
Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review

 ,
,
Abstract
:1. Introduction
2. Environment Relation
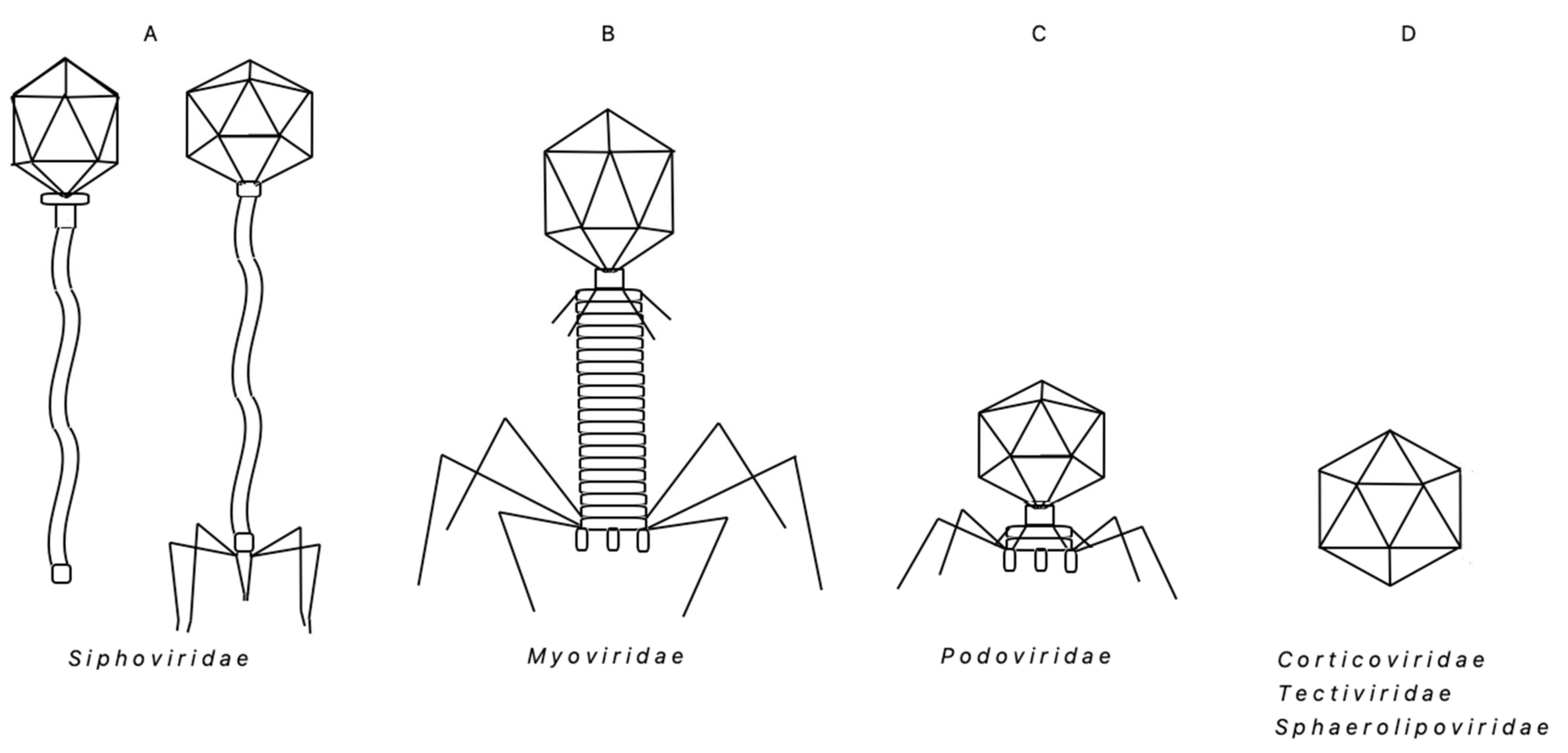
3. ‘Bacillus Group’ Bacteriophages—Chronological Review
3.1. ‘Thermophilic Lytic Principle’ (Family/Genus Not Determined (ND), Host Putative G. stearothermophilus)
3.2. ‘Thermophilic Bacteriophage’ (1) (Family/Genus ND, Host Thermophilic Bacillus sp.)
3.3. ’Thermophilic Bacteriophage’ (2) (Family/Genus ND, Host ‘Thermophilic Bacterium No. 10′)
3.4. ‘Thermophilic Bacteriophage’ (3) (Family/Genus ND, Host Thermophilic Bacillus sp.)
3.5. TP-84 (Siphoviridae Family, Tp84virus Genus, Host B. stearothermophilus)
3.6. φμ-4 (Family/Genus Not Determined, Host B. stearothermophilus)
3.7. TP-1 (Putative Siphoviridae Family, Host B. stearothermophilus)
3.8. ST1 (Siphoviridae Family, Genus ND, Host B. stearothermophilus)
3.9. Tφ3 (Tphi3) (Siphoviridae Family, Genus ND, Host G. stearothermophilus)
3.10. GH5, GH8 (Family/Genus ND, Host B. stearothermophilus)
3.11. PhB1 (Siphoviridae Family, Genus ND, Host Thermophilic Bacillus sp.)
3.12. D5–D8 (Family/Genus ND, Host B. stearothermophilus)
3.13. φNS11 (PhiNS11) (Putative Sphaerolipoviridae, Tectiviridae or Corticoviridae Family, Host Acidophilic-Thermophilic B. acidocaldarius)
3.14. JS001-JS027 Series (Section 3.14, Section 3.15, Section 3.16, Section 3.17, Section 3.18, Section 3.19, Section 3.20, Section 3.21, Section 3.22, Section 3.23, Section 3.24, Section 3.25, Section 3.26, Section 3.27, Section 3.28, Section 3.29, Section 3.30, Section 3.31, Section 3.32, Section 3.33, Section 3.34, Section 3.35, Section 3.36 and Section 3.37 Below)
3.15. JS001 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.16. JS004 (Putative Podoviridae Family, Hosts Thermophilic Bacillus sp.)
3.17. JS005 (Family/Genu ND, Hosts Thermophilic Bacillus sp.)
3.18. JS006 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.19. JS007 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.20. JS008 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.21. JS009 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.22. JS010 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.23. JS011 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.24. JS012 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.25. JS013 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.26. JS014 (Putative Siphoviridae, Hosts Thermophilic Bacillus sp.)
3.27. JS015 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
3.28. JS017 (Putative Myoviridae Family, Hosts Thermophilic Bacillus sp.)
3.29. JS018 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.30. JS019 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.31. JS020 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
3.32. JS021 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
3.33. JS022 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.34. JS023 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
3.35. JS024 (Putative Siphoviridae or Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.36. JS025 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.37. JS026 (Putative Siphoviridae or Myoviridae Family, Hosts Thermophilic B. caldotenax)
3.38. JS027 (Putative Podoviridae Family, Hosts Thermophilic Bacillus sp.)
3.39. W1 (BVW1) (Siphoviridae Family, Genus ND, Host Thermophilic Bacillus sp.)
3.40. GVE1 (E1) (Siphoviridae Family, Genu NDs, Host Geobacillus sp.)
3.41. GVE2 (E2) (Siphoviridae Family, Genus ND, Hosts Geobacillus sp.)
3.42. GBSV1 (Myoviridae Family, Svunavirus Genus, Hosts Geobacillus sp.)
3.43. BV1 (Myoviridae, Family, Svunavirus Genus, Hosts Geobacillus sp.)
3.44. D6E (Myoviridae Family, Genus ND, Host Geobacillus sp.)
3.45. ϕOH2 (phiOH2) (Family/Genus ND, Host G. kaustophilus)
3.46. GBK2 (Siphoviridae Family, Genus ND, Host G. kaustophilus)
3.47. GVE3 (Siphoviridae Family, Genus ND, Host G. thermoglucosidasius)
3.48. AP45 (Siphoviridae Family, Genus ND, Host Aeribacillus sp.)
3.49. Series of 5 Bacteriophages from Haloalkaline Lake Elmenteita of Kenyan Rift Valley (Section 3.49, Section 3.50, Section 3.51, Section 3.52, Section 3.53 and Section 3.54 Below)
3.50. vB_BpsS-36 (Family/Genus ND, Host B. pseudalcaliphilus)
3.51. vB_BpsM-61 (Family/Genus ND, Host B. pseudofirmus)
3.52. vB_BboS-125 (Family/Genus ND, Host B. bogoriensis)
3.53. vB_BcoS-136 (Family/Genus ND, Host B. cohnii)
3.54. vB_BpsS-140 (Family/Genus ND, Host B. pseudalcaliphilus)
4. Scientific, Biotechnology, Environmental and Medical Potential—A Short Note
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shapiro, J.W.; Putonti, C. Gene co-occurrence networks reflect bacteriophage ecology and evolution. mBio 2018, 9, e01870-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowron, P.M.; Kropinski, A.M.; Zebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; et al. Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84virus genus. PLoS ONE 2018, 13, e0195449. [Google Scholar] [CrossRef]
- Zablocki, O.; van Zyl, L.; Trindade, M. Biogeography and taxonomic overview of terrestrial hot spring thermophilic bacteriophages. Extremophiles 2018, 22, 827–837. [Google Scholar] [CrossRef]
- Wisotzkey, J.D.; Jurtshuk, P., Jr.; Fox, G.E.; Deinhard, G.; Poralla, K. Comparative sequence analyses on the 16S rRNA (rDNA) of Bacillus acidocaldarius, Bacillus acidoterrestris, and Bacillus cycloheptanicus and proposal for creation of a new genus, Alicyclobacillus gen. nov. Int. J. Syst. Bacteriol. 1992, 42, 263–269. [Google Scholar] [CrossRef]
- Dufresne, S.; Bousquet, J.; Boissinot, M.; Guay, R. Sulfobacillus disulfidooxidans sp. nov., a new acidophilic, disulfide-oxidizing, gram-positive, spore-forming bacterium. Int. J. Syst. Bacteriol. 1996, 46, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyndrickx, M.; Lebbe, L.; Vancanneyt, M.; Kersters, K.; De Vos, P.; Logan, N.A.; Forsyth, G.; Nazli, S.; Ali, N.; Berkeley, R.C.W. A polyphasic reassessment of the genus Aneurinibacillus, reclassification of Bacillus thermoaerophilus (Meier-Stauffer et al. 1996) as Aneurinibacillus thermoaeropilus comb. nov., and emended descriptions of A. aneurinilyticus corrig., A migulanus, and A. thermoaerophilus. Int. J. Syst. Bacteriol. 1997, 47, 808–817. [Google Scholar]
- Touzel, J.P.; O’Donohue, M.; Debeire, P.; Samain, E.; Breton, C. Thermobacillus xylanilyticus gen. nov., sp. nov., a new aerobic thermophilic xylan-degrading bacterium isolated from farm soil. Int. J. Syst. Evol. Microbiol. 2000, 50, 315–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainey, F.A.; Fritze, D.; Stackebrandt, E. The phylogenetic diversity of thermophilic members of the genus Bacillus as revealed by 16S rDNA analysis. FEMS Microbiol. Lett. 1994, 115, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic study of aerobic thermophilic bacilli: Descriptions of Geobacillus subterraneus gen. nov., sp. nov. and Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermoglucosidasius and Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. thermocatenulatus, G. thermoleovorans, G. kaustophilus, G. thermoglucosidasius and G. thermodenitrificans. Int. J. Syst. Evol. Microbiol. 2001, 51, 433–446. [Google Scholar] [CrossRef] [Green Version]
- McMullan, G.; Christie, J.M.; Rahman, T.J.; Banat, I.M.; Ternan, N.G.; Marchant, R. Habitat, applications and genomics of the aerobic, thermophilic genus Geobacillus. Biochem. Soc. Trans. 2004, 32, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Yoshinari, A.; Aoyagi, T.; Hirota, Y.; Iwamoto, K.; Sato, T. Regulated DNA rearrangement during sporulation in Bacillus weihenstephanensis KBAB4. Mol. Microbiol. 2013, 90, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Roger, W.; Hendrix, R.W. Bacteriophages: Evolution of the mMajority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar]
- Reysenbach, A.-L.; Shock, E. Merging genomes with geochemistry in hydrothermal ecosystems. Science 2002, 296, 1077–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, X. Genome analysis of deep-sea thermophilic bacteriophage D6E. Appl. Environ. Microbiol. 2010, 76, 7861–7866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, R.J.; Riley, P.W.; White, D. Thermophilic Bacilli; Kristjansson, J.K., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 19–50. [Google Scholar]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef]
- Blanc, M.; Marilley, L.; Beffa, T.; Aragno, M. Rapid identification of heterotrophic, thermophilic, spore-form-ing bacteria isolated from hot composts. Int. J. Syst. Bacteriol. 1997, 47, 1246–1248. [Google Scholar] [CrossRef] [Green Version]
- Marks, T.J.; Hamilton, P.T. Characterization of a thermophilic bacteriophage of Geobacillus kaustophilus. Arch. Virol. 2014, 159, 2771–2775. [Google Scholar] [CrossRef]
- Bell, E.M. Life at Extremes: Environments, Organisms, and Strategies for Survival; CABI: Wallingford/Oxfordshire, UK, 2012. [Google Scholar]
- Pentecost, A. High temperature ecosystems and their chemical interactions with their environment. In Ciba Foundation Symposium 202-Evolution of Hydrothermal Ecosystems on Earth (And Mars?): Ciba Foundation Symposium 202; Bock, G.R., Goode, J.A., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 1996; pp. 99–111. [Google Scholar] [CrossRef]
- Sharp, R.J.; Ahmad, S.I.; Munster, A.; Dowsett, B.; Atkinson, T. The isolation and characterization of bacterio-bacteriophages infecting obligately thermophilic strains of Bacillus. J. Gen. Microbiol. 1986, 132, 1709–1722. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wu, S.; Song, Q.; Zhang, X.; Xie, L. Two novel bacteriophages of thermophilic bacteria isolated from deep-sea hydrothermal fields. Curr. Microbiol. 2006, 53, 163–166. [Google Scholar] [CrossRef]
- Nagayoshi, Y.; Kumagae, K.; Mori, K.; Tashiro, K.; Nakamura, A.; Fujino, Y.; Hiromasa, Y.; Iwamoto, T.; Kuhara, S.; Ohshima, T.; et al. Physiological properties and genome structure of the hyperthermophilic filamentous bacteriophage φOH3 which infects Thermus thermophilus HB8. Front. Microbiol. 2016, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zhang, X. Deep-sea thermophilic Geobacillus bacteriophage GVE2 transcriptional profile and proteomic characterization of virions. Appl. Microbiol. Biotechnol. 2008, 80, 697–707. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, F.; Wu, S.; Xu, Y.; Zhang, X. Genomic and proteomic characterization of a thermophilic Geobacillus bacteriophage GBSV1. Res. Microbiol. 2009, 16, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Mori, K.; Martono, H.; Nagayoshi, Y.; Fujino, Y.; Tashiro, K.; Kuhara, S.; Ohshima, T. Draft genome sequence of Geobacillus kaustophilus GBlys, a lysogenic strain with bacteriophage ϕOH2. Genome Announc. 2013, 1, e00634-13. [Google Scholar] [CrossRef] [Green Version]
- Morozova, V.; Bokovaya, O.; Kozlova, Y.; Kurilshikov, A.; Babkin, I.; Tupikin, A.; Bondar, A.; Ryabchikova, E.; Brayanskaya, A.; Peltek, S.; et al. A novel thermophilic Aeribacillus bacteriophage AP45 isolated from the Valley of Geysers, Kamchatka: Genome analysis suggests the existence of a new genus within the Siphoviridae family. Extremophiles 2019, 23, 599–612. [Google Scholar] [CrossRef]
- Wei, D.; Zhang, X. Identification and characterization of a single-stranded DNA-binding protein from thermophilic bacteriophage GVE2. Virus Genes 2008, 36, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Brister, J.R.; Ako-Adjei, D.; Bao, Y.; Blinkova, O. NCBI viral genomes resource. Nucleic Acids Res. 2015, 43, D571–D577. [Google Scholar] [CrossRef] [Green Version]
- Saunders, G.F.; Campbell, L.L.; Postgate, J.R. Abstract. In Proceedings of the 148th National Meeting of the American Chemical Society, Chicago, IL, USA, 30 August–4 September 1964. [Google Scholar]
- Koser, S.A. Action of the bacteriophage on a thermophilic Bacillus. Exp. Biol. Med. 1926, 24, 109–111. [Google Scholar] [CrossRef]
- Adant, M. Les bacteriophages des microbes thermophiles. Compt. Rend. Soc. Biol. 1928, 99, 1244–1245. [Google Scholar]
- White, R.; Georgi, C.E.; Militzer, W. Heat studies on a thermophilic bacteriophage. Proc. Soc. Exp. Biol. Med. 1954, 85, 137–139. [Google Scholar] [CrossRef]
- Marsh, C.L.; Larsen, D.H. Characterization of some thermophilic bacteria from the Hot Springs of Yellowstone National Park. J. Bacteriol. 1953, 65, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.; Georgi, C.E.; Militzer, W.E. Characteristics of a thermophilic bacteriophage. Proc. Soc. Exp. Biol. Med. 1955, 88, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Hirano, J.; Mutoh, Y.; Kitamura, M.; Asai, S.; Nakajima, I.; Takamiya, A. Effect of change of temperature upon the viability of a thermophilic bacterium. J. Gen. Appl. Microbiol. 1958, 4, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Onodera, N. Electron-Microscopy 1959, 7, 2–3, 19. (In Japanese)
- Onodera, N. On some characteristics of a newly isolated thermophilic bacteriophage and consideration of its thermostability. J. Electron. Microsc. (Tokyo) 1961, 10, 91–102. [Google Scholar] [CrossRef]
- Saunders, G.F.; Campbell, L.L. Characterization of a thermophilic bacteriophage for Bacillus stearothermophilus. J. Bacteriol. 1966, 91, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Zeigler, D.R. The genus Geobacillus. In Bacillus Genetic Stock Center, Catalog of Strains, 7th ed.; Department of Biochemistry, The Ohio State University: Columbus, OH, USA, 2001; Volume 3, pp. 1–25. [Google Scholar]
- Epstein, I.; Campbell, L. Production and purification of the thermophilic bacteriophage TP-84. Appl. Microbiol. 1975, 29, 219–223. [Google Scholar] [CrossRef]
- Shafiai, F.; Thompson, T.L. Isolation and preliminary characterization of bacteriophage φμ-4. J. Bacteriol. 1964, 87, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.; Campbell, L. Induction and properties of a temperate bacteriophage from Bacillus stearothermophilus. J. Bacteriol. 1965, 89, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.; Campbell, L. Biochemical changes in lysogenic Bacillus stearothermophilus after bacteriophage induction. J. Bacteriol. 1965, 90, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Welker, E. Transduction in Bacillus stearothermophilus. J. Bacteriol. 1988, 170, 3761–3764. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, F.; Donelli, G. Some properties of a thermophilic phage DNA. Arch. Biochem. Biophys. 1968, 125, 376–377. [Google Scholar] [CrossRef]
- Egbert, L.N.; Mitchell, H.K. Characteristics of Tφ3, a bacteriophage for Bacillus stearothermophilus. J. Virol. 1967, 1, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Egbert, L.N. Characteristics of the deoxyribonucleic acid of T phi 3, a bacteriophage for Bacillus stearothermophilus. J. Virol. 1969, 3, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Humbert, R.D.; Fields, M.L. Study of two bacteriophages of Bacillus stearothermophilus strain NCA1518. J. Virol. 1972, 9, 397–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurgen, C.L.; Edero, M. The influence of inorganic ions on the heat stability of a thermophilic bacteriophage. Physiol. Plant. 1972, 27, 182–186. [Google Scholar]
- Reanney, D.C.; Marsh, S.C.N. The ecology of viruses attacking Bacillus stearothermophilus in soil. Soil Biol. Biochem. 1973, 5, 399–406. [Google Scholar] [CrossRef]
- Sakaki, Y.; Oshima, T. A new lipid-containing phage infecting acidophilic thermophilic bacteria. Virology 1976, 75, 256–259. [Google Scholar] [CrossRef]
- Sharp, R.J.; Bown, K.J.; Atkinson, A. Phenotypic and genotypic characterization of some thermophilic species of Bacillus. J. Gen. Microbiol. 1980, 117, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.; Sharp, R.J. Taxonomic and related aspects of thermophiles within the Genus Bacillus. In The Aerobic Endospore-Forming Bacteria: Classification and Identification; Berkeley, R.C.W., Goodfellow, M., Eds.; Academic Press: London, UK, 1981; pp. 251–296. [Google Scholar]
- Sharp, R.J.; Woodrow, R. Numerical taxonomy of Bacillus thermophiles. In Abstracts of the XHIth International Congress of Microbiology Boston USA; Nottingham Trent University: Nottingham, UK, 1982; pp. 61–62. [Google Scholar]
- Wang, Y.; Zhang, X. Characterization of a novel portal protein from deep-sea thermophilic bacteriophage GVE2. Gene 2008, 421, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Ye, T.; Zhang, X. Roles of bacteriophage GVE2 endolysin in host lysis at high temperatures. Microbiology 2013, 159, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wu, S.; Xie, L. Complete genome sequence and proteomic analysis of a thermophilic bacteriophage BV1. Acta Oceanol. Sin. 2010, 29, 84–89. [Google Scholar] [CrossRef]
- Fujimoto, D.F.; Higginbotham, R.H.; Sterba, K.M.; Maleki, S.J.; Segall, A.M.; Smeltzer, M.S.; Hurlburt, B.K. Staphylococcus aureus SarA is a regulatory protein responsive to redox and pH that can support bacteriophage lambda integrase-mediated excision/recombination. Mol. Microbiol. 2009, 74, 1445–1458. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J.C.; Lüder, G.; Stiege, A.C.; Chai, S.; Weise, F.; Trautner, T.A. The complete nucleotide sequence and functional organization of Bacillus subtilis bacteriophage SPP1. Gene 1997, 204, 201–212. [Google Scholar] [CrossRef]
- Levine, M.; Truesdall, S.; Ramakrishan, T.; Bronson, M.J. Dual control of lysogeny by bacteriophage P22: An antirepressor locus and its controlling elements. J. Mol. Biol. 1975, 91, 421–438. [Google Scholar] [CrossRef] [Green Version]
- van Zyl, L.J.; Sunda, F.; Taylor, M.P.; Cowan, D.; Trindade, M.I. Identification and characterization of a novel Geobacillus thermoglucosidasius bacteriophage, GVE3. Arch. Virol. 2015, 160, 2269–2282. [Google Scholar] [CrossRef] [Green Version]
- van Zyl, L.J.; Taylor, M.P.; Trindade, M. Engineering resistance to bacteriophage GVE3 in Geobacillus thermoglucosidasius. Appl. Microbiol. Biotechnol. 2016, 100, 1833–1841. [Google Scholar] [CrossRef]
- Maszewska, A. Bacteriophage associated polysaccharide depolymerases—Characteristics and application. Postepy Hig. Med. Dosw. 2015, 69, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.C.; Danchin, A. Base composition bias might result from competition for metabolic resources. TRENDS Genet. 2002, 18, 291–294. [Google Scholar] [CrossRef]
- Akhwale, J.K.; Rohde, M.; Rohde, C.; Bunk, B.; Sproer, C.; Klenk, H.-P.; Boga, H.I.; Wittmann, J. Comparative genomic analysis of eight novel haloalkaliphilic bacteriophages from Lake Elmenteita, Kenya. PLoS ONE 2019, 14, e0212102. [Google Scholar] [CrossRef]
- Mwirichia, R.; Cousin, S.; Muigai, A.W.; Boga, H.I.; Stackebrandt, E. Archaeal diversity in the haloalkaline Lake Elmenteita in Kenya. Curr. Microbiol. 2010, 60, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Streisinger, G.; Emrich, J.; Stahl, M.M. Chromosome structure in phage T4, iii. Terminal redundancy and length determination. Proc. Natl. Acad. Sci. USA 1967, 57, 292–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, J.; Dreiseikelmann, B.; Rohde, C.; Rohde, M.; Sikorski, J. Isolation and characterization of numerous novel phages targeting diverse strains of the ubiquitous and opportunistic pathogen Achromobacter xylosoxidans. PLoS ONE 2014, 9, e86935. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, K.S.; Yang, M.; Mao, C. Phage-Enabled Nanomedicine: From Probes to Therapeutics in Precision Medicine. Angew Chem. Int. Ed. Engl. 2017, 56, 1964–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustard, M.T.; Whiting, S.; Cowan, D.A.; Wright, P.C. Biodegradation of high-concentration isopropanol by a solvent-tolerant thermophile, Bacillus pallidus. Extremophiles 2002, 6, 319–323. [Google Scholar] [CrossRef]
- Taylor, M.P.; Eley, K.L.; Martin, S.; Tuffin, M.I.; Burton, S.G.; Cowan, D.A. Thermophilic ethanologenesis: Future prospects for second-generation bioethanol production. Trends Biotechnol. 2009, 27, 398–405. [Google Scholar] [CrossRef]
- Cuebasg, M.; Sannino, D.; Bini, B. Isolation and characterization of an arsenic resistant Geobacillus kaustophilus strain from geothermal soils. J. Basic Microbiol. 2011, 51, 364–371. [Google Scholar] [CrossRef]
- Feng, L.; Wang, W.; Cheng, J.; Ren, Y.; Zhao, G.; Gao, C.; Tang, Y.; Lui, X.; Han, W.; Peng, X.; et al. Genome and proteome of long-chain alkane degrading Geobacillus thermodenitrificans NG80-2 isolated from a deep-subsurface oil reservoir. Proc. Natl. Acad. Sci. USA 2007, 104, 5602–5607. [Google Scholar] [CrossRef] [Green Version]
- Moriello, V.S.; Lama, L.; Poli, A.; Gugliandolo, C.; Maugeri, T.L.; Gamacorta, A.; Nicolaus, B. Production of exopolysaccharides from a thermophilic microorganism isolated from a marine hot spring in flegrean areas. J. Ind. Microbiol. Biotechnol. 2003, 30, 95–101. [Google Scholar] [CrossRef]
- Novotny, J.F.; Perry, J.J. Characterization of bacteriocins from two strains of Bacillus thermoleovorans, a thermophilic hydrocarbon-utilizing species. Appl. Environ. Microbiol. 1992, 58, 2393–2396. [Google Scholar] [CrossRef] [Green Version]
- Jeżewska-Frąckowiak, J.; Seroczyńska, K.; Banaszczyk, J.; Jedrzejczak, G.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Korza, G.; De Marco, A.M.; Kuipers, O.P.; Li, Y.Q.; Setlow, P. Properties of spores of Bacillus subtilis with or without a transposon that decreases spore germination and increases spore wet heat resistance. J. Appl. Microbiol. 2021. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Mu, Y.; Cong, Y. Bacillus coagulans and its applications in medicine. Benef. Microbes 2019, 10, 679–688. [Google Scholar] [CrossRef]
- Logan, N.A. Bacillus and relatives in foodborne illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med. Virol. 2019, 29, e2041. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, H.; Mulvenna, N.; Sanz-Hernandez, M.; Zhang, P.; Li, Y.; Ma, J.; Wang, Y.; Matthews, S.; Wigneshweraraj, S.; et al. A Bacteriophage DNA Mimic Protein Employs a Non-specific Strategy to Inhibit the Bacterial RNA Polymerase. Front. Microbiol. 2021, 12, 692512. [Google Scholar] [CrossRef] [PubMed]
- Mulvenna, N.; Hantke, I.; Burchell, L.; Nicod, S.; Bell, D.; Turgay, K.; Wigneshweraraj, S. Xenogeneic modulation of the ClpCP protease of Bacillus subtilis by a phage-encoded adaptor-like protein. J. Biol. Chem. 2019, 294, 17501–17511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.F.; Mesa, V.; Estrada-Ortiz, N.; Lopez-Obando, M.; Gómez, A.; Plácido, J. Viruses in Extreme Environments, Current Overview, and Biotechnological Potential. Viruses 2021, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Maat, D.S.; Biggs, T.; Evans, C.; Van Bleijswijk, J.D.L.; Van Der Wel, N.N.; Dutilh, B.E.; Brussaard, C.P.D. Characterization and temperature dependence of arctic micromonas polaris viruses. Viruses 2017, 9, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, K.A.; Sutherland, I.W.; Clark, J.; Jones, M.V. Bacteriophage and associated polysaccharide depolymerases—Novel tools for study of bacterial biofilms. J. Appl. Microbiol. 1998, 85, 583–590. [Google Scholar] [CrossRef]
- Hughes, K.A.; Sutherland, I.W.; Clark, J.; Jones, M.V. Biofilm susceptibility to bacteriophage attack: The role of bacteriophageborne polysaccharide depolymerase. Microbiology 1998, 144, 3039–3047. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Preventing biofilms of clinically relevant organisms using bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Parasion, S.; Kwiatek, M.; Gryko, R.; Mizak, L.; Malm, A. Bacteriophages as an alternative strategy for fighting biofilm development. Pol. J. Microbiol. 2014, 63, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Szekera, K.; Zhou, X.; Schwab, T.; Casanueva, A.; Cowan, D.; Mikhailopulo, I.A.; Neubauer, P. Comparative investigations on thermostable pyrimidine nucleoside phosphorylases from Geobacillus thermoglucosidasius and Thermus thermophiles. J. Mol. Cat. B Enzym. 2012, 84, 27–34. [Google Scholar] [CrossRef]
- Julien, B. Characterization of the integrase gene and attachment site for the Myxococcus xanthus bacteriophage Mx9. J. Bacteriol. 2003, 185, 6325–6330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzolla, D.; Calì, S.; Spoldi, E.; Forti, F.; Sala, C.; Magnoni, F.; Dehò, G.; Ghisotti, D. Expression of phage P4 integrase is regulated negatively by both Int and Vis. J. Gen. Virol. 2006, 87, 2423–2431. [Google Scholar] [CrossRef]


{kind=link}
| CDS Name | CDS Length (bp) | Location in the Genome (bp) | CDS Arbitrary Orientation | Polypeptide Length (aa) | Predicted Polypeptide Molecular Weight (kDa) | Experimentally Determined Polypeptide Molecular Weight (kDa) | Predicted Isoelectric Point | Hypothetical Function (Analysis) | Confirmed by Proteomic Analysis |
|---|---|---|---|---|---|---|---|---|---|
| TP84_01 | 567 | 13–579 | + | 188 | 22.4 | 22.4 | 7.76 | terminase, small subunit | terminase, small subunit |
| TP84_02 | 1299 | 576–1874 | + | 432 | 50.0 | 50.1 | 7.74 | terminase, large subunit | terminase, large subunit |
| TP84_04 | 354 | 2586–2939 | + | 117 | 13.6 | ND | 4.36 | unknown | unknown |
| TP84_15 | 324 | 10358–10681 | + | 107 | 11.9 | ND | 6.26 | unknown | unknown |
| TP84_20 | 375 | 12687–13061 | + | 124 | 14.7 | 14.7 | 9.80 | unknown | unknown |
| TP84_26 | 2976 | 20179–23154 | + | 991 | 112.2 | 112.9 | 5.49 | glycosylase | glycosylase |
| TP84_27 | 432 | 23240–23671 | + | 143 | 15.5 | 15.5 | 6.20 | holin | holin |
| TP84_30 | 573 | 25329–25901 | + | 190 | 21.7 | 21.7 | 4.98 | unknown | unknown |
| TP84_33 | 291 | 26597–26887 | + | 96 | 11.0 | ND | 5.10 | unknown | unknown |
| TP84_41 | 273 | 28824–29096 | + | 90 | 10.6 | ND | 9.69 | unknown | unknown |
| TP84_43 | 309 | 29330–29639 | + | 102 | 11.0 | ND | 9.81 | unknown | unknown |
| TP84_48 | 2265 | 30527–32791 | + | 754 | 86 | ND | 7.09 | unknown | unknown |
| TP84_49 | 474 | 32871–33344 | + | 157 | 18.0 | ND | 5.85 | unknown | unknown |
| TP84_58 | 318 | 37942–38259 | + | 105 | 12.3 | ND | 4.92 | replicative helicase inhibitor | replicative helicase inhibitor |
| TP84_59 | 1305 | 38259–39563 | + | 434 | 49.1 | 49.2 | 5.53 | replicative DNA helicase | replicative DNA helicase |
| TP84_60 | 672 | 39632–40303 | + | 223 | 26.4 | 26.4 | 8.41 | unknown | unknown |
| TP84_61 | 435 | 40281–40715 | + | 144 | 16.7 | ND | 9.08 | HNH homing endonuclease | HNH homing endonuclease |
| TP84_70 | 330 | 43704–44033 | + | 109 | 12.7 | ND | 9.47 | unknown | unknown |
| TP84_73 | 264 | 44746–45009 | + | 87 | 10.4 | ND | 5.60 | unknown | unknown |
| N° | Bacteriophage Species | Virus Family, Genus | GenBank Accession Number | Host (Used for Propagation) | Genome | Original Discovery Reference | Isolate Location | Life Cycle | Growth Temperature (°C, Optimal/Range) and pH (Optimal/Range) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Type and Size [bp] | ORFs | G+C [%] | |||||||||
| 1 | ‘thermophilic lytic principle’ | ND | ND | B. stearothermophilus T60 | ND | ND | ND | Koser, 1926 | sewage polluted river water (USA) | lytic | 52–60, app. 7 |
| 2 | ‘thermophilic bacteriophage’ | ND | ND | ND | ND | ND | ND | Adant, 1928 | ND | lytic | 52–55 |
| 3 | ‘thermophilic bacteriophage’ | ND | ND | thermophilic bacterium no. 10 | ND | ND | ND | White et al., 1954, 1955 | greenhouse soil (USA) | lytic | 65 (50–70), 7 |
| 4 | ‘thermophilic bacteriophage’ | ND | ND | Bacillus sp. | DNA, RNA(?) | ND | ND | Onodera; 1959; Onodera 1961 | compost (Japan) | lytic | 65 (55–70), 7.2 |
| 5 | TP-84 | Siphoviridae, Tp84virus | KY565347.1 | B. stearothermophilus strain 10 | circular, dsDNA, 47,718 | 81 | 54.5 | Saunders & Campbell, 1964 | greenhouse soil (USA) | lytic | 58 (43–76), 7.2 |
| 6 | φμ-4 | ND | ND | B. stearothermophilus | ND | ND | ND | Shafia & Thompson, 1964 | ND | lytic/lysogenic | 50–65, 7 |
| 7 | TP-1 | Siphoviridae (putative) | ND | B. stearothermophilus | dsDNA, app. 18,516 bp, (MW 12.1 Mda) | ND | 42 | Welker and Campbell, 1965 | ND | lysogenic/lytic | 55 (50–65), 7 |
| 8 | ST1 | Siphoviridae Myoviridae (putative) | ND | B. stearothermophilus strain S13 | dsDNA | ND | 43 | Carnevali & Donelli, 1968 | ND | lytic | 60, app. 7 |
| 9 | Tφ3 | Siphoviridae | ND | B. stearothermophilus ATCC 8005 SR | ds DNA, app. 35,700 (MW app. 23.2 MDa) | ND | 40.2 | Egbert&Mitchel, 1967; Egbert, 1969 | soil (USA) | lytic | 60, 7.3 |
| 10 | GH5 | ND | ND | B. stearothermophilus NCA1518 | ND | ND | ND | Humbert & Fields, 1972 | greenhouse soil (USA) | lytic | 42.5–67, app. 7 |
| 11 | GH8 | Siphoviridae | ND | B.stearothermophilus NCA1518 | ND | ND | ND | Humbert & Fields, 1972 | greenhouse soli (USA) | lytic | 42.5–67, app. 7 |
| 12 | PhB1 | Siphoviridae | ND | Bacillus sp. strain B | ND | ND | ND | Junger & Edebo, 1972 | farm soil (Sweden) | lytic | 55, 7.3 |
| 13 | D5 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 14 | D6 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 15 | D7 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 16 | D8 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 17 | φNS11 | Sphaerolipoviridae, (putative) | ND | B.acidocaldarius TA6 | dsDNA | ND | ND | Sakaki & Oshima, 1976 | hot spring (Beppu, Japan) | lytic | 60, 3.5 (2–5) |
| 18 | JS001 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | ND | lytic/lysogenic | 55 (50–70), 7.3 ± 0.2 |
| 19 | JS004 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 20 | JS005 | ND | ND | B. thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 21 | JS006 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 22 | JS007 | ND | ND | Bacillus thermophile RS 240 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 23 | JS008 | ND | ND | Bacillus thermophile RS 241 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 24 | JS009 | ND | ND | Bacillus thermophile RS 242 | dsDNA | ND | ND | Sharp et al., 1986 | stable manure | lytic | 55 (50–70), 7.3 ± 0.2 |
| 25 | JS010 | ND | ND | Bacillus thermophile RS 242 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 26 | JS011 | unclassified family | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 27 | JS012 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 28 | JS013 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | soil | lytic | 55 (50–70), 7.3 ± 0.2 |
| 29 | JS014 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 30 | JS015 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 31 | JS017 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 32 | JS018 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 33 | JS019 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 34 | JS020 | ND | ND | B.s caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 35 | JS021 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 36 | JS022 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 37 | JS023 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 38 | JS024 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 39 | JS025 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 40 | JS026 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 41 | JS027 | ND | ND | Bacillus thermophile RS 241 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 42 | BVW1 (W1) | Siphoviridae | ND | Bacillus sp. w13 | dsDNA, app. 18 kb | ND | ND | Liu et al., 2006 | deep-sea hydrothermal fields (West Pacific) | lytic | 60, 7.0 |
| 43 | GVE1 (E1) | Siphoviridae | ND | Geobacillus sp. E 26323 | dsDNA, app. 41 kb | ND | ND | Liu et al., 2006 | deep-sea hydrothermal fields (East Pacific) | lytic | 60, 7.0 |
| 44 | GVE2 (E2) | Siphoviridae, unclassified Siphoviridae | NC_009552 DQ453159 | Geobacillus sp. E 263 | linear, dsDNA, 40,863 | 62 | 44.8 | Liu & Zhang, 2008a,b | deep sea (China) | lysogenic | 65, 7.0 |
| 45 | GBSV1 | Myoviridae, Svunavirus | Geobacillus sp. 6k512 | linear, dsDNA, 34,683 | 54 | 44.4 | Liu et al., 2009, 2010 | off shore hot spring, (Xiamen, China) | lytic | 65, 7.2 | |
| 46 | BV1 | Myoviridae ,Svunavirus | NC_009737.2, DQ840344 | Geobacillus sp. 6k512 | linear, dsDNA, 35,055 | 54 | 44.4 | Liu et al., 2009, 2010 | off shore hot spring, (Xiamen, China) | lytic | 65, 7.2 |
| 47 | D6E | Myoviridae | NC_019544 | Geobacillus sp. E 26323 | circular, dsDNA, 49,335 | 49 | 46 | Wang & Zhang, 2010 | deep-sea hydrothermal fields (East Pacific) | lytic | 65, 7.0 |
| 48 | ϕOH2 (phiOH2) | Siphoviridae | AB823818, NC_021784 | G. kaustophilus GBlys, G. kaustophilus NBRC 102445(T), lysogenic G. kaustophilus GBlys) | dsDNA, 38,099 | 60 | 45 | Doi et al., 2013 | hot spring sediment (Japan) | lytic/lysogenic | 55 |
| 49 | GBK2 | Siphoviridae | KJ159566 | G. kaustophilus | Circularly permuted, dsDNA, 39,078 | 62 | 43 | Marks & Hamilton, 2014 | compost (Cary, NC, USA) | lytic | 55, 7.3 |
| 50 | GVE3 (E3) | Siphoviridae | NC_029073, KP144388 | G. thermoglucosidasius | dsDNA 141,298 | 202 | 29.6 | Van Zyl et al., 2015 | ND | lytic/lysogenic | 60, 7.3 |
| 51 | AP45 | Siphoviridae | KX965989 | Aeribacillus sp. CEMTC656 | dsDNA 51,606 | 71 | 38.3 | Morozowa et al., 2019 | soil (Valley of Geysers, Kamchatka, Russia) | lytic/lysogenic | 55, 7.5 |
| 52 | vB_Bps-36 | ND | MH884513 | B. pseudalcaliphilus | dsDNA 50,485 | 68 | 41.1 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 53 | vB_BpsM-61 | ND | MH884514 | B. pseudofirmus | dsDNA 48,160 | 75 | 43.5 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 54 | vB_BboS-125 | ND | MH884509 | B. bogoriensis | dsDNA 58,528 | 81 | 48.6 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 55 | vB_BcoS-136 | ND | MH884508 | B. cohnii | dsDNA 160,590 | 240 | 32.2 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 56 | vB_BpsS-140 | ND | MH884512 | B. pseudalcaliphilus | dsDNA 55,091 | 68 | 39.8 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sobolewski, I.; Skowron, P.M. Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms 2021, 9, 1522. https://doi.org/10.3390/microorganisms9071522
Łubkowska B, Jeżewska-Frąckowiak J, Sobolewski I, Skowron PM. Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms. 2021; 9(7):1522. https://doi.org/10.3390/microorganisms9071522
Chicago/Turabian StyleŁubkowska, Beata, Joanna Jeżewska-Frąckowiak, Ireneusz Sobolewski, and Piotr M. Skowron. 2021. "Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review" Microorganisms 9, no. 7: 1522. https://doi.org/10.3390/microorganisms9071522
APA StyleŁubkowska, B., Jeżewska-Frąckowiak, J., Sobolewski, I., & Skowron, P. M. (2021). Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms, 9(7), 1522. https://doi.org/10.3390/microorganisms9071522