
Analyses of Virulence Genes of Clavibacter michiganensis subsp. michiganensis Strains Reveal Heterogeneity and Deletions That Correlate with Pathogenicity

 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Virulence Assay in Tomato Seedlings
2.3. Statistical Analysis
2.4. Assay for Cellulase Activity
2.5. PCR-Amplification of Virulence Genes
2.6. Sequence and Cluster Analysis of Virulence Genes
2.7. Analysis of Virulence Genes and Their Proteins in Cmm Genomes
3. Results
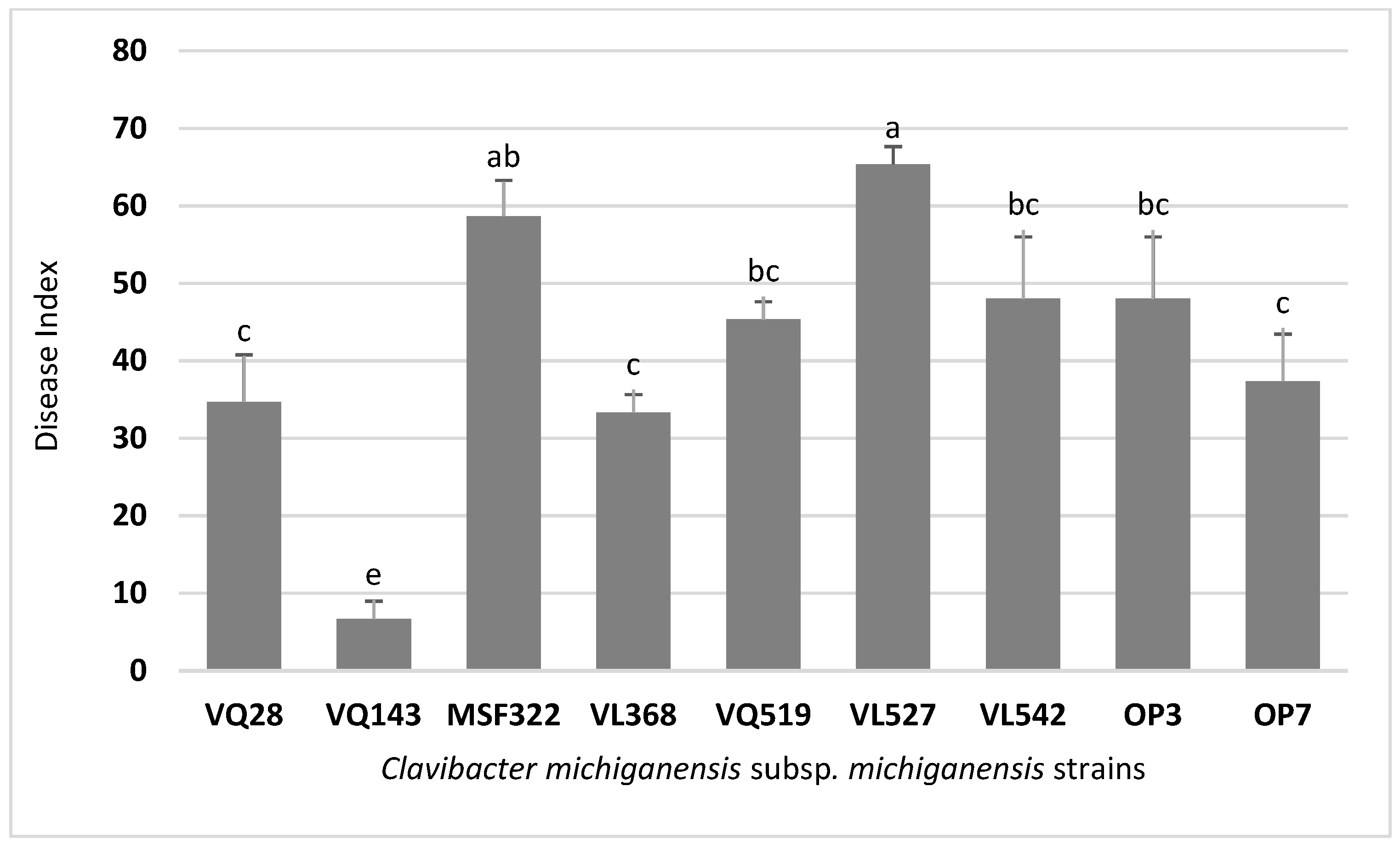
3.1. Disease Severity Varied between the Strains
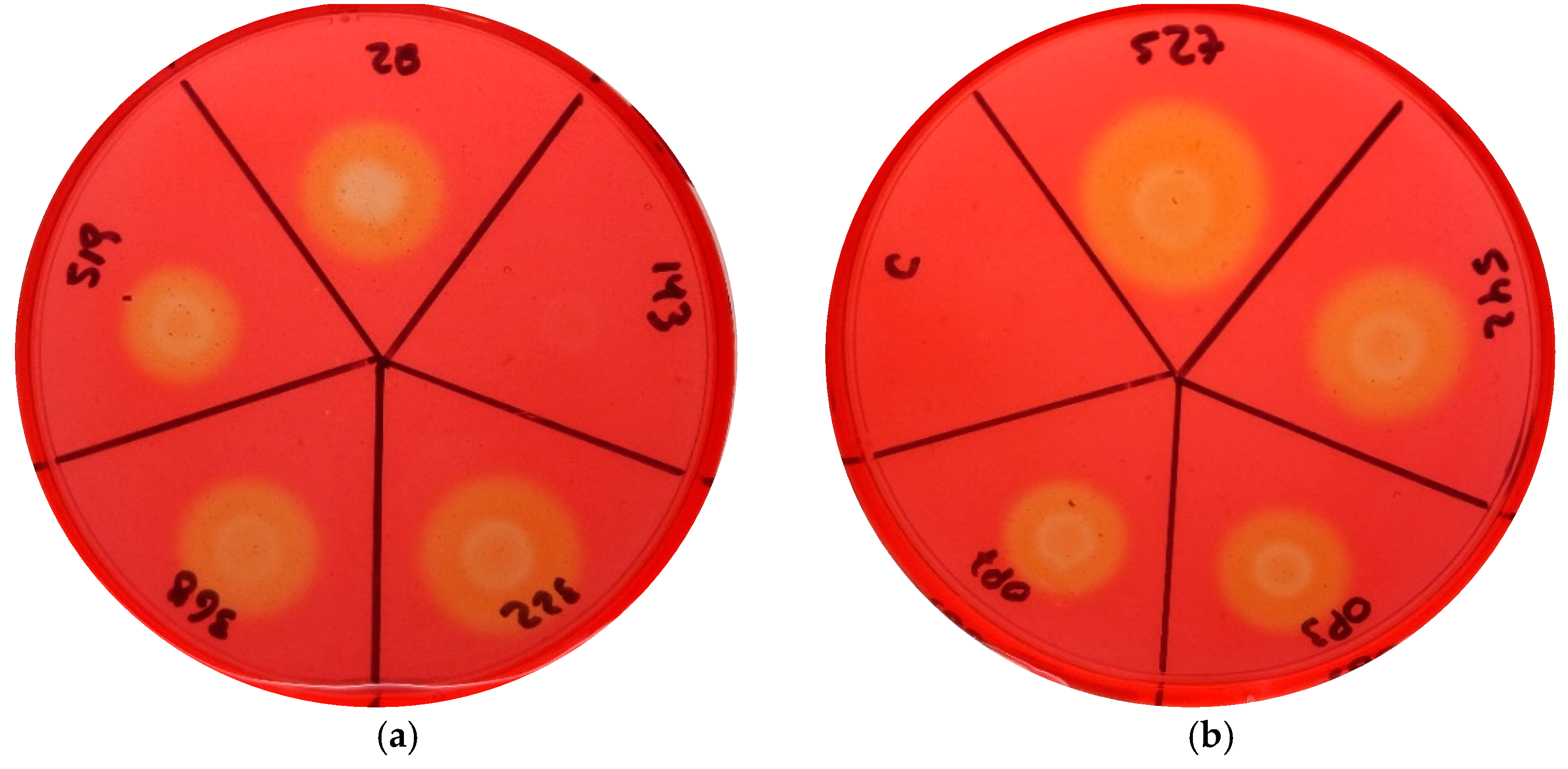
3.2. Cellulase Activity Was Higher in More Virulent Strains
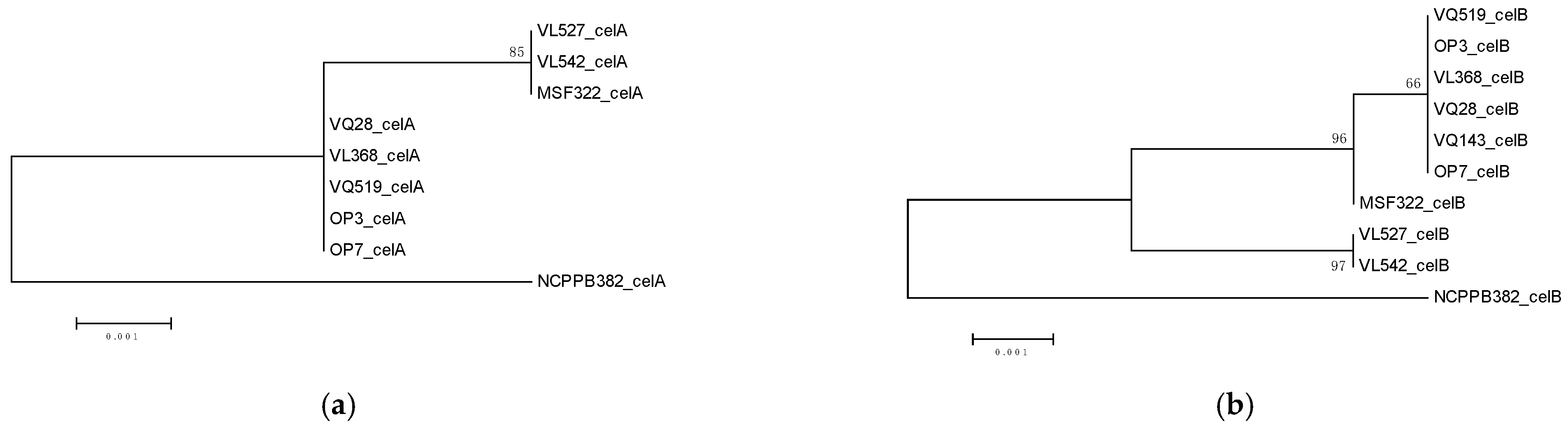
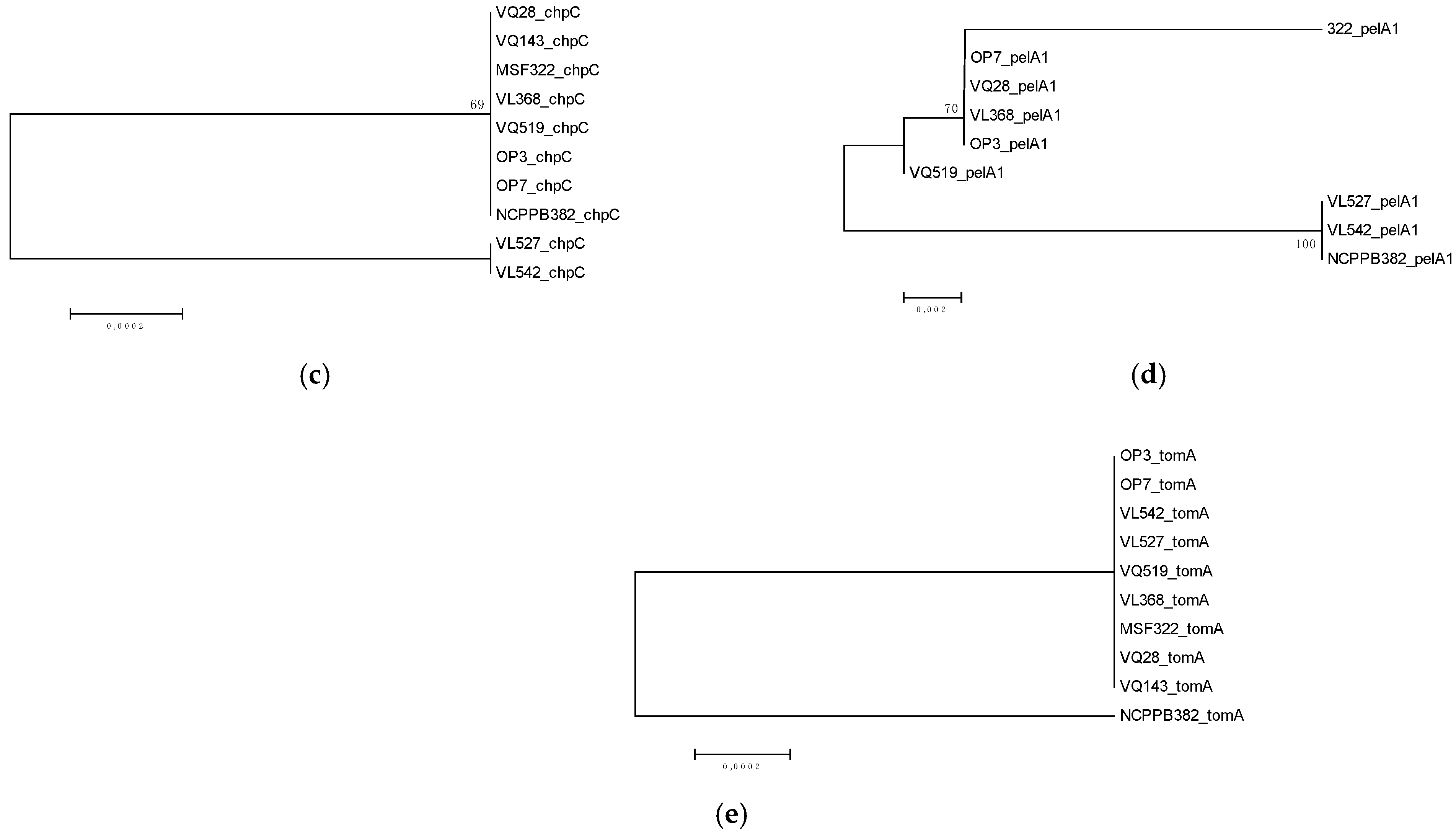
3.3. Virulence Genes’ Repertoire and Cluster Analysis Increased Variation between Strains
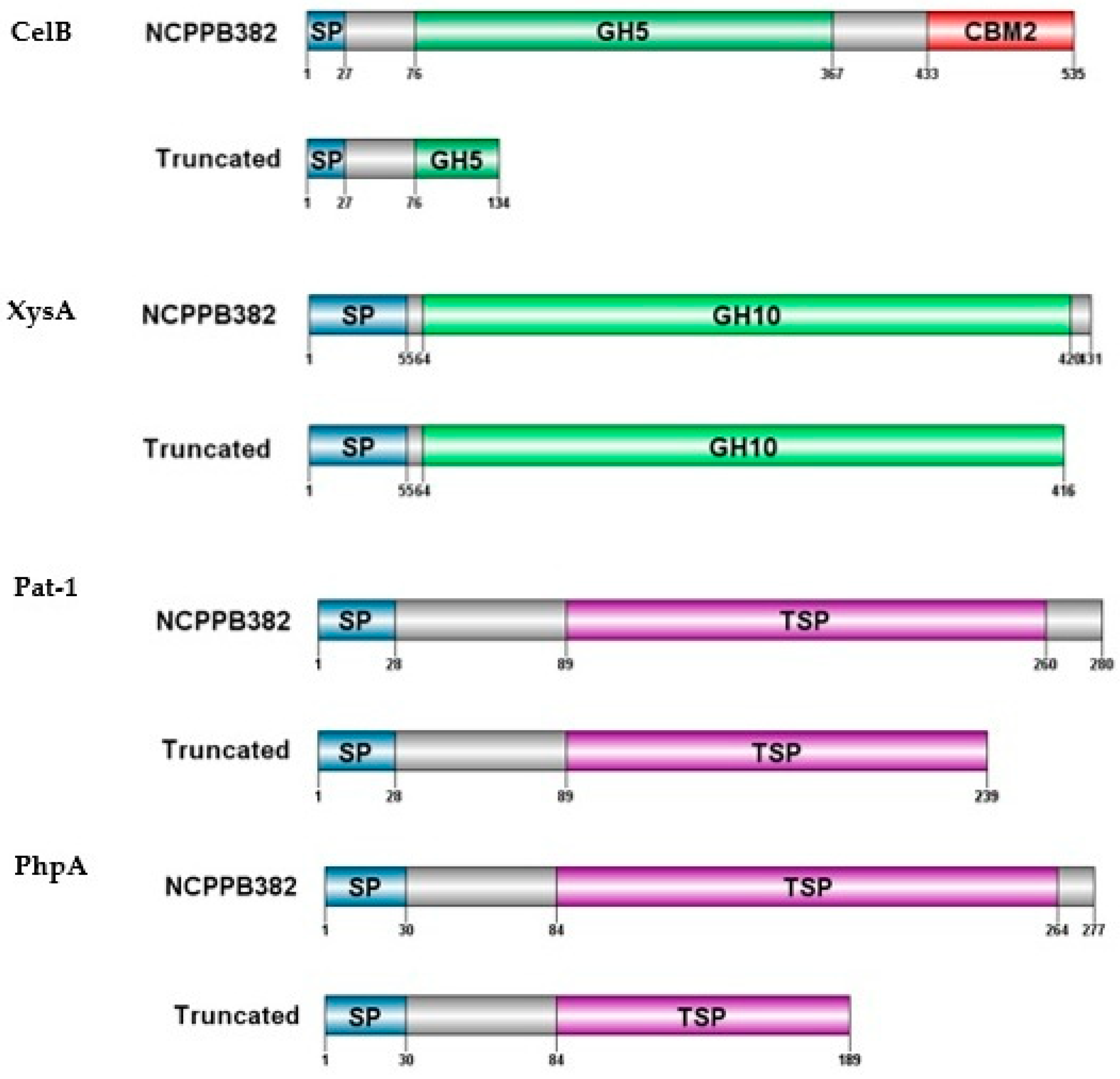
3.4. Deletions Detected in the Sequences of Virulence Genes celB, xysA, pat-1, and phpA Truncate the Respective Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wassermann, E.; Montecchia, M.S.; Correa, O.S.; Damian, V.; Romero, A.M. Clavibacter michiganensis subsp. michiganensis strains virulence and genetic diversity. A first study in Argentina. Eur. J. Plant Pathol. 2017, 149, 35–42. [Google Scholar] [CrossRef]
- Thapa, S.P.; Pattathil, S.; Hahn, M.G.; Jacques, M.A.; Gilbertson, R.L.; Coaker, G. Genomic analysis of Clavibacter michiganensis reveals insight into virulence strategies and genetic diversity of a gram-positive bacterial pathogen. Mol. Plant Microbe Intercat. 2017, 30, 786–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartemann, K.H.; Abt, B.; Bekel, T.; Burger, A.; Engemann, J.; Flügel, M.; Gaigalat, L.; Goesmann, A.; Gräfen, I.; Kalinowski, J.; et al. The genome sequence of the tomato-pathogenic actinomycete Clavibacter michiganensis subsp. michiganensis NCPPB382 reveals a large island involved in pathogenicity. J. Bacteriol. 2008, 190, 2138–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, M.; Macdonald, J.; Liu, P.; Weselowski, B.; Yuan, Z.C. Clavibacter michiganensis ssp. michiganensis: Bacterial canker of tomato, molecular interactions and disease management. Mol. Plant Pathol. 2018, 19, 2036–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleitman, F.; Barash, I.; Burger, A.; Iraki, N.; Falah, Y.; Sessa, G.; Weinthal, D.; Chalupowicz, L.; Gartemann, K.-H.; Eichenlaub, R.; et al. Characterization of a Clavibacter michiganensis subsp. michiganensis population in Israel. Eur. J. Plant Pathol. 2008, 121, 463–475. [Google Scholar] [CrossRef]
- Milijašević-Marčić, S.; Gartemann, K.-H.; Frohwitter, J.; Eichenlaub, R.; Todorović, B.; Rekanović, E.; Potočnik, I. Characterization of Clavibacter michiganensis subsp. michiganensis strains from recent outbreaks of bacterial wilt and canker in Serbia. Eur. J. Plant Pathol. 2012, 134, 697–711. [Google Scholar] [CrossRef]
- Bella, P.; Ialacci, G.; Licciardello, G.; La Rosa, R.; Catara, V. Characterization of atypical Clavibacter michiganensis subsp. michiganensis populations in greenhouse tomatoes in Italy. J. Plant Pathol. 2012, 94, 635–642. [Google Scholar]
- Ialacci, G.M.; Bella, P.; Licciardello, G.; Strano, C.P.; Eichenlaub, R.; Gartemann, K.H.; La Rosa, R.; Catara, V. Clonal populations of Clavibacter michiganensis subsp. michiganensis are responsible for the outbreaks of bacterial canker in greenhouse tomatoes in Italy. Plant Pathol. 2016, 65, 484–495. [Google Scholar] [CrossRef]
- Tancos, M.A.; Lange, H.W.; Smart, C.D. Characterizing the genetic diversity of the Clavibacter michiganensis subsp. michiganensis population in New York. Phytopathology 2015, 105, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Dreier, J.; Meletzus, D.; Eichenlaub, R. Characterization of the plasmid-encoded virulence region pat-1 of phytopathogenic Clavibacter michiganensis subsp. michiganensis. Mol. Plant Microbe Interact. 1997, 2, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Jahr, H.; Dreier, J.; Meletzus, D.; Bahro, R.; Eichenlaub, R. The endo-beta-1,4-glucanase of Clavibacter michiganensis subsp. michiganensis is a pathogenicity determinant required for the induction of bacterial wilt of tomato. Mol. Plant Microbe Interact. 2000, 13, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.S.; Oh, E.J.; Lee, H.B.; Oh, C.S. Functional characterization of two cellulase genes in the Gram-positive pathogenic bacterium Clavibacter michiganensis for wilting in tomato. Mol. Plant Microbe Interact. 2019, 32, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A. Microbial minimalism: Genome reduction in bacterial pathogens. Cell 2002, 108, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.C.; Amiri, H.; Andersson, S.G. Genome deterioration: Loss of repeated sequences and accumulation of junk DNA. Genetica 2002, 115, 1–12. [Google Scholar] [CrossRef]
- Valenzuela, M.; Besoain, X.; Durand, K.; Cesbron, S.; Fuentes, S.; Claverías, F.; Jacques, M.A.; Seeger, M. Clavibacter michiganensis subsp. michiganensis strains from central Chile exhibit low genetic diversity and sequence types match strains in other parts of the world. Plant Pathol. 2018, 67, 1944–1954. [Google Scholar] [CrossRef]
- Méndez, V.; Valenzuela, M.; Salvà-Serra, F.; Jaén-Luchoro, D.; Besoain, X.; Moore, E.R.; Seeger, M. Comparative Genomics of Pathogenic Clavibacter michiganensis subsp. michiganensis Strains from Chile Reveals Potential Virulence Features for Tomato Plants. Microorganisms 2020, 8, 1679. [Google Scholar] [CrossRef]
- Meletzus, D.; Bermpohl, A.; Dreier, J.; Eichenlaub, R. Evidence for plasmid-encoded virulence factors in the phytopathogenic bacterium Clavibacter michiganensis subsp. michiganensis NCPPB382. J. Bacteriol. 1993, 175, 2131–2136. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L.; et al. InterPro: The integrative protein signature database. Nucleic Acids Res. 2009, 37, D211–D215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Cohen-Kandli, M.; Dror, O.; Eichenlaub, R.; Gartemann, K.H.; Sessa, G.; Barash, I.; Manulis-Sasson, S. Sequential expression of bacterial virulence and plant defense genes during infection of tomato with Clavibacter michiganensis subsp. michiganensis. Phytopathology 2010, 100, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Burger, A.; Gräfen, I.; Engemann, J.; Niermann, E.; Pieper, M.; Kirchner, O.; Gartemann, K.-H.; Eichenlaub, R. Identification of homologues to the pathogenicity factor Pat-1, a putative serine protease of Clavibacter michiganensis subsp. michiganensis. Microbiol. Res. 2005, 160, 417–427. [Google Scholar] [CrossRef]
- Valenzuela, M.; Méndez, V.; Montenegro, I.; Besoain, X.; Seeger, M. Streptomycin resistance in Clavibacter michiganensis subsp. michiganensis strains from Chile is related to a rpsL gene mutation. Plant Pathol. 2018, 68, 426–433. [Google Scholar] [CrossRef]
- Quesada-Ocampo, L.M.; Landers, N.A.; Lebeis, A.C.; Fulbright, D.W.; Hausbeck, M.K. Genetic structure of Clavibacter michiganensis subsp. michiganensis populations in Michigan commercial tomato fields. Plant Dis. 2012, 96, 788–796. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara-Bell, J.; Alvarez, A.M. Seed-associated subspecies of the genus Clavibacter are clearly distinguishable from Clavibacter michiganensis subsp. michiganensis. Int. J. Syst. Evol. Microbiol. 2015, 65, 811–826. [Google Scholar] [CrossRef]
- Mira, A.; Ochman, H.; Moran, N.A. Deletional bias and the evolution of bacterial genomes. Trends Genet. 2001, 17, 589–596. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin (Region/Locality) | Year of Isolation | Sequence Type (ST)-MLST | GenBank Accession (Genome) |
|---|---|---|---|---|
| VQ28 | Valparaíso/Quillota | 1996 | 32 | CP076349-CP076351 |
| VQ143 | Valparaíso/Quillota | 2000 | 32 | CP076352-CP076353 |
| VL368 | Valparaíso/Limache | 2007 | 32 | --- |
| VQ519 | Valparaíso/Quillota | 2011 | 32 | --- |
| OP3 | O’Higgins/Pichidegua | 2015 | 32 | WTCS00000000 |
| OP7 | O’Higgins/Pichidegua | 2015 | 32 | --- |
| MSF322 | Maule/Sagrada Familia | 2005 | 36 | CP047051–CP047053 |
| VL527 | Valparaíso/Limache | 2012 | 18 | CP047054–CP047055 |
| VL542 | Valparaíso/Limache | 2013 | 18 | --- |
| Gene | Gene Product | Gene Location | Primer Sequences (5′→3′) | Tm (°C) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|---|
| chpC | Pat-1 type serine protease | PAI | F: GCTCTTGGGCTAATGGCCG | 60 | 639 | [5] |
| R: GTCAGTTGTGGAAGATGCTG | ||||||
| tomA | Tomatinase | PAI | F: CGAACTCGACCAGGT TCTC | 60 | 529 | [5] |
| R: GGTCTCACGATCGGATCC | ||||||
| celA | Cellulase | pCM1 | F: GTAGGGCACGCATTTCAGAG | 58 | 1240 | [2] |
| R: CAATGTCCTTCTTCGCCAGG | ||||||
| pat-1 | Pat-1 type serine protease | pCM2 | F: TGTAGACCGTATAGCCCGTG | 55 | 850 | [2] |
| R: CCTGAGACCTATTACCGCCC | ||||||
| phpA | Pat-1 type serine protease | pCM2 | F: CATTGGGTTGCTGTGTCGTT | 60 | 605 | This study |
| R: GAACGTTTCCGCTTCGACTTC | ||||||
| phpB | Pat-1 type serine protease | pCM2 | F: GAGAACCAGCCTTCCCGTTC | 60 | 596 | This study |
| R: CCACGAATCCTCCTGAGTCG | ||||||
| celB | Cellulase | Chromosome | F: GGCTCGACAAGATCACCCTC | 60 | 1283 | This study |
| R: ACCGACATGGACGGTCTGA | ||||||
| xysA | Xylanase | Chromosome | F: CGATTCGACTTCTCGGGCAT | 60 | 683 | This study |
| R: TCGTCCGGGTTCGAGTAGAT | ||||||
| pelA1 | Pectinase | PAI | F: AGAACGTGATCATCGGCTCG | 60 | 520 | This study |
| R: TGTTCGAAGAGGATGGTGGC | ||||||
| pelA2 | Pectinase | PAI | F: ATCAACCATCTCGACCCTCCC | 60 | 685 | This study |
| R: GTAACTGAAGTCGCACACCC |
| Cell Wall-Degrading Enzymes | Serine-Proteases | Tomatinase | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Sequence Type | celA | celB | pelA1 | pelA2 | xysA | chpC | pat-1 | phpA | phpB | tomA | Halo Diam. * |
| VQ28 | 32 | + | + | + | + | + | + | - | - | - | + | 17 |
| VQ143 | 32 | - | + | - | - | + | + | + | - | - | + | 0 |
| VL368 | 32 | + | + | + | + | + | + | + | - | - | + | 17 |
| VQ519 | 32 | + | + | + | + | + | + | + | - | - | + | 13 |
| OP3 | 32 | + | + | + | - | + | + | + | - | - | + | 17 |
| OP7 | 32 | + | + | + | - | + | + | + | - | - | + | 17 |
| MSF322 | 36 | + | + | + | + | + | + | + | + | + | + | 19 |
| VL527 | 18 | + | + | + | + | + | + | + | + | + | + | 21 |
| VL542 | 18 | + | + | + | + | + | + | + | + | + | + | 18 |
| Strain | Gene | %ID NCPPB382 | Coverage | Observations |
|---|---|---|---|---|
| MSF322 | celA | 99.55 | 100% | |
| celB | 98.88 | 100% | 1 deletion, pseudogene | |
| pelA1 | 97.77 | 100% | ||
| pelA2 | 98.36 | 100% | ||
| xysA | 99.54 | 100% | ||
| xysB | 99.85 | 100% | ||
| pat-1 | 99.76 | 100% | 2 deletions, pseudogene | |
| phpA | 99.52 | 100% | 4 deletions, pseudogene | |
| phpB | 100.0 | 100% | ||
| VL527 | celA | 99.55 | 100% | |
| celB | 99.07 | 100% | ||
| pelA1 | 100.0 | 100% | ||
| pelA2 | 97.65 | 100% | ||
| xysA | 99.54 | 100% | 1 deletion, pseudogene | |
| xysB | 99.90 | 100% | ||
| pat-1 | 100.0 | 100% | ||
| phpA | 100.0 | 100% | ||
| phpB | 100.0 | 100% | ||
| OP3 | celA | 99.64 | 100% | |
| celB | 98.82 | 100% | 1 deletion, pseudogene | |
| pelA1 | 98.59 | 100% | ||
| pelA2 | 93.16 | 97% | ||
| xysA | 99.54 | 100% | 1 deletion, pseudogene | |
| xysB | 99.85 | 100% | ||
| pat-1 | 100.0 | 100% | ||
| phpA | NF | -- | ||
| phpB | NF | -- | ||
| VQ28 | celA | 99.64 | 100% | |
| celB | 98.82 | 100% | 1 deletion, pseudogene | |
| pelA1 | 98.59 | 100% | ||
| pelA2 | 98.24 | 100% | ||
| xysA | 99.54 | 100% | 1 deletion, pseudogene | |
| xysB | 99.85 | 100% | ||
| pat-1 | NF | -- | ||
| phpA | NF | -- | ||
| phpB | NF | -- | ||
| VQ143 | celA | NF | -- | |
| celB | 98.82 | 100% | 1 deletion, pseudogene | |
| pelA1 | NF | -- | ||
| pelA2 | NF | -- | ||
| xysA | 99.54 | 100% | 1 deletion, pseudogene | |
| xysB | 99.85 | 100% | ||
| pat-1 | 100.0 | 100% | ||
| phpA | NF | -- | ||
| phpB | NF | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, M.; González, M.; Velásquez, A.; Dorta, F.; Montenegro, I.; Besoain, X.; Salvà-Serra, F.; Jaén-Luchoro, D.; Moore, E.R.B.; Seeger, M. Analyses of Virulence Genes of Clavibacter michiganensis subsp. michiganensis Strains Reveal Heterogeneity and Deletions That Correlate with Pathogenicity. Microorganisms 2021, 9, 1530. https://doi.org/10.3390/microorganisms9071530
Valenzuela M, González M, Velásquez A, Dorta F, Montenegro I, Besoain X, Salvà-Serra F, Jaén-Luchoro D, Moore ERB, Seeger M. Analyses of Virulence Genes of Clavibacter michiganensis subsp. michiganensis Strains Reveal Heterogeneity and Deletions That Correlate with Pathogenicity. Microorganisms. 2021; 9(7):1530. https://doi.org/10.3390/microorganisms9071530
Chicago/Turabian StyleValenzuela, Miryam, Marianela González, Alexis Velásquez, Fernando Dorta, Iván Montenegro, Ximena Besoain, Francisco Salvà-Serra, Daniel Jaén-Luchoro, Edward R. B. Moore, and Michael Seeger. 2021. "Analyses of Virulence Genes of Clavibacter michiganensis subsp. michiganensis Strains Reveal Heterogeneity and Deletions That Correlate with Pathogenicity" Microorganisms 9, no. 7: 1530. https://doi.org/10.3390/microorganisms9071530
APA StyleValenzuela, M., González, M., Velásquez, A., Dorta, F., Montenegro, I., Besoain, X., Salvà-Serra, F., Jaén-Luchoro, D., Moore, E. R. B., & Seeger, M. (2021). Analyses of Virulence Genes of Clavibacter michiganensis subsp. michiganensis Strains Reveal Heterogeneity and Deletions That Correlate with Pathogenicity. Microorganisms, 9(7), 1530. https://doi.org/10.3390/microorganisms9071530