witsenburg, F.; Clément, L.; López-Baucells, A.; Palmeirim, J.; Pavlinić, I.; Scaravelli, D.; Ševčík, M.; Dutoit, L.; Salamin, N.; Goudet, J.; et al. How a haemosporidian parasite of bats gets around: The genetic structure of a parasite, vector and host compared. Mol. Ecol. 2015, 24, 926–940. [Google Scholar] [CrossRef] [Green Version]
- Duval, L.; Mejean, C.; Maganga, G.D.; Makanga, B.K.; Mangama Koumba, L.B.; Peirce, M.A.; Ariey, F.; Bourgarel, M.; Bourgarel, M. The chiropteran haemosporidian Polychromophilus melanipherus: A worldwide species complex restricted to the family Miniopteridae. Infec. Genet. Evol. 2012, 12, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Rosskopf, S.P.; Held, J.; Gmeiner, M.; Mordmüller, B.; Matsiégui, P.; Eckerle, I.; Weber, N.; Matuschewski, K.; Schaer, J. Nycteria and Polychromophilus parasite infections of bats in Central Gabon. Infect. Genet. Evol. 2019, 68, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Ramasindrazana, B.; Goodman, S.M.; Dsouli, N.; Gomard, Y.; Lagadec, E.; Randrianarivelojosia, M.; Dellagi, K.; Tortosa, P. Polychromophilus spp. (Haemosporida) in Malagasy bats: Host specificity and insights on invertebrate vectors. Malar. J. 2018, 17, 318. [Google Scholar] [CrossRef]
- Rasoanoro, M.; Goodman, S.M.; Randrianarivelojosia, M.; Rakotondratsimba, M.; Dellagi, K.; Tortosa, P.; Ramasindrazana, B. Diversity, distribution, and drivers of Polychromophilus infection in Malagasy bats. Malar. J. 2021, 20, 157. [Google Scholar] [CrossRef]
- Megali, A.; Yannic, G.; Christe, P. Disease in the dark: Molecular characterization of Polychromophilus murinus in temperate zone bats revealed a worldwide distribution of this malaria-like disease. Mol. Ecology 2011, 20, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Chumnandee, C.; Pha-obnga, N.; Werb, O.; Matuschewski, K.; Schaer, J. Molecular characterization of Polychromophilus parasites of Scotophilus kuhlii bats in Thailand. Parasitology 2020, 1–5. [Google Scholar] [CrossRef]
- Arnuphapprasert, A.; Riana, E.; Ngamprasertwong, T.; Wangthongchaicharoen, M.; Soisook, P.; Thanee, S.; Bhodhibundit, P.; Kaewthamasorn, M. First molecular investigation of haemosporidian parasites in Thai bat species. Int. J. Parasitol. Parasites Wildl. 2020, 13, 51–61. [Google Scholar] [CrossRef]
- Reis, N.R.; Peracchi, A.L.; Pedro, W.A.; Lima, I.P. Morcegos do Brasil; Nélio, R. Reis: Londrina, Brazil, 2007; p. 253. [Google Scholar]
- Sekiama, M.L.; Reis, N.R.; Peracchi, A.L.; Rocha, V.J. Morcegos do Parque Nacional do Iguaçu, Paraná (Chiroptera, Mammalia). Rev. Bras. Zool. 2001, 18, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Bianconi, G.V.; Mikich, S.B.; Pedro, W.A. Diversidade de morcegos (Mammalia, Chiroptera) em remanescentes florestais do município de Fênix, noroeste do Paraná, Brasil. Rev. Bras. Zool. 2004, 21, 943–954. [Google Scholar] [CrossRef]
- Machado, A.B.M.; Martins, C.S.; Drummond, G.M. Lista da Fauna Brasileira Ameaçada de Extinção: Incluindo a Lista de Espécies Quase Ameaçadas e Deficientes em Dados; Fundação Biodiversitas: Belo Horizonte, Brazil, 2005; p. 158. [Google Scholar]
- IUCN. IUCN Red List of Threatened Species. Version 2021.1. 2019. Available online: http://www.iucnredlist.org (accessed on 15 April 2021).
- Barquez, R.M.; Mares, M.A.; Braun, J.K. The bats of Argentina. Spec. Publ. Texas Tech Univ. 1999, 42, 1–275. [Google Scholar]
- Lopez-Gonzalez, C.; Presley, S.J.; Owen, R.D.; Willig, M.R. Taxonomic status of Myotis (Chiroptera: Vespertilionidae) in Paraguay. J. Mammal. 2001, 82, 138–160. [Google Scholar] [CrossRef] [Green Version]
- Achaval, F.; Clara, M.; Olmos, A. Mamiferos de la Republica Oriental del Uruguay; Biophoto: Montevideo, Uruguay, 2008; p. 213. [Google Scholar]
- Weber, M.M.; Terribile, L.C.; Caceres, N.C. Potential geographic distribution of Myotis ruber (Chiroptera, Vespertilionidae), a threatened Neotropical bat species. Mammalia 2010, 74, 333–338. [Google Scholar] [CrossRef]
- Thurber, M.I.; Ghai, R.R.; Hyeroba, D.; Weny, G.; Tumukunde, A.; Chapman, C.A.; Wiseman, R.W.; Dinis, J.; Steeil, J.; Greiner, E.C.; et al. Co-infection and cross-species transmission of divergent Hepatocystis lineages in a wild African primate community. Int. J. Parasitol. 2013, 43, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinsen, E.S.; Paperna, I.; Schall, J.J. Morphological versus molecular identification of avian Haemosporidia: An exploration of three species concepts. Parasitology 2006, 133, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javanbakht, H.; Kvičerová, J.; Dvořáková, N.; Mikulíček, P.; Sharifi, M.; Kautman, M.; Maršíková, A.; Široký, P. Phylogeny, Diversity, Distribution, and Host Specificity of Haemoproteus spp. (Apicomplexa: Haemosporida: Haemoproteidae) of Palaearctic Tortoises. J. Eukaryot Microbiol. 2015, 62, 670–678. [Google Scholar] [CrossRef]
- Martinsen, E.S.; Perkins, S.L.; Schall, J.J. A three-genome phylogeny of malaria parasites (Plasmodium and closely related genera): Evolution of life-history traits and host switches. Mol. Phylogenet Evol. 2008, 47, 261–273. [Google Scholar] [CrossRef]



| Isolate | 219 | 247 | 261 | 273 | 339 | 405 | 512 | 789 | 792 | 810 | 811 | 853 | 885 | 945 | 1086 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 116 | C | T | A | T | T | T | T | T | C | C | T | C | A | T | A |
| 198 | C | T | A | T | G | T | T | T | T | C | T | C | A | T | G |
| 335 | C | T | A | T | G | T | T | T | T | C | T | C | A | T | G |
| 650 | T | T | C | A | A | C | T | C | T | T | C | T | T | T | A |
| 69642 | C | T | A | T | T | T | T | T | C | C | T | C | A | C | A |
| MYOPA01 | C | C | A | T | T | T | G |
| Bat Species | 116 | 198 | 335 | 650 | 69642 | MYOPA01 | |
|---|---|---|---|---|---|---|---|
| 116 | Myotis ruber | 1116 | 99% | 99% | 99% | 99% | 99% |
| 198 | Myotis riparius | 1113 | 1116 | 100% | 99% | 99% | 99% |
| 335 | Myotis riparius | 1113 | 1116 | 1116 | 99% | 99% | 99% |
| 650 | Eptesicus diminutus | 1105 | 1105 | 1105 | 1116 | 98% | 98% |
| 69642 | Myotis riparius | 1115 | 1112 | 1112 | 1104 | 1116 | 99% |
| MYOPA01 | Myotis nigricans | 592 | 591 | 591 | 587 | 592 | 595 |
| Country or Continent | Analyzed Samples | Positive Samples (Positivity) | Positive Host Species | Reference |
|---|---|---|---|---|
| Africa 1 | 505 | 56 (11%) | Miniopterus africanus, M. fraterculus, M. minor, M. natalensis, M. rufus, Myotis tricolor | [17] |
| Australia 2 | 85 | 47 (55%) | Miniopterus orianae | [32] |
| Brazil 3 | 406 | 5 (1.2%) | Eptesicus diminutus, Myotis ruber, Myotis riparius | This study |
| Europe 4 | 310 | 231 (74.5%) | Miniopterus schreibersii | [33] |
| Gabon | 164 | 5 (3%) | Miniopterus inflatus | [34] |
| Gabon | 92 | 2 (2%) | Miniopterus minor | [35] |
| Guinea | 274 | 5 (2%) | Miniopterus villiersi, Neoromicia capensis, Pipistrellus aff. grandidieri | [15] |
| Madagascar | 947 | 130 (13.5%) | Paratriaenops furculus, Miniopterus aelleni, M. manavi, M. gleni, M. grifthsi, M. griveaudi, M. mahafaliensis, M. majori, M. sororculus, Myotis goudoti | [36] |
| Madagascar | 222 | 27 (12.2%) | Miniopterus egeri, M. griveaudi, M. ambohitrensis, M. gleni, Scotophilus robustus, Myotis goudoti | [37] |
| Switzerland | 207 | 70 (34%) | Myotis daubentonii, M. myotis, Nyctalus noctula, Eptesicus serotinus | [38] |
| Thailand | 44 | 5 (11%) | Scotophilus kuhlii | [39] |
| Thailand | 271 | 13 (4.8%) | Myotis siligorensis, Taphozous melanopogon | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minozzo, G.A.; da Silva Mathias, B.; Riediger, I.N.; de Oliveira Guimarães, L.; dos Anjos, C.C.; Monteiro, E.F.; dos Santos, A.P.; Biondo, A.W.; Kirchgatter, K. First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species. Microorganisms 2021, 9, 1240. https://doi.org/10.3390/microorganisms9061240
Minozzo GA, da Silva Mathias B, Riediger IN, de Oliveira Guimarães L, dos Anjos CC, Monteiro EF, dos Santos AP, Biondo AW, Kirchgatter K. First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species. Microorganisms. 2021; 9(6):1240. https://doi.org/10.3390/microorganisms9061240
Chicago/Turabian StyleMinozzo, Guilherme Augusto, Bruno da Silva Mathias, Irina Nastassja Riediger, Lilian de Oliveira Guimarães, Carolina Clares dos Anjos, Eliana Ferreira Monteiro, Andrea Pires dos Santos, Alexander Welker Biondo, and Karin Kirchgatter. 2021. "First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species" Microorganisms 9, no. 6: 1240. https://doi.org/10.3390/microorganisms9061240
APA StyleMinozzo, G. A., da Silva Mathias, B., Riediger, I. N., de Oliveira Guimarães, L., dos Anjos, C. C., Monteiro, E. F., dos Santos, A. P., Biondo, A. W., & Kirchgatter, K. (2021). First Molecular Detection of Polychromophilus Parasites in Brazilian Bat Species. Microorganisms, 9(6), 1240. https://doi.org/10.3390/microorganisms9061240