
Free-Living Amoebae in Soil Samples from Santiago Island, Cape Verde

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
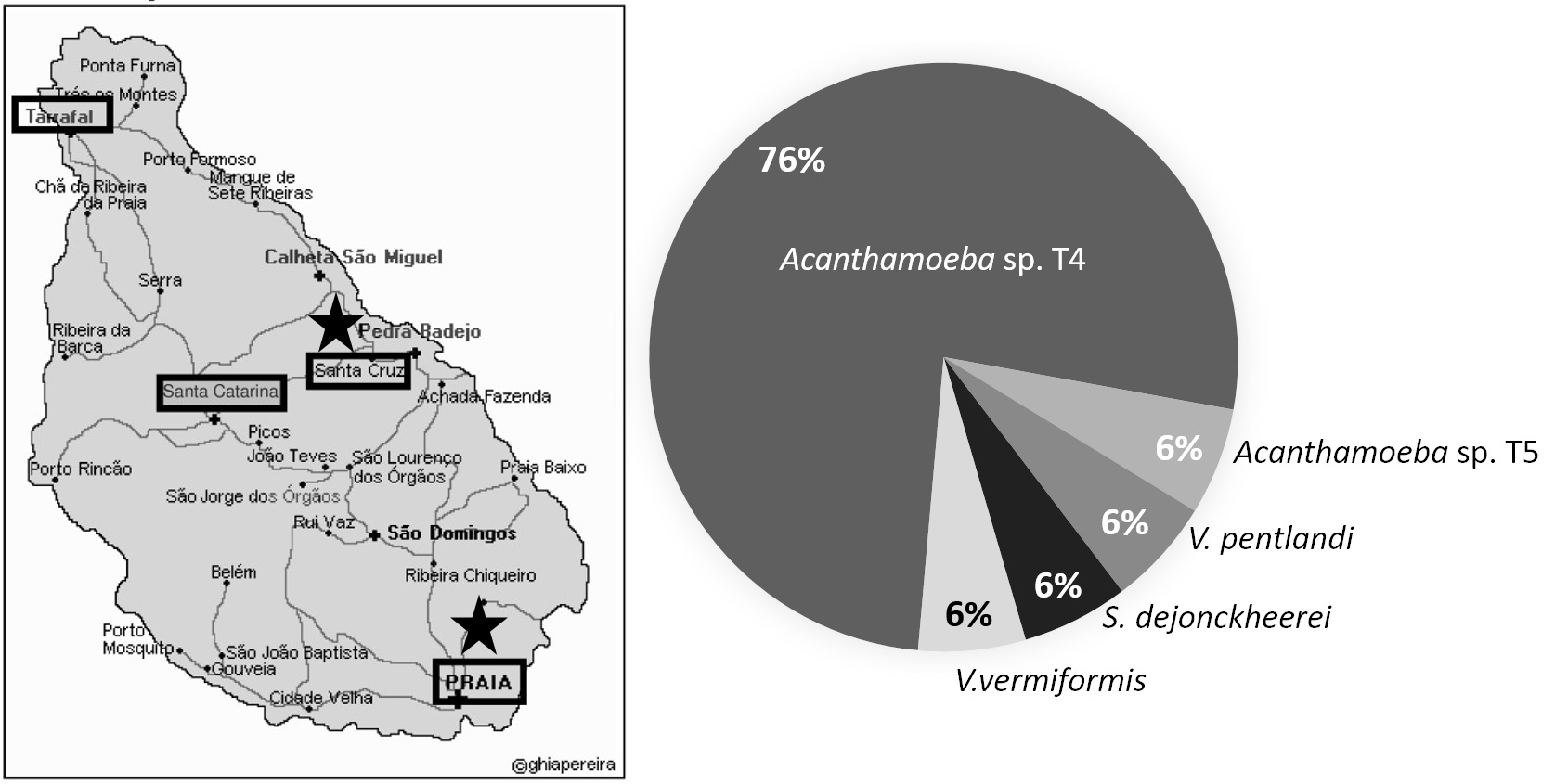
2.1. Sample Sites and Culture of FLA
2.2. DNA Extraction
2.3. PCR and Molecular Characterization of Isolates
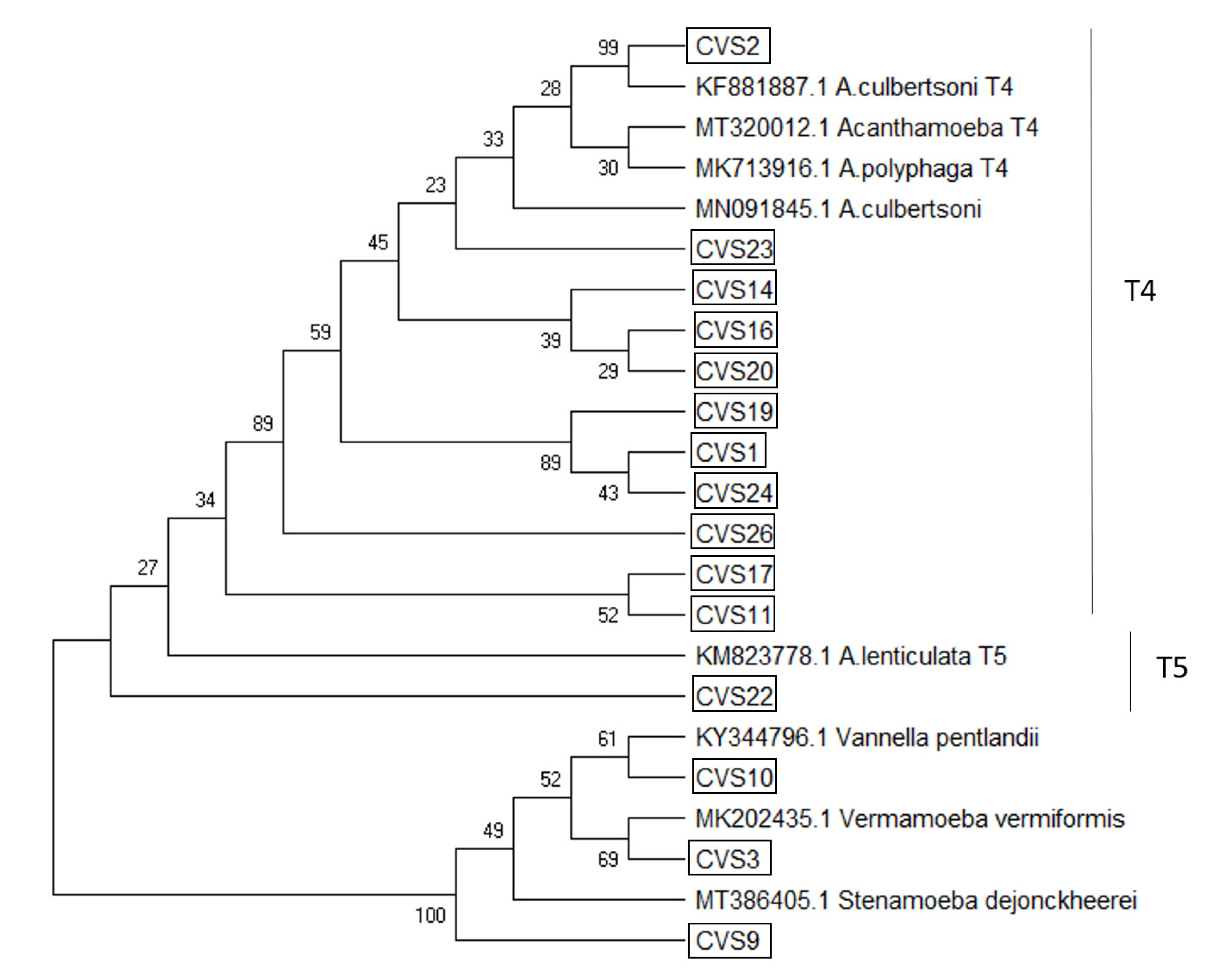
2.4. Phylogenetic Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schuster, F.L.; Visvesvara, G.S. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int. J. Parasitol. 2004, 34, 1001–1027. [Google Scholar] [CrossRef]
- Guimaraes, A.J.; Gomes, K.X.; Cortines, J.R.; Peralta, J.M.; Peralta, R.H. Acanthamoeba spp. as a universal host for pathogenic microorganisms: One bridge from environment to host virulence. Microbiol. Res. 2016, 193, 30–38. [Google Scholar] [CrossRef]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasites Vectors 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Scheid, P.L.; Lâm, T.T.; Sinsch, U.; Balczun, C. Vermamoeba vermiformis as etiological agent of a painful ulcer close to the eye. Parasitol. Res. 2019, 118, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Cateau, E.; Delafont, V.; Hechard, Y.; Rodier, M.H. Free-living amoebae: What part de they play in healthcare-associated infections? J. Hosp. Infec. 2014, 87, 131–140. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Geisen, S.; Wubs, E.R.J.; Song, C.; de Boer, W.; Garbeva, P. The prey’s scent—Volatile organic compound mediated interactions between soil bacteria and their protist predators. ISME J. 2017, 11, 817–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mella, C.; Medina, G.; Flores-Martin, S.; Toledo, Z.; Simaluiza, R.J.; Pérez-Pérez, G.; Fernández, H. Interaction between zoonotic bacteria and free living amoebas: A new angle of an epidemiological polyhedron of public health importance? Arch. Med. Vet. 2016, 48, 1–10. [Google Scholar] [CrossRef]
- Neto, C.; Costa, J.C.; Figueiredo, A.; Capelo, J.; Gomes, I.; Vitória, S.; Semedo, J.M.; Lopes, A.; Dinis, H.; Correia, E.; et al. The Role of Climate and Topography in Shaping the Diversity of Plant Communities in Cabo Verde Islands. Diversity 2020, 12, 80. [Google Scholar] [CrossRef] [Green Version]
- Freire, S.M.; Relvas, H.; Lopes, M. Impact of traffic emissions on air quality in Cabo Verde. Environ. Monit. Assess. 2020, 192, 726. [Google Scholar] [CrossRef]
- Ferreira Couto, C. Incerteza, Adaptabilidade e Inovação na Sociedade Rural da Ilha de Santiago de Cabo Verde; Fundação Galouste Gulbenkian: Lisbon, Portugal, 2010; Volume XXV, 461p. [Google Scholar]
- Berlitz Pocket Guide Cape Verde, 3rd revised ed.; Berlitz Travel, 2018; ISBN 978-1785730627.
- Page, F.C. Mitosis and pseudopod formation in Vexillifera bacillipedes n. sp.; A majorellid amoeba. Trans. Am. Microsc. Soc. 1969, 88, 394–400. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Monteverde-Miranda, C.A.; Jiménez, C.; Tejedor, M.L.; Valladares, B.; Ortega-Rivas, A. Evaluation of Acanthamoeba isolates from environmental sources in Tenerife, Canary Islands, Spain. Ann. Agric. Environ. Med. 2005, 12, 233–236. [Google Scholar] [PubMed]
- Lorenzo-Morales, J.; Ortega-Rivas, A.; Foronda, P.; Martínez, E.; Valladares, B. Isolation and identification of pathogenic Acanthamoeba strains in Tenerife, Canary Islands, Spain from water sources. Parasitol. Res. 2005, 95, 273–277. [Google Scholar] [CrossRef]
- Reyes-Batlle, M.; Niyyati, M.; Martín-Navarro, C.M.; López-Arencibia, A.; Valladares, B.; Martínez-Carretero, E.; Piñero, J.E.; Lorenzo-Morales, J. Unusual Vermamoeba vermiformis strain isolated from snow in Mount Teide, Tenerife, Canary Islands, Spain. Nov. Biomed. 2015, 3, 189–192. [Google Scholar] [CrossRef]
- Alves, J.; Roque, A.L.; Cravo, P.; Valdez, T.; Jelinek, T.; Rosário, V.E.; Arez, A.P. Epidemiological characterization of Plasmodium falciparum in the Republic of Cabo Verde: Implications for potential large-scale re-emergence of malaria. Malar. J. 2006, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Batlle, M.; Zamora-Herrera, J.; Vargas-Mesa, A.; Valerón-Tejera, M.A.; Wagner, C.; Martín-Navarro, C.M.; López-Arencibia, A.; Sifaoui, I.; Martínez-Carretero, E.; Valladares, B.; et al. Acanthamoeba genotypes T2, T4, and T11 in soil sources from El Hierro island, Canary Islands, Spain. Parasitol. Res. 2016, 115, 2953–2956. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, N.; Schild, M.; Panaiotov, S.; Kurdova-Mintcheva, R.; Gottstein, B.; Walochnik, J.; Aspöck, H.; Lucas, M.S.; Müller, N. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasit. Res. 2004, 92, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.Y.; Ji, D.R.; Hsia, K.T.; Hung, C.C.; Sheng, W.H.; Hsu, B.M.; Chen, J.S.; Wu, M.H.; Lai, C.H.; Ji, D.D. Isolation and identification of Acanthamoeba species related to amoebic encephalitis and nonpathogenic free-living amoeba species from the rice field. J. Appl. Microbiol. 2010, 109, 1422–1429. [Google Scholar] [CrossRef]
- Schroeder, J.M.; Booton, G.C.; Hay, J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of Acanthamoebae from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39, 1903–1911. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; López-Darias, M.; Martínez-Carretero, E.; Valladares, B. Isolation of potentially pathogenic strains of Acanthamoeba in wild squirrels from the Canary Islands and Morocco. Exp. Parasitol. 2007, 117, 74–79. [Google Scholar] [CrossRef] [PubMed]
- De Jonckheere, J.F. Isolation and Molecular Identification of Vahlkampfiid Amoebae from an Island (Tenerife, Spain). Acta Protozool. 2006, 45, 91–96. [Google Scholar]
- Dyková, I.; Lorenzo-Morales, J.; Kostka, M.; Valladares, B.; Pecková, H. Neoparamoeba branchiphila infections in moribund sea urchins Diadema aff. antillarum in Tenerife, Canary Islands, Spain. Dis. Aquat. Organ. 2011, 95, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Batlle, M.; Todd, C.D.; Martín-Navarro, C.M.; López-Arencibia, A.; Cabello-Vilchez, A.M.; González, A.C.; Córdoba-Lanús, E.; Lindo, J.F.; Valladares, B.; Piñero, J.E.; et al. Isolation and characterization of Acanthamoeba strains from soil samples in Gran Canaria, Canary Islands, Spain. Parasitol. Res. 2014, 113, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Batlle, M.; Wagner, C.; Zamora-Herrera, J.; Vargas-Mesa, A.; Sifaoui, I.; González, A.C.; López-Arencibia, A.; Valladares, B.; Martínez-Carretero, E.; Piñero, J.E.; et al. Isolation and Molecular Identification of Vermamoeba vermiformis Strains from Soil Sources in El Hierro Island, Canary Islands, Spain. Curr. Microbiol. 2016, 73, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Batlle, M.; Wagner, C.; Zamora-Herrera, J.; Vargas-Mesa, A.; Sifaoui, I.; González, A.C.; López-Arencibia, A.; Valladares, B.; Martínez-Carretero, E.; Piñero, J.E.; et al. Isolation of thermotolerant Vermamoeba vermiformis strains from water sources in Lanzarote Island, Canary Islands, Spain. Acta Parasitol. 2016, 61, 650–653. [Google Scholar] [CrossRef]
- Reyes-Batlle, M.; Hernández-Piñero, I.; Rizo-Liendo, A.; López-Arencibia, A.; Sifaoui, I.; Bethencourt-Estrella, C.J.; Chiboub, O.; Valladares, B.; Piñero, J.E.; Lorenzo-Morales, J. Isolation and molecular identification of free-living amoebae from dishcloths in Tenerife, Canary Islands, Spain. Parasitol. Res. 2019, 118, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Batlle, M.; Rizo-Liendo, A.; Viera-Santana, R.A.; Afonso-Morales, S.; López-Arencibia, A.; Sifaoui, I.; Chiboub, O.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Rodríguez-Expósito, R.L.; et al. Isolation and molecular identification of Naegleria australiensis in irrigation water of Fuerteventura Island, Spain. Acta Parasitol. 2019, 64, 331–335. [Google Scholar] [CrossRef]
- Reyes-Batlle, M.; Díaz, F.J.; Sifaoui, I.; Rodríguez-Expósito, R.; Rizo-Liendo, A.; Piñero, J.E.; Lorenzo-Morales, J. Free living amoebae isolation in irrigation waters and soils of an insular arid agroecosystem. Sci. Total Environ. 2021, 753, 141833. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Valladares, M.; Sancho, J.; Reyes-Batlle, M.; Martin-Navarro, C.M.; Lopez-Arencibia, A.; Gonzalez, A.C.; Lopez-Medina, L.; Piñero, J.E.; Martinez-Carretero, E.; et al. First report of a case of prostatitis due to Acanthamoeba in a dog. Acta Protozool. 2013, 52, 325–329. [Google Scholar] [CrossRef]
- Valladares, M.; Reyes-Batlle, M.; Mora-Peces, I.; Martín-Navarro, C.M.; López-Arencibia, A.; Dorta-Gorrín, A.; Comyn-Afonso, E.; Martínez-Carretero, E.; Maciver, S.K.; Piñero, J.E.; et al. A multisystemic Acanthamoeba infection in a dog in Tenerife, Canary Islands, Spain. Vet. Parasitol. 2014, 205, 707–711. [Google Scholar] [CrossRef]
- Valladares, M.; Reyes-Batlle, M.; Martín-Navarro, C.M.; López-Arencibia, A.; Dorta-Gorrín, A.; Wagner, C.; Martínez-Carretero, E.; Piñero, J.E.; Valladares, B.; Lorenzo-Morales, J. Molecular characterization of Acanthamoeba strains isolated from domestic dogs in Tenerife, Canary Islands, Spain. Arch. Microbiol. 2015, 197, 639–643. [Google Scholar] [CrossRef]
- Petters, S.; Groß, V.; Söllinger, A.; Pichler, M.; Reinhard, A.; Bengtsson, M.M.; Urich, T. The soil microbial food web revisited: Predatory myxobacteria as keystone taxa? ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, D.; Walochnik, J.; Kohsler, M.; Rott, M.B. Acanthamoeba misidentifcation and multiple labels: Redefning genotypes T16, T19, and T20 and proposal for Acanthamoeba micheli sp. nov. (genotype T19). Parasitol. Res. 2015, 114, 2481–2490. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, P.A.; Booton, G.C. Species, Sequence Types and Alleles: Dissecting Genetic Variation in Acanthamoeba. Pathogens 2020, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; Vanzzini-Zago, V.; Hernandez-Martinez, D.; Gonzalez-Robles, A.; Salazar-Villatoro, L.; Ramirez-Flores, E.; Oregon-Miranda, E.; Lorenzo-Morales, J.; Martinez-Palomo, A. Acanthamoeba genotypes T3 and T4 as causative agents of amoebic keratitis in Mexico. Parasitol. Res. 2016, 115, 873–878. [Google Scholar] [CrossRef]
- Castro-Artavia, E.; Retana-Moreira, L.; Lorenzo-Morales, J.; Abrahams-Sandi, E. Potentially pathogenic Acanthamoeba genotype T4 isolated from dental units and emergency combination showers. Memórias Inst. Oswaldo Cruz. 2017, 112, 817–821. [Google Scholar] [CrossRef] [Green Version]
- Geisen, S.; Koller, R.; Hünninghaus, M.; Dumack, K.; Urich, T.; Bonkowski, M. The soil food web revisited: Diverse and widespread mycophagous soil protists. Soil Biol. Biochem. 2016, 94, 10–18. [Google Scholar] [CrossRef]
- Twafeek, G.M.; Biahara, S.A.H.; Sarhan, R.M.; Taher, E.E.; Khayyal, A.E. Genotypic, physiological, and biochemical characterization of potentially pathogenic Acanthamoeba isolated from environment in Cairo, Egypt. Parasitol. Res. 2016, 115, 1871–1881. [Google Scholar] [CrossRef]
- Khan, N.A. The pathogenesis of Acanthamoeba infections: Current status and future implications. Microb. Pathog. 2003, 34, 277–285. [Google Scholar] [CrossRef]
- Leede, D.R.; Lovieno, A.; Miller, D.; Mandal, N.; Diaz, M.; Fell, J.; Fini, M.E.; Alfonso, E.C. Molecular identification of T4 and T5 genotypes in isolates from Acanthamoeba keratitis patients. J. Clin. Microbiol. 2009, 47, 1458–1462. [Google Scholar] [CrossRef] [Green Version]
- Barete, S.; Combes, A.; De Jomcheere, J.; Datry, A.; Varnous, S.; Martinez, V.; Ptacek, S.G.; Caumes, E.; Capron, F.; Frances, C.; et al. Fatal disseminated Acanthamoeba lenticulata acanthamebiasis in a heart transplant patient. Emerg. Infect. Dis. 2007, 13, 736–738. [Google Scholar] [CrossRef]
- Centeno, M.; Rivera, F.; Cerva, L.; Tsutsumi, V.; Gallegos, E.; Calderón, A.; Ortiz, R.; Bonilla, P.; Ramírez, E.; Suárez, G. Hartmannella vermiformis isolated from the cerebrospinal fluid of a young male patient with meningoencephalitis and bronchopneumonia. Arch. Med. Res. 1996, 27, 579–586. [Google Scholar]
- Delafont, V.; Rodier, M.H.; Maisonneuve, E.; Cateau, E. Vermamoeba vermiformis: A Free-Living Amoeba of Interest. Microbial. Ecol. 2018, 76, 991–1001. [Google Scholar] [CrossRef]
- Loret, J.F.; Jousset, M.; Robert, S.; Saucedo, G.; Ribas, F.; Thomas, V.; Greub, G. Amoebae-resisting bacteria in drinking water: Risk assessment and management. Water Sci. Tech. 2008, 58, 571–577. [Google Scholar] [CrossRef]
- Siddiqui, R.; Makhlouf, Z.; Khan, N.A. The Increasing Importance of Vermamoeba vermiformis. J. Eukaryot. Microbiol. 2021, e12857. [Google Scholar] [CrossRef]
- Abedkhojasteh, H.; Niyyati, M.; Rahimi, F.; Heidari, M.; Farnia, S.; Rezaeian, M. First Report of Hartmannella keratitis in a Cosmetic Soft Contact Lens Wearer in Iran. Iran. J. Parasitol. 2013, 8, 481–485. [Google Scholar] [PubMed]
- Scheid, L.P. Vermamoeba vermiformis-A free-living Amoeba with public health and environmental health significance. Open Parasitol. J. 2019, 7. [Google Scholar] [CrossRef]
- Thomas, V.; Loret, J.-F.; Jousset, M.; Greub, G. Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant. Environ. Microbiol. 2008, 10, 2728–2745. [Google Scholar] [CrossRef]
- Poitelon, J.-B.; Joyeux, M.; Welte, B.; Duguet, J.-P.; Peplies, J.; DuBow, M.S. Identification and phylogeny of eukaryotic 18S rDNA phylotypes detected in chlorinated finished drinking water samples from three Parisian surface water treatment plants. Lett. Appl. Microbiol. 2009, 49, 589–595. [Google Scholar] [CrossRef]
- Rivera, F.; Ramírez, E.; Bonilla, P.; Calderon, A.; Gallegos, E.; Rodríguez, S.; Ortiz, R.; Zaldívar, B.; Ramírez, P.; Duran, A. Pathogenic and free living amoebae isolated from swimming pools and physiotherapy tubs in Mexico. Environ. Res. 1993, 62, 43–52. [Google Scholar] [CrossRef]
- Chavatte, N.; Lambrecht, E.; Van Damme, I.; Sabbe, K.; Houf, K. Abundance, diversity and community composition of free-living protozoa on vegetable sprouts. Food Microbiol. 2016, 55, 55–63. [Google Scholar] [CrossRef]
- Maciver, S.K.; De Obeso Fernandez Del Valle, A.; Koutsogiannis, Z. Vannella pentlandii n. sp.; (Amoebozoa, Discosea, Vannellida) a small, cyst-forming soil amoeba. Exp. Parasitol. 2017, 183, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Schulz, F.; Tyml, T.; Pizzetti, I.; Dykova, I.; Fazi, S.; Kostka, M.; Horn, M. Marine amoebae with cytoplasmic and perinuclear symbionts deeply branching in the Gammaproteobacteria. Sci. Rep. 2015, 5, 13381. Available online: https://pubmed.ncbi.nlm.nih.gov/26303516/ (accessed on 7 July 2021). [CrossRef] [Green Version]
- Kuroki, T.; Yagita, K.; Yabuuchi, E.; Agata, K.; Ishima, T.; Katsube, Y.; Endo, T. Kansenshogaku zasshi. JJID 1998, 72, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Scheid, P. Mechanism of intrusion of a microspordian-like organism into the nucleus of host amoebae (Vannella sp.) isolated from a keratitis patient. Parasitol. Res. 2007, 101, 1097–1102. [Google Scholar] [CrossRef]
- Smirnov, A.V.; Nassonova, E.S.; Chao, E.; Cavalier-Smith, T. Phylogeny, Evolution, and Taxonomy of Vannellid Amoebae. Protist 2007, 158, 295–324. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianinazzi, C.; Schild, M.; Zumkehr, B.; Wuthrich, F.; Nuesch, I.; Ryter, R.; Schurch, N.; Gottstein, B.; Muller, N. Screening of Swiss hot spring resorts for potentially pathogenic free-living amoebae. Exp. Parasitol. 2010, 126, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Lares-Jiménez, L.F.; Borquez-Román, M.A.; Lares-García, C.; Otero-Ruiz, A.; Gonzalez-Galaviz, J.R.; Ibarra-Gámez, J.C.; Lares-Villa, F. Potentially pathogenic genera of free-living amoebae coexisting in a thermal spring. Exp. Parasitol. 2018, 195, 54–58. [Google Scholar] [CrossRef]
- Borquez-Román, M.A.; Lares-Jiménez, L.F.; Rodriguez-Anaya, L.Z.; Gonzalez-Galaviz, J.R.; Fuerst, P.A.; Ibarra-Gámez, J.C.; Casillas-Hernández, R.; Lares-Villa, F. Stenamoeba dejonckheerei sp. nov.; a Free-Living Amoeba Isolated from a Thermal Spring. Pathogens 2020, 9, 586. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Sample Code | Locality | Soil Type | NNA | PCR | Isolate Specie | Seq Length (bp) | Identity (%) * | Sequence ID BLASTn |
|---|---|---|---|---|---|---|---|---|
| CVS1 | Varzea, Praia | Garden | + | + | Acanthamoeba sp. T4 | 412 | >99% | MN153014.1 |
| CVS2 | Santa Cruz | Garden | + | + | Acanthamoeba sp. T4 | 368 | >92% | MN239986.1 |
| CVS3 | Camboinha, Praia | Beach | + | + | Vermamoeba vermiformis | 422 | >99% | MK202435.1 |
| CVS9 | Praia | Garden | + | + | Stenamoeba dejonckheerei | 200 | >96% | MT386405.1 |
| CVS10 | Palmarejo, Praia | Garden | + | + | Vannella pentlandi | 189 | >95% | KY344796.1 |
| CVS11 | Palmarejo, Praia | Garden | + | + | Acanthamoeba sp. T4 | 276 | >95% | MT613705.1 |
| CVS14 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 414 | >95% | KU936118.1 |
| CVS16 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 412 | 100% | LC373015.1 |
| CVS17 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 246 | >95% | MK390856.1 |
| CVS19 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 403 | >99% | JQ418500.1 |
| CVS20 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 406 | >99% | MN700306.1 |
| CVS21 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 437 | >98% | GU808328.1 |
| CVS22 | Santa Cruz | Crop | + | + | Acanthamoeba lenticulata T5 | 363 | >99% | KM189378.1 |
| CVS23 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 427 | >98% | MT645313.1 |
| CVS24 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 406 | 100% | JF702882.1 |
| CVS25 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 408 | >95% | KY827390.1 |
| CVS26 | Santa Cruz | Crop | + | + | Acanthamoeba sp. T4 | 315 | 100% | MT613705.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa-Ramos, D.; Reyes-Batlle, M.; Bellini, N.K.; Rodríguez-Expósito, R.L.; Piñero, J.E.; Lorenzo-Morales, J. Free-Living Amoebae in Soil Samples from Santiago Island, Cape Verde. Microorganisms 2021, 9, 1460. https://doi.org/10.3390/microorganisms9071460
Sousa-Ramos D, Reyes-Batlle M, Bellini NK, Rodríguez-Expósito RL, Piñero JE, Lorenzo-Morales J. Free-Living Amoebae in Soil Samples from Santiago Island, Cape Verde. Microorganisms. 2021; 9(7):1460. https://doi.org/10.3390/microorganisms9071460
Chicago/Turabian StyleSousa-Ramos, Djeniffer, María Reyes-Batlle, Natália K. Bellini, Rubén L. Rodríguez-Expósito, José E. Piñero, and Jacob Lorenzo-Morales. 2021. "Free-Living Amoebae in Soil Samples from Santiago Island, Cape Verde" Microorganisms 9, no. 7: 1460. https://doi.org/10.3390/microorganisms9071460
APA StyleSousa-Ramos, D., Reyes-Batlle, M., Bellini, N. K., Rodríguez-Expósito, R. L., Piñero, J. E., & Lorenzo-Morales, J. (2021). Free-Living Amoebae in Soil Samples from Santiago Island, Cape Verde. Microorganisms, 9(7), 1460. https://doi.org/10.3390/microorganisms9071460