
Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Microbiota-Depleted IL-10−/− Mice
2.3. Campylobacter Infection and Treatment Regimens
2.4. Gastrointestinal Pathogen Burdens
2.5. Monitoring of Clinical Conditions of Animals
2.6. Sampling Procedures
2.7. Histopathology
2.8. In Situ Immunohistochemistry
2.9. Pro-Inflammatory Mediators
2.10. Statistics
3. Results
3.1. Gastrointestinal Pathogen Numbers Following Treatment of Infected IL-10−/− Mice with Activated Charcoal
3.2. Clinical Outcome Following Treatment of Infected Mice with Activated Charcoal
3.3. Microscopic Inflammatory Changes in the Colon Following Treatment of Infected Mice with Activated Charcoal
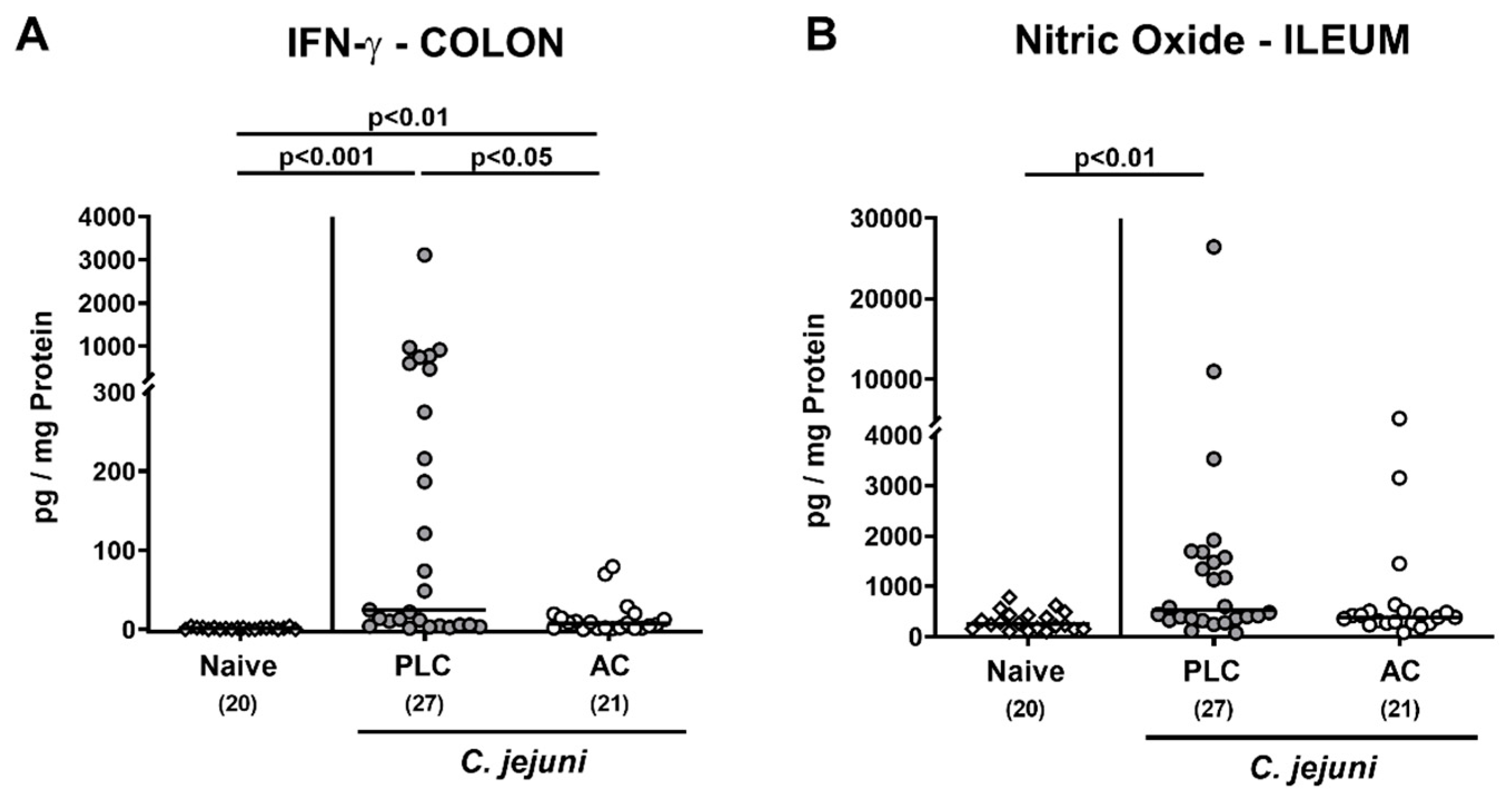
3.4. Intestinal Pro-Inflammatory Immune Responses following Treatment of Infected Mice with Activated Charcoal
3.5. Pro-Inflammatory Mediator Secretion in Extra-Intestinal Including Systemic Organs Following Treatment of Infected Mice with Activated Charcoal
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organisation). Campylobacter. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 4 June 2020).
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human Campylobacteriosis-A Serious Infectious Threat in a One Health Perspective. Curr. Top. Microbiol. Immunol. 2021, 431, 1–23. [Google Scholar] [PubMed]
- Alter, T.; Reich, F. Management Strategies for Prevention of Campylobacter Infections Through the Poultry Food Chain: A European Perspective. In Fighting Campylobacter Infections: Towards a One Health Approach; Springer: Cham, Switzerland, 2021; pp. 79–102. [Google Scholar]
- Mortensen, N.; Jonasson, S.A.; Lavesson, I.V.; Emberland, K.E.; Litleskare, S.; Wensaas, K.-A.; Rortveit, G.; Langeland, N.; Hanevik, K. Characteristics of hospitalized patients during a large waterborne outbreak of Campylobacter jejuni in Norway. PLoS ONE 2021, 16, e0248464. [Google Scholar] [CrossRef]
- Karmali, M.; Fleming, P. Campylobacter enteritis. Can. Med. Assoc. J. 1979, 120, 1525. [Google Scholar] [PubMed]
- Backert, S.; Tegtmeyer, N.; Cróinín, T.Ó.; Boehm, M.; Heimesaat, M.M. Chapter 1—Human campylobacteriosis. In Campylobacter; Klein, G., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 1–25. [Google Scholar] [CrossRef]
- Yuki, N. Pathogenesis of Guillain-Barré and Miller Fisher syndromes subsequent to Campylobacter jejuni enteritis. Jpn. J. Infect. Dis. 1999, 52, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.M.; Flint, N.J.; Livsey, S.A.; Wong, R.; Spiers, P.; Bukhari, S.S. Bickerstaff’s brainstem encephalitis related to Campylobacter jejuni gastroenteritis. J. Clin. Pathol. 2007, 60, 1161–1162. [Google Scholar] [CrossRef]
- Kirkpatrick, B.D.; Tribble, D.R. Update on human Campylobacter jejuni infections. Curr. Opin. Gastroenterol. 2011, 27, 1–7. [Google Scholar] [CrossRef]
- Mortensen, N.P.; Kuijf, M.L.; Ang, C.W.; Schiellerup, P.; Krogfelt, K.A.; Jacobs, B.C.; van Belkum, A.; Endtz, H.P.; Bergman, M.P. Sialylation of Campylobacter jejuni lipo-oligosaccharides is associated with severe gastro-enteritis and reactive arthritis. Microbes Infect. 2009, 11, 988–994. [Google Scholar] [CrossRef]
- Louwen, R.; Heikema, A.; van Belkum, A.; Ott, A.; Gilbert, M.; Ang, W.; Endtz, H.P.; Bergman, M.P.; Nieuwenhuis, E.E. The sialylated lipooligosaccharide outer core in Campylobacter jejuni is an important determinant for epithelial cell invasion. Infect. Immun. 2008, 76, 4431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Novel Clinical Campylobacter jejuni Infection Models Based on Sensitization of Mice to Lipooligosaccharide, a Major Bacterial Factor Triggering Innate Immune Responses in Human Campylobacteriosis. Microorganisms 2020, 8, 482. [Google Scholar] [CrossRef] [Green Version]
- Price, A.B.; Jewkes, J.; Sanderson, P.J. Acute diarrhoea: Campylobacter colitis and the role of rectal biopsy. J. Clin. Pathol. 1979, 32, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Tegtmeyer, N.; Sharafutdinov, I.; Harrer, A.; Esmaeili, D.S.; Linz, B.; Backert, S. Campylobacter Virulence Factors and Molecular Host–Pathogen Interactions. In Fighting Campylobacter Infections: Towards a One Health Approach; Springer: Cham, Switzerland, 2021; pp. 169–202. [Google Scholar]
- Hickey, T.E.; McVeigh, A.L.; Scott, D.A.; Michielutti, R.E.; Bixby, A.; Carroll, S.A.; Bourgeois, A.L.; Guerry, P. Campylobacter jejuni cytolethal distending toxin mediates release of interleukin-8 from intestinal epithelial cells. Infect. Immun. 2000, 68, 6535–6541. [Google Scholar] [CrossRef] [Green Version]
- Larson, C.L.; Shah, D.H.; Dhillon, A.S.; Call, D.R.; Ahn, S.; Haldorson, G.J.; Davitt, C.; Konkel, M.E. Campylobacter jejuni invade chicken LMH cells inefficiently and stimulate differential expression of the chicken CXCLi1 and CXCLi2 cytokines. Microbiology 2008, 154, 3835–3847. [Google Scholar] [CrossRef] [Green Version]
- de Sá, F.L.; Schulzke, J.-D.; Bücker, R. Diarrheal mechanisms and the role of intestinal barrier dysfunction in Campylobacter infections. Curr. Top. Microbiol. Immunol. 2021, 431, 203–231. [Google Scholar]
- Bücker, R.; Krug, S.M.; Moos, V.; Bojarski, C.; Schweiger, M.R.; Kerick, M.; Fromm, A.; Janssen, S.; Fromm, M.; Hering, N.A.; et al. Campylobacter jejuni impairs sodium transport and epithelial barrier function via cytokine release in human colon. Mucosal. Immunol. 2018, 11, 575–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, L.-M.; Fischer, A.; Otto, B.; Plickert, R.; Kühl, A.A.; Göbel, U.B.; Bereswill, S.; Heimesaat, M.M. Campylobacter jejuni induces acute enterocolitis in gnotobiotic IL-10−/− mice via Toll-like-receptor-2 and-4 signaling. PloS ONE 2012, 7, e40761. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Threadgill, D.; Jobin, C. Campylobacter jejuni induces colitis through activation of mammalian target of rapamycin signaling. Gastroenterology 2012, 142, 86–95.e85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Murine Models for the Investigation of Colonization Resistance and Innate Immune Responses in Campylobacter Jejuni Infections. Curr. Top. Microbiol. Immunol. 2021, 431, 233–263. [Google Scholar]
- Mousavi, S.; Escher, U.; Thunhorst, E.; Kittler, S.; Kehrenberg, C.; Bereswill, S.; Heimesaat, M.M. Vitamin C alleviates acute enterocolitis in Campylobacter jejuni infected mice. Sci. Rep. 2020, 10, 2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Lobo de Sa, F.D.; Schulzke, J.D.; Bucker, R.; Bereswill, S.; Heimesaat, M.M. Vitamin D in Acute Campylobacteriosis-Results From an Intervention Study Applying a Clinical Campylobacter jejuni Induced Enterocolitis Model. Front. Immunol. 2019, 10, 2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Schmidt, A.-M.; Escher, U.; Kittler, S.; Kehrenberg, C.; Thunhorst, E.; Bereswill, S.; Heimesaat, M.M. Carvacrol ameliorates acute campylobacteriosis in a clinical murine infection model. Gut Pathog. 2020, 12, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimesaat, M.M.; Mousavi, S.; Escher, U.; de Sá, F.D.L.; Peh, E.; Schulzke, J.-D.; Kittler, S.; Bücker, R.; Bereswill, S. Resveratrol Alleviates Acute Campylobacter jejuni Induced Enterocolitis in a Preclinical Murine Intervention Study. Microorganisms 2020, 8, 1858. [Google Scholar] [CrossRef]
- De Sá, F.D.L.; Heimesaat, M.M.; Bereswill, S.; Nattramilarasu, P.K.; Schulzke, J.-D.; Bücker, R. Resveratrol prevents Campylobacter jejuni-induced leaky gut by restoring occludin and claudin-5 in the paracellular leak pathway. Front. Pharmacol. 2021, 12, 454. [Google Scholar]
- Mousavi, S.; Weschka, D.; Bereswill, S.; Heimesaat, M.M. Preclinical Evaluation of Oral Urolithin-A for the Treatment of Acute Campylobacteriosis in Campylobacter jejuni Infected Microbiota-Depleted IL-10−/− Mice. Pathogens 2021, 10, 7. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Anti-Pathogenic and Immune-Modulatory Effects of Peroral Treatment with Cardamom Essential Oil in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 169. [Google Scholar] [CrossRef]
- Bereswill, S.; Mousavi, S.; Weschka, D.; Buczkowski, A.; Schmidt, S.; Heimesaat, M.M. Peroral Clove Essential Oil Treatment Ameliorates Acute Campylobacteriosis-Results from a Preclinical Murine Intervention Study. Microorganisms 2021, 9, 735. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Garlic Essential Oil as Promising Option for the Treatment of Acute Campylobacteriosis—Results from a Preclinical Placebo-Controlled Intervention Study. Microorganisms 2021, 9, 1140. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Kløve, S.; Genger, C.; Weschka, D.; Giladi, E.; Bereswill, S.; Gozes, I. Immune-modulatory Properties of the Octapeptide NAP in Campylobacter jejuni Infected Mice Suffering from Acute Enterocolitis. Microorganisms 2020, 8, 802. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Kløve, S.; Genger, C.; Weschka, D.; Tamas, A.; Reglodi, D.; Bereswill, S. Pituitary Adenylate Cyclase-Activating Polypeptide Alleviates Intestinal, Extra-Intestinal and Systemic Inflammatory Responses during Acute Campylobacter jejuni-induced Enterocolitis in Mice. Pathogens 2020, 9, 805. [Google Scholar] [CrossRef]
- Ekmekciu, I.; Fiebiger, U.; Stingl, K.; Bereswill, S.; Heimesaat, M.M. Amelioration of intestinal and systemic sequelae of murine Campylobacter jejuni infection by probiotic VSL#3 treatment. Gut Pathog. 2017, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimesaat, M.M.; Mrazek, K.; Bereswill, S. Murine Fecal Microbiota Transplantation Alleviates Intestinal and Systemic Immune Responses in Campylobacter jejuni Infected Mice Harboring a Human Gut Microbiota. Front. Immunol. 2019, 10, 2272. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mrazek, K.; Bereswill, S. Murine fecal microbiota transplantation lowers gastrointestinal pathogen loads and dampens pro-inflammatory immune responses in Campylobacter jejuni infected secondary abiotic mice. Sci. Rep. 2019, 9, 19797. [Google Scholar] [CrossRef]
- Juurlink, D.N. Activated charcoal for acute overdose: A reappraisal. Br. J. Clin. Pharmacol. 2016, 81, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Senderovich, H.; Vierhout, M.J. Is there a role for charcoal in palliative diarrhea management? Curr. Med Res. Opin. 2018, 34, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Nagaki, M.; Hughes, R.; Lau, J.; Williams, R. Removal of endotoxin and cytokines by adsorbents and the effect of plasma protein binding. Int. J. Artif. Organs 1991, 14, 43–50. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Bereswill, S.; Fischer, A.; Fuchs, D.; Struck, D.; Niebergall, J.; Jahn, H.-K.; Dunay, I.R.; Moter, A.; Gescher, D.M.; et al. Gram-negative bacteria aggravate murine small intestinal Th1-type immunopathology following oral infection with Toxoplasma gondii. J. Immunol. 2006, 177, 8785–8795. [Google Scholar] [CrossRef] [Green Version]
- Bereswill, S.; Fischer, A.; Plickert, R.; Haag, L.-M.; Otto, B.; Kuhl, A.A.; Dasti, J.I.; Zautner, A.E.; Munoz, M.; Loddenkemper, C.; et al. Novel murine infection models provide deep insights into the “menage a trois” of Campylobacter jejuni, microbiota and host innate immunity. PLoS ONE 2011, 6, e20953. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Haag, L.M.; Fischer, A.; Otto, B.; Kuhl, A.A.; Gobel, U.B.; Bereswill, S. Survey of extra-intestinal immune responses in asymptomatic long-term Campylobacter jejuni-infected mice. Eur. J. Microbiol. Immunol. 2013, 3, 174–182. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Alutis, M.; Grundmann, U.; Fischer, A.; Tegtmeyer, N.; Bohm, M.; Kuhl, A.A.; Gobel, U.B.; Backert, S.; Bereswill, S. The role of serine protease HtrA in acute ulcerative enterocolitis and extra-intestinal immune responses during Campylobacter jejuni infection of gnotobiotic IL-10 deficient mice. Front. Cell. Infect. Microbiol. 2014, 4, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar] [PubMed]
- Heimesaat, M.M.; Giladi, E.; Kuhl, A.A.; Bereswill, S.; Gozes, I. The octapetide NAP alleviates intestinal and extra-intestinal anti-inflammatory sequelae of acute experimental colitis. Peptides 2018, 101, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramu, J.; Clark, K.; Woode, G.N.; Sarr, A.B.; Phillips, T.D. Adsorption of cholera and heat-labile Escherichia coli enterotoxins by various adsorbents: An in vitro study. J. Food Prot. 1997, 60, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Rollinger, Y.; Dott, W. Survival of selected bacterial species in sterilized activated carbon filters and biological activated carbon filters. Appl. Environ. Microbiol. 1987, 53, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, R.A.; Hunt, J.; Sanders, E.; Tran, M.; Burk, G.A.; Mlsna, T.E.; Fitzkee, N.C. Effect of biochar on microbial growth: A metabolomics and bacteriological investigation in Escherichia coli. Environ. Sci. Technol. 2019, 53, 2635–2646. [Google Scholar] [CrossRef]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.; Alwen, J. Effect of activated charcoal on the swarming of proteus. Nature 1966, 212, 941–942. [Google Scholar] [CrossRef]
- Karmali, M.A.; Simor, A.; Roscoe, M.; Fleming, P.; Smith, S.; Lane, J. Evaluation of a blood-free, charcoal-based, selective medium for the isolation of Campylobacter organisms from feces. J. Clin. Microbiol. 1986, 23, 456. [Google Scholar] [CrossRef] [Green Version]
- Grall, N.; Massias, L.; Nguyen, T.T.; Sayah-Jeanne, S.; Ducrot, N.; Chachaty, E.; de Gunzburg, J.; Andremont, A. Oral DAV131, a charcoal-based adsorbent, inhibits intestinal colonization by β-lactam-resistant Klebsiella pneumoniae in cefotaxime-treated mice. Antimicrob. Agents Chemother. 2013, 57, 5423. [Google Scholar] [CrossRef] [Green Version]
- Naka, K.; Watarai, S.; Inoue, K.; Kodama, Y.; Oguma, K.; Yasuda, T.; Kodama, H. Adsorption effect of activated charcoal on enterohemorrhagic Escherichia coli. J. Vet. Med. Sci. 2001, 63, 281–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bysani, G.K.; Shenep, J.L.; Hildner, W.K.; Stidham, G.L.; Roberson, P.K. Detoxification of plasma containing lipopolysaccharide by adsorption. Crit. Care Med. 1990, 18, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.S.; Pine, L.; Bell, S. Production of superoxide and hydrogen peroxide in medium used to culture Legionella pneumophila: Catalytic decomposition by charcoal. Appl. Environ. Microbiol. 1983, 45, 784–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrin, M.R.; Arumugam, S.; Rahman, M.A.; Karuppagounder, V.; Sreedhar, R.; Harima, M.; Suzuki, H.; Nakamura, T.; Miyashita, S.; Suzuki, K.; et al. Le Carbone, a charcoal supplement, modulates DSS-induced acute colitis in mice through activation of AMPKα and downregulation of STAT3 and caspase 3 dependent apoptotic pathways. Int. Immunopharmacol. 2017, 43, 70–78. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bereswill, S.; Mousavi, S.; Weschka, D.; Heimesaat, M.M. Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 1424. https://doi.org/10.3390/microorganisms9071424
Bereswill S, Mousavi S, Weschka D, Heimesaat MM. Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis. Microorganisms. 2021; 9(7):1424. https://doi.org/10.3390/microorganisms9071424
Chicago/Turabian StyleBereswill, Stefan, Soraya Mousavi, Dennis Weschka, and Markus M. Heimesaat. 2021. "Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis" Microorganisms 9, no. 7: 1424. https://doi.org/10.3390/microorganisms9071424
APA StyleBereswill, S., Mousavi, S., Weschka, D., & Heimesaat, M. M. (2021). Disease-Alleviating Effects of Peroral Activated Charcoal Treatment in Acute Murine Campylobacteriosis. Microorganisms, 9(7), 1424. https://doi.org/10.3390/microorganisms9071424