
A Long-Term Record of Quaternary Facies Patterns and Palaeonvironmental Trends from the Po Plain (NE Italy) as Revealed by Bio-Sedimentary Data

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
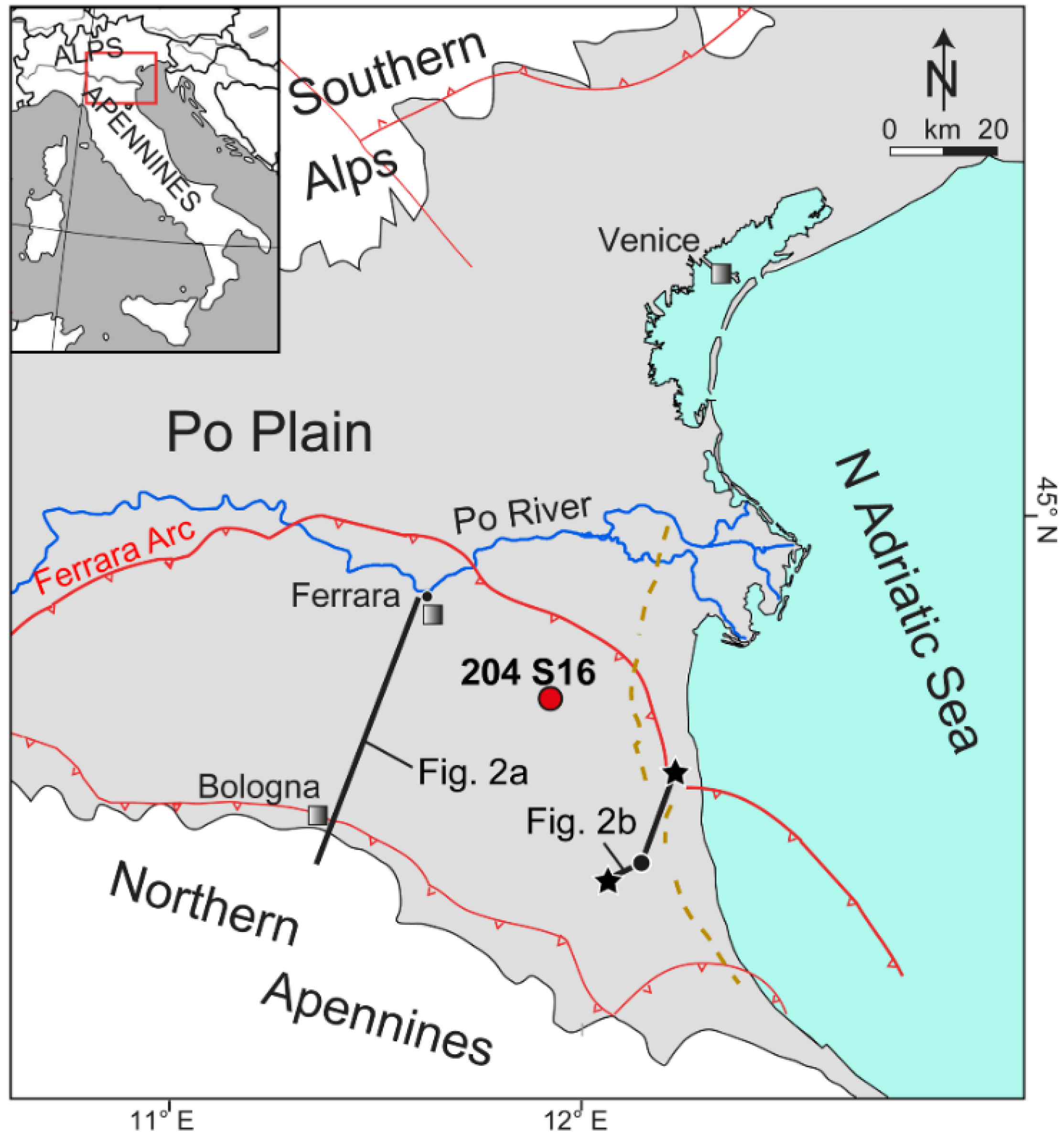
2. Study Area
2.1. Geological Context
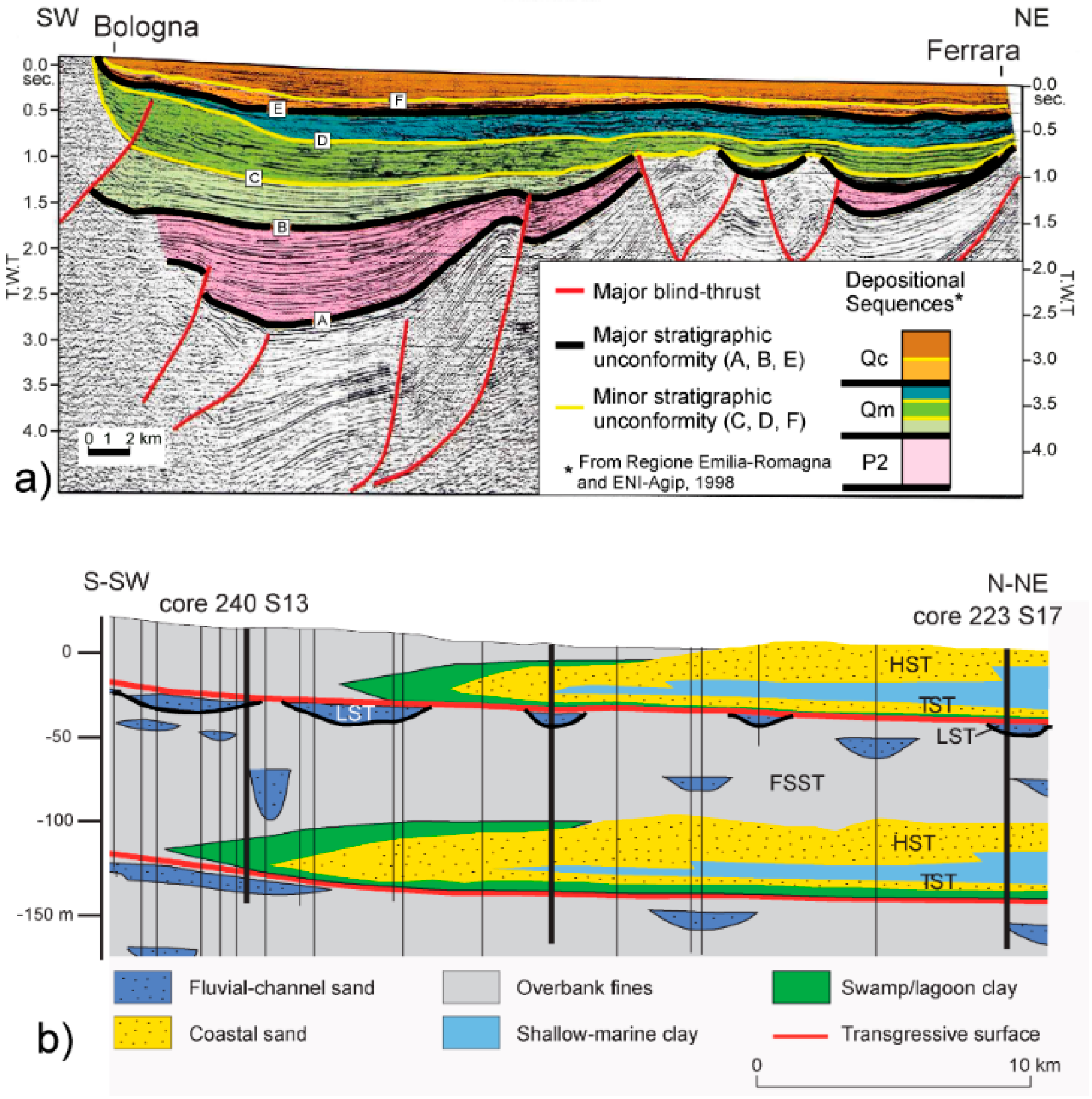
2.2. Stratigraphic Setting
3. Materials and Methods
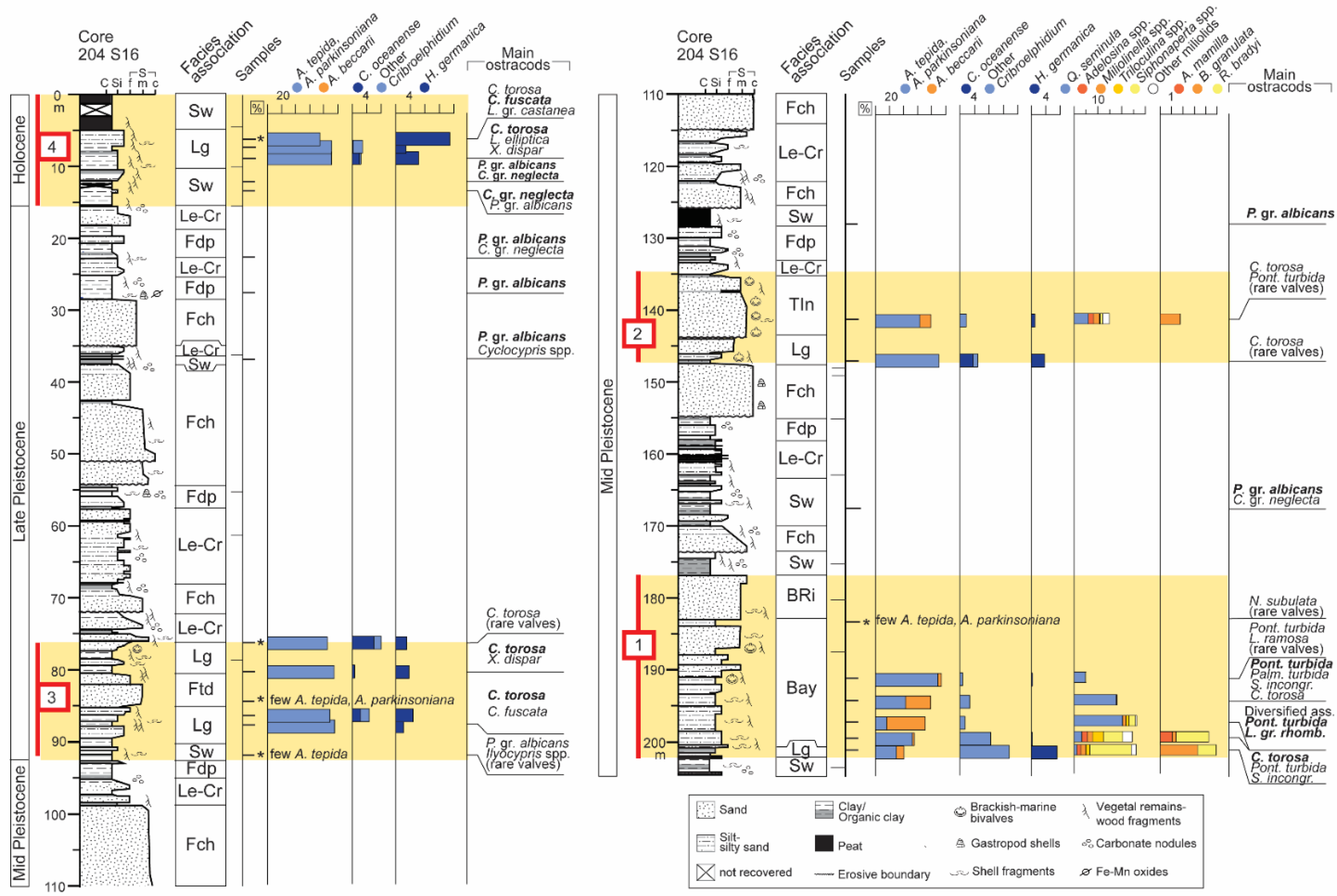
4. Results
4.1. Fluvial-Channel Facies Association (Fch)
4.1.1. Description
4.1.2. Interpretation
4.2. Levee and Crevasse Facies Association (Le-Cr)
4.2.1. Description
4.2.2. Interpretation
4.3. Floodplain Facies Association (Fdp)
4.3.1. Description
4.3.2. Interpretation
4.4. Swamp Facies Association (Sw)
4.4.1. Description
4.4.2. Interpretation
4.5. Lagoon Facies Association (Lg)
4.5.1. Description
4.5.2. Interpretation
4.6. Flood-Tidal Delta Facies Association (Ftd)
4.6.1. Description
4.6.2. Interpretation
4.7. Tidal Inlet Facies Association (TIn)
4.7.1. Description
4.7.2. Interpretation
4.8. Beach Ridge Facies Association (BRi)
4.8.1. Description
4.8.2. Interpretation
4.9. Bay Facies Association (Bay)
4.9.1. Description
4.9.2. Interpretation
5. Discussion
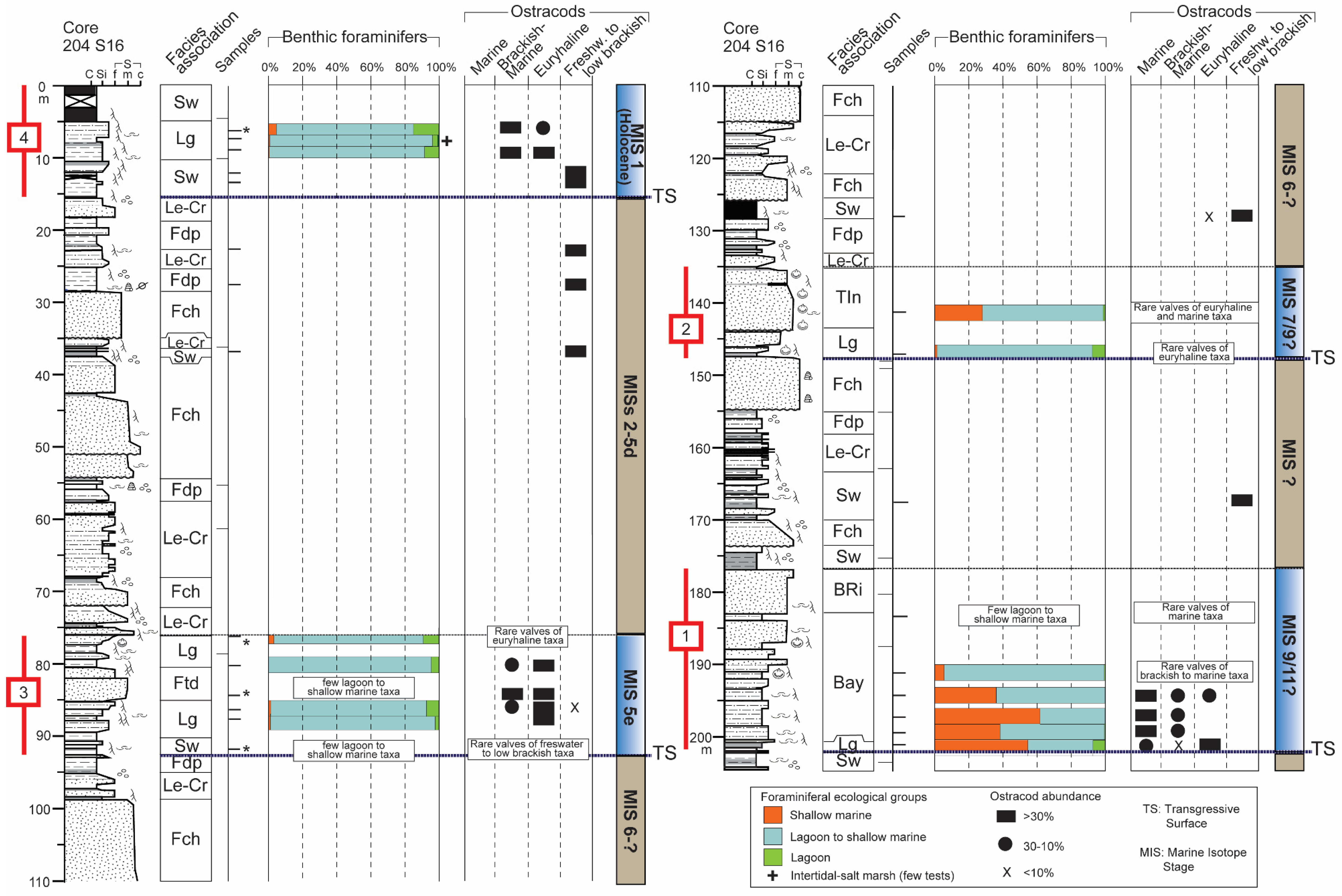
5.1. Core Stratigraphy and Bio-Sedimentary Trends
5.2. Cyclic Sedimentation during the Middle Pleistocene–Holocene Period: Facies Patterns and Controlling Factors
6. Conclusions
- The long-term bio-sedimentary record of the Po coastal plain reconstructed over a significant portion (last ~300–400 kyrs) of the post-Middle Pleistocene Revolution interval, confirms the pivotal role of glacio-eustatic oscillations on depositional and environmental dynamics.
- A cyclic sedimentation is highlighted by the alternation of tens m-thick alluvial deposits and paralic to shallow-marine units (Units 1–4). These latter are invariably underlined by prominent transgressive surfaces that mark major flooding events in the plain and the abrupt landward shift of depositional environments. Based on previous stratigraphic reconstructions, the two youngest units (Units 3 and 4) are attributed to the Last Interglacial and to the Holocene, respectively. Therefore, Unit 2 ad Unit 1 are chronologically constrained to the Middle Pleistocene.
- The strategic position of core 204 S16, well behind the Holocene maximum marine ingression, and the application of an integrated methodology involving meiofauna ecological groups allow reconstructing distinct stratigraphic patterns for the Midde Pleistocene (lowermost ~100 m) and Late Pleistocene-Holocene (uppermost ~100 m) intervals. The former includes coastal to shallow-marine deposits (Units 1, 2), while the latter is typified by the occurrence of back-barrier sediments (Units 3 and 4).
- The lower portions of Units 1–4 invariably exhibit a retrogradational pattern of facies that tracks the transgression under the forcing of rising sea levels; an evident progradational pattern characterises the upper portions of Units 1, 2 and 4, likely reflecting RSL highstand conditions.
- The meiofauna tracks the deepest conditions within the oldest Unit 1; the development of a bay, reasonably in response to a prominent rise in RSL, is attributable to MIS 11 or MIS 9.
- An overall regressive trend is documented within the Middle Pleistocene-Holocene succession of the SE Po Plain, as progressively fewer marine conditions established during interglacials through time, reasonably in response to a complex interaction between glacio-eustatic dynamics and tectonic evolution of the Po Basin over hundreds of thousands of years.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anthony, E.J.; Marriner, N.; Morhange, C. Human Influence and the Changing Geomorphology of Mediterranean Deltas and Coasts over the Last 6000 Years: From Progradation to Destruction Phase? Earth-Sci. Rev. 2014, 139, 336–361. [Google Scholar] [CrossRef]
- Peeters, J.; Busschers, F.S.; Stouthamer, E. Fluvial Evolution of the Rhine during the Last Interglacial-Glacial Cycle in the Southern North Sea Basin: A Review and Look Forward. Quat. Int. 2015, 357, 176–188. [Google Scholar] [CrossRef]
- Peeters, J.; Busschers, F.S.; Stouthamer, E.; Bosch, J.H.A.; Van den Berg, M.W.; Wallinga, J.; Versendaal, A.J.; Bunnik, F.P.M.; Middelkoop, H. Sedimentary Architecture and Chronostratigraphy of a Late Quaternary Incised-Valley Fill: A Case Study of the Late Middle and Late Pleistocene Rhine System in the Netherlands. Quat. Sci. Rev. 2016, 131, 211–236. [Google Scholar] [CrossRef]
- Sarti, G.; Rossi, V.; Amorosi, A.; Bini, M.; Giacomelli, S.; Pappalardo, M.; Ribecai, C.; Ribolini, A.; Sammartino, I. Climatic Signature of Two Mid-Late Holocene Fluvial Incisions Formed under Sea-Level Highstand Conditions (Pisa Coastal Plain, NW Tuscany, Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 424, 183–195. [Google Scholar] [CrossRef]
- Amorosi, A.; Maselli, V.; Trincardi, F. Onshore to Offshore Anatomy of a Late Quaternary Source-to-Sink System (Po Plain-Adriatic Sea, Italy). Earth-Sci. Rev. 2016, 153, 212–237. [Google Scholar] [CrossRef]
- Antonioli, F.; Anzidei, M.; Amorosi, A.; Lo Presti, V.; Mastronuzzi, G.; Deiana, G.; De Falco, G.; Fontana, A.; Fontolan, G.; Lisco, S.; et al. Sea-Level Rise and Potential Drowning of the Italian Coastal Plains: Flooding Risk Scenarios for 2100. Quat. Sci. Rev. 2017, 158, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Alizad, K.; Hagen, S.C.; Medeiros, S.C.; Bilskie, M.V.; Morris, J.T.; Balthis, L.; Buckel, C.A. Dynamic Responses and Implications to Coastal Wetlands and the Surrounding Regions under Sea Level Rise. PLoS ONE 2018, 13, 1–27. [Google Scholar] [CrossRef]
- Anderson, C.C.; Renaud, F.G.; Hagenlocher, M.; Day, J.W. Assessing Multi-Hazard Vulnerability and Dynamic Coastal Flood Risk in the Mississippi Delta: The Global Delta Risk Index as a Social-Ecological Systems Approach. Water 2021, 13, 577. [Google Scholar] [CrossRef]
- Grottoli, E.; Bertoni, D.; Ciavola, P. Short- and Medium-Term Response to Storms on Three Mediterranean Coarse-Grained Beaches. Geomorphology 2017, 295, 738–748. [Google Scholar] [CrossRef]
- Luppichini, M.; Bini, M.; Paterni, M.; Berton, A.; Merlino, S. A New Beach Topography-Based Method for Shoreline Identification. Water 2020, 12, 3110. [Google Scholar] [CrossRef]
- Melet, A.; Teatini, P.; Le Cozannet, G.; Jamet, C.; Conversi, A.; Benveniste, J.; Almar, R. Earth Observations for Monitoring Marine Coastal Hazards and Their Drivers; Springer: Berlin/Heidelberg, Germany, 2020; Volume 41, ISBN 0123456789. [Google Scholar]
- Salvi, G.; Acquavita, A.; Celio, M.; Ciriaco, S.; Cirilli, S.; Fernetti, M.; Pugliese, N. Ostracod Fauna: Eyewitness to Fifty Years of Anthropic Impact in the Gulf of Trieste. A Potential Key to the Future Evolution of Urban Ecosystems. Sustainability 2020, 12, 6954. [Google Scholar] [CrossRef]
- Leorri, E.; Cearreta, A.; Irabien, M.J.; Yusta, I. Geochemical and Microfaunal Proxies to Assess Environmental Quality Conditions during the Recovery Process of a Heavily Polluted Estuary: The Bilbao Estuary Case (N. Spain). Sci. Total Environ. 2008, 396, 12–27. [Google Scholar] [CrossRef]
- Francescangeli, F.; Armynot du Chatelet, E.; Billon, G.; Trentesaux, A.; Bouchet, V.M.P. Palaeo-Ecological Quality Status Based on Foraminifera of Boulogne-Sur-Mer Harbour (Pas-de-Calais, Northeastern France) over the Last 200 Years. Mar. Environ. Res. 2016, 117, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, G.; Rossi, V.; Ghosh, A.; Vaiani, S.C. Conservation Paleobiology as a Tool to Define Reference Conditions in Naturally Stressed Transitional Settings: Micropaleontological Insights from the Holocene of the Po Coastal Plain (Italy). Water 2020, 12, 3420. [Google Scholar] [CrossRef]
- Pasquetti, F.; Vaselli, O.; Zanchetta, G.; Nisi, B.; Lezzerini, M.; Bini, M.; Mele, D. Sedimentological, Mineralogical and Geochemical Features of Late Quaternary Sediment Profiles from the Southern Tuscany Hg Mercury District (Italy): Evidence for the Presence of Pre-Industrial Mercury and Arsenic Concentrations. Water 2020, 12, 1998. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Frontalini, F.; Francescangeli, F.; Sauriau, P.G.; Geslin, E.; Martins, M.V.A.; Almogi-Labin, A.; Avnaim-Katav, S.; Di Bella, L.; Cearreta, A.; et al. Indicative Value of Benthic Foraminifera for Biomonitoring: Assignment to Ecological Groups of Sensitivity to Total Organic Carbon of Species from European Intertidal Areas and Transitional Waters. Mar. Pollut. Bull. 2021, 164, 112071. [Google Scholar] [CrossRef] [PubMed]
- Schuerch, M.; Spencer, T.; Temmerman, S.; Kirwan, M.L.; Wolff, C.; Lincke, D.; McOwen, C.J.; Pickering, M.D.; Reef, R.; Vafeidis, A.T.; et al. Future Response of Global Coastal Wetlands to Sea-Level Rise. Nature 2018, 561, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Milli, S.; Mancini, M.; Moscatelli, M.; Stigliano, F.; Marini, M.; Cavinato, G.P. From River to Shelf, Anatomy of a High-Frequency Depositional Sequence: The Late Pleistocene to Holocene Tiber Depositional Sequence. Sedimentology 2016, 63, 1886–1928. [Google Scholar] [CrossRef] [Green Version]
- Melis, R.T.; Di Rita, F.; French, C.; Marriner, N.; Montis, F.; Serreli, G.; Sulas, F.; Vacchi, M. 8000 years of Coastal Changes on a Western Mediterranean Island: A Multiproxy Approach from the Posada Plain of Sardinia. Mar. Geol. 2018, 403, 93–108. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Campo, B.; Morelli, A.; Rossi, V.; Scarponi, D.; Hong, W.; Bohacs, K.M.; Drexler, T.M. Global Sea-Level Control on Local Parasequence Architecture from the Holocene Record of the Po Plain, Italy. Mar. Pet. Geol. 2017, 87, 99–111. [Google Scholar] [CrossRef]
- Ronchi, L.; Fontana, A.; Cohen, K.M.; Stouthamer, E. Late Quaternary Landscape Evolution of the Buried Incised Valley of Concordia Sagittaria (Tagliamento River, NE Italy): A Reconstruction of Incision and Transgression. Geomorphology 2021, 373, 107509. [Google Scholar] [CrossRef]
- Rossi, V.; Barbieri, G.; Vaiani, S.C.; Cacciari, M.; Bruno, L.; Campo, B.; Marchesini, M.; Marvelli, S.; Amorosi, A. Millennial-Scale Shifts in Microtidal Ecosystems during the Holocene: Dynamics and Drivers of Change from the Po Plain Coastal Record (NE Italy). J. Quat. Sci. 2021, 36, 961–979. [Google Scholar] [CrossRef]
- Waelbroeck, C.; Labeyrie, L.; Michel, E.; Duplessy, J.C.; McManus, J.F.; Lambeck, K.; Balbon, E.; Labracherie, M. Sea-Level and Deep Water Temperature Changes Derived from Benthic Foraminifera Isotopic Records. Quat. Sci. Rev. 2002, 21, 295–305. [Google Scholar] [CrossRef]
- Antonioli, F.; Dai Pra, G.; Hearty, P.J. I Sedimenti Quaternari Nella Fascia Costiera Della Piana Di Fondi (Lazio Meridionale). Boll. Soc. Geol. Ital. (Ital. J. Geosci.) 1988, 107, 491–501. [Google Scholar]
- Romano, P.; Santo, A.; Voltaggio, M. L’evoluzione Geomorfologica Della Pianura Del Fiume Volturno (Campania) Durante Il Tardo Quaternario (Pleistocene Medio-Superiore—Olocene). Alp. Mediterr. Quat. 1994, 7, 41–55. [Google Scholar]
- Amorosi, A.; Colalongo, M.L.; Fiorini, F.; Fusco, F.; Pasini, G.; Vaiani, S.C.; Sarti, G. Palaeogeographic and Palaeoclimatic Evolution of the Po Plain from 150-Ky Core Records. Glob. Planet. Chang. 2004, 40, 55–78. [Google Scholar] [CrossRef]
- Carboni, M.G.; Bergamin, L.; Di Bella, L.; Esu, D.; Cerone, E.P.; Antonioli, F.; Verrubbi, V. Palaeoenvironmental Reconstruction of Late Quaternary Foraminifera and Molluscs from the ENEA Borehole (Versilian Plain, Tuscany, Italy). Quat. Res. 2010, 74, 265–276. [Google Scholar] [CrossRef]
- Campo, B.; Bruno, L.; Amorosi, A. Basin-Scale Stratigraphic Correlation of Late Pleistocene-Holocene (MIS 5e-MIS 1) Strata across the Rapidly Subsiding Po Basin (Northern Italy). Quat. Sci. Rev. 2020, 237. [Google Scholar] [CrossRef]
- Amorosi, A.; Colalongo, M.L.; Fusco, F.; Pasini, G.; Fiorini, F. Glacio-Eustatic Control of Continental-Shallow Marine Cyclicity from Late Quaternary Deposits of the Southeastern Po Plain, Northern Italy. Quat. Res. 1999, 52, 1–13. [Google Scholar] [CrossRef]
- Amorosi, A.; Forlani, L.; Fusco, F.; Severi, P. Cyclic Patterns of Facies and Pollen Associations from Late Quaternary Deposits in the Subsurface of Bologna. GeoActa 2001, 1, 83–94. [Google Scholar]
- Kent, D.V.; Rio, D.; Massari, F.; Kukla, G.; Lanci, L. Emergence of Venice during the Pleistocene. Quat. Sci. Rev. 2002, 21, 1719–1727. [Google Scholar] [CrossRef] [Green Version]
- Bossio, A.; Ciampo, G.; Dall’Antonia, B. Quaternary Environmental Evolution of the Venice Area Based upon Ostracod Assemblages. Boll. Della Soc. Paleontol. Ital. 2004, 43, 113–122. [Google Scholar]
- Massari, F.; Rio, D.; Serandrei Barbero, R.; Asioli, A.; Capraro, L.; Fornaciari, E.; Vergerio, P.P. The Environment of Venice Area in the Past Two Million Years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 202, 273–308. [Google Scholar] [CrossRef]
- Carminati, E.; Di Donato, G. Separating Natural and Anthropogenic Vertical Movements in Fast Subsiding Areas: The Po Plain (N. Italy) Case. Geophys. Res. Lett. 1999, 26, 2291–2294. [Google Scholar] [CrossRef]
- Ghielmi, M.; Minervini, M.; Nini, C.; Rogledi, S.; Rossi, M. Late Miocene-Middle Pleistocene Sequences in the Po Plain—Northern Adriatic Sea (Italy): The Stratigraphic Record of Modification Phases Affecting a Complex Foreland Basin. Mar. Pet. Geol. 2013, 42, 50–81. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Campo, B.; Costagli, B.; Hong, W.; Picotti, V.; Vaiani, S.C. Deformation Patterns of Upper Quaternary Strata and Their Relation to Active Tectonics, Po Basin, Italy. Sedimentology 2021, 68, 402–424. [Google Scholar] [CrossRef]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene Stack of 57 Globally Distributed Benthic δ 18O Records. Paleoceanography 2005, 20, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Burrato, P.; Ciucci, F.; Valensise, G. An Inventory of River Anomalies in the Po Plain, Northern Italy: Evidence for Active Blind Thrust Faulting. Ann. Geophys. 2003, 46, 865–882. [Google Scholar]
- Bondesan, M.; Favero, V.; Viñals, M.J. New Evidence on the Evolution of the Po-Delta Coastal Plain during the Holocene. Quat. Int. 1995, 29–30, 105–110. [Google Scholar] [CrossRef]
- Correggiari, A.; Cattaneo, A.; Trincardi, F. Depositional Patterns in the Late Holocene Po Delta System. SEPM Spec. Publ. 2005, 83, 365–392. [Google Scholar] [CrossRef] [Green Version]
- Carminati, E.; Doglioni, C. Alps vs. Apennines: The Paradigm of a Tectonically Asymmetric Earth. Earth-Sci. Rev. 2012, 112, 67–96. [Google Scholar] [CrossRef]
- Pieri, M.; Groppi, G. Subsurface Geological Structure of the Po Plain. Italy Pubbl. 1981, 441, 1–11. [Google Scholar]
- Boccaletti, M.; Corti, G.; Martelli, L. Recent and Active Tectonics of the External Zone of the Northern Apennines (Italy). Int. J. Earth Sci. 2011, 100, 1331–1348. [Google Scholar] [CrossRef]
- Carminati, E.; Doglioni, C.; Scrocca, D. Apennines Subduction-Related Subsidence of Venice (Italy). Geophys. Res. Lett. 2003, 30, 1–4. [Google Scholar] [CrossRef]
- Bruno, L.; Campo, B.; Costagli, B.; Stouthamer, E.; Teatini, P.; Zoccarato, C.; Amorosi, A. Factors Controlling Natural Subsidence in the Po Plain. Proc. Int. Assoc. Hydrol. Sci. 2020, 382, 285–290. [Google Scholar] [CrossRef]
- Amadori, C.; Toscani, G.; Di Giulio, A.; Maesano, F.E.; D’Ambrogi, C.; Ghielmi, M.; Fantoni, R. From Cylindrical to Non-Cylindrical Foreland Basin: Pliocene–Pleistocene Evolution of the Po Plain–Northern Adriatic Basin (Italy). Basin Res. 2019, 31, 991–1015. [Google Scholar] [CrossRef] [Green Version]
- Mitchum, J.R.M.; Vail, P.R.; III, T.S. Seismic stratigraphy and global changes of sealevel, Part 2: The depositional sequence as a basic unit for stratigraphic analysis. In American Association of Petroleum Geologists Memoir 26; Payton, C.E., Ed.; American Association of Petroleum Geologists: Tulsa, OR, USA, 1977; pp. 53–62. [Google Scholar]
- Emilia-Romagna, R. ENI-AGIP Riserve Idriche Sotterranee Della Regione Emilia-Romagna; S.EL.CA: Firenze, Italy, 1998. [Google Scholar]
- Dondi, L.; Mostardini, F.; Rizzini, A. Evoluzione sedimentaria e paleogeografia nella Pianura Padana. In Guida alla Geologia del Margine Appenninico-Padano; Cremonini, G., Ricci Lucchi, F., Eds.; IBS: Hong Kong, China, 1982; pp. 47–58. [Google Scholar]
- Ghielmi, M.; Minervini, M.; Nini, C.; Rogledi, S.; Rossi, M.; Vignolo, A. Sedimentary and Tectonic Evolution in the Eastern Po-Plain and Northern Adriatic Sea Area from Messinian to Middle Pleistocene (Italy). Rend. Lincei 2010, 21, 131–166. [Google Scholar] [CrossRef]
- Amorosi, A.; Colalongo, M.L. The linkage between alluvial and coeval nearshoremarine succession: Evidence from the late Quaternary Record of the Po River Plain, Italy. In Fluvial Sedimentology VII. IAS Special Publication; Blum, M.D., Marriott, S., Leclair, S.F., Eds.; Wiley Online: Oxford, UK, 2005; pp. 257–275. [Google Scholar]
- Amorosi, A.; Colalongo, M.L.; Dinelli, E.; Lucchini, F.; Vaiani, S.C. Cyclic Variations in Sediment Provenance from Late Pleistocene Deposits of the Eastern Po Plain, Italy. Spec. Pap. Geol. Soc. Am. 2007, 420, 13–24. [Google Scholar] [CrossRef]
- Amorosi, A.; Pavesi, M.; Ricci Lucchi, M.; Sarti, G.; Piccin, A. Climatic Signature of Cyclic Fluvial Architecture from the Quaternary of the Central Po Plain, Italy. Sediment. Geol. 2008, 209, 58–68. [Google Scholar] [CrossRef]
- Bondesan, M.; Cibin, U.; Colalongo, M.L.; Pugliese, N.; Stefani, M.; Tsakiridis, E.; Vaiani, S.C.; Vincenzi, S. Benthic Communities and Sedimentary Facies Recording Late Quaternary Environmental Fluctuations in a Po Delta Subsurface Succession (Northern Italy). In Proceedings of the Second and Third Italian Meeting of Environmental Micropaleontology, Urbino, Italy, 4–6 June 2006; Coccioni, R., Lirer, F., Marsili, A., Eds.; The Grzybowski Foundation Special Publication 11: Krakov, Poland, 2006; pp. 21–31. [Google Scholar]
- Carta Geologica d’Italia Scala 1:50,000. Available online: https://www.isprambiente.gov.it/Media/carg/emilia.html (accessed on 14 August 2021).
- Emilia-Romagna Geological, Seismic and Soil Survey, Geological Cartography Webgis. Available online: Mappegis.regione.emilia-romagna.it/gstatico/documenti/Masterlog/204080P517X.pdf (accessed on 14 August 2021).
- Amorosi, A.; Dinelli, E.; Rossi, V.; Vaiani, S.C.; Sacchetto, M. Late Quaternary Palaeoenvironmental Evolution of the Adriatic Coastal Plain and the Onset of Po River Delta. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 268, 80–90. [Google Scholar] [CrossRef]
- Ellis, B.F.; Messina, A.R. Catalogue of Foraminifera; Micropaleontology Press: New York, NY, USA, 1940. [Google Scholar]
- Ellis, B.F.; Messina, A.R. Catalogue of Ostracoda; The American Museum of Natural History Special Publications: New York, NY, USA, 1952. [Google Scholar]
- Bonaduce, G.; Ciampo, G.; Masoli, M. Distribution of Ostracoda in the Adriatic Sea; Olschki, L.S., Ed.; Pubblicazioni della Stazione Zoologica di Napoli; The Micropaleontology Project., Inc.: Napoli, Italy, 1975; Volume 40. [Google Scholar]
- Jorissen, F.J. Benthic Foraminifera from the Adriatic Sea; Principles of Phenotypic Variation; Utrecht Micropaleontological Bulletins: Utrecht, The Netherlands, 1988; Volume 37. [Google Scholar]
- Athersuch, J.; Horne, D.J.; Whittaker, J.E. Marine and Brackish Water Ostracods; Kermack, D.M., Barnes, R.S.K., Eds.; Synopses of the British Fauna (New Series) 43 Brill, E.J.: Leiden, The Netherlands, 1989. [Google Scholar]
- Albani, A.D.; Serandrei Barbero, R. I Foraminiferi Della Laguna e Del Golfo Di Venezia. Mem. Di Sci. Geol. 1990, 42, 271–341. [Google Scholar]
- Henderson, P.A. Freshwater Ostracods; Kermack, D.M., Barnes, R.S.K., Eds.; Synopses of the British Fauna (New Series) 42 Brill, E.J.: Leiden, The Netherlands, 1990. [Google Scholar]
- Sgarrella, F.; Moncharmont Zei, M. Benthic Foraminifera of the Gulf of Naples (Italy): Systematics and Autoecology. Boll. Della Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Mazzini, I.; Anadon, P.; Barbieri, M.; Castorina, F.; Ferreli, L.; Gliozzi, E.; Mola, M.; Vittori, E. Late Quaternary Sea-Level Changes along the Tyrrhenian Coast near Orbetello (Tuscany, Central Italy): Palaeoenvironmental Reconstruction Using Ostracods. Mar. Micropaleontol. 1999, 37, 289–311. [Google Scholar] [CrossRef]
- Fiorini, F.; Vaiani, S.C. Benthic Foraminifers and Transgressive-Regressive Cycles in the Late Quaternary Subsurface Sediments of the Po Plain near Ravenna (Northern Italy). Boll. Della Soc. Paleontol. Ital. 2001, 40, 357–403. [Google Scholar]
- Barbieri, G.; Vaiani, S.C. Benthic Foraminifera or Ostracoda? Comparing the Accuracy of Palaeoenvironmental Indicators from a Pleistocene Lagoon of the Romagna Coastal Plain (Italy). J. Micropalaeontol. 2018, 37, 203–230. [Google Scholar] [CrossRef]
- Coccioni, R. Benthic foraminifera as bioindicators of heavy metal pollution—A case study from the Goro Lagoon (Italy). In Environmental Micropaleontology: The Application of Microfossils to Environmental Geology; Martin, R.E., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2000; pp. 71–103. [Google Scholar]
- Debenay, J.P.; Guillou, J.J. Ecological Transitions Indicated by Foraminiferal Assemblages in Paralic Environments. Estuaries 2002, 25, 1107–1120. [Google Scholar] [CrossRef]
- Melis, R.; Covelli, S. Distribution and Morphological Abnormalities of Recent Foraminifera in the Marano and Grado Lagoon (North Adriatic Sea, Italy). Mediterr. Mar. Sci. 2013, 14, 432–450. [Google Scholar] [CrossRef] [Green Version]
- Mazzini, I.; Rossi, V.; Da Prato, S.; Ruscito, V. Ostracods in archaeological sites along the Mediterranean coastlines: Three case studies from the Italian peninsula. In The Archaeological and Forensic Applications of Microfossils: A Deeper Understanding of Human History; Geological Society of London: London, UK, 2017; pp. 121–142. [Google Scholar]
- Frenzel, P.; Boomer, I. The Use of Ostracods from Marginal Marine, Brackish Waters as Bioindicators of Modern and Quaternary Environmental Change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 225, 68–92. [Google Scholar] [CrossRef]
- Aiello, G.; Amato, V.; Barra, D.; Caporaso, L.; Caruso, T.; Giaccio, B.; Parisi, R.; Rossi, A. Late Quaternary Benthic Foraminiferal and Ostracod Response to Palaeoenvironmental Changes in a Mediterranean Coastal Area, Port of Salerno, Tyrrhenian Sea. Reg. Stud. Mar. Sci. 2020, 40, 101498. [Google Scholar] [CrossRef]
- D’Orefice, M.; Bellotti, P.; Bertini, A.; Calderoni, G.; Neri, P.C.; Di Bella, L.; Fiorenza, D.; Foresi, L.M.; Louvari, M.A.; Rainone, L.; et al. Holocene Evolution of the Burano Paleo-Lagoon (Southern Tuscany, Italy). Water 2020, 12, 1007. [Google Scholar] [CrossRef] [Green Version]
- Bruno, L.; Bohacs, K.M.; Campo, B.; Drexler, T.M.; Rossi, V.; Sammartino, I.; Scarponi, D.; Hong, W.; Amorosi, A. Early Holocene Transgressive Palaeogeography in the Po Coastal Plain (Northern Italy). Sedimentology 2017, 64, 1792–1816. [Google Scholar] [CrossRef]
- Campo, B.; Amorosi, A.; Vaiani, S.C. Sequence Stratigraphy and Late Quaternary Paleoenvironmental Evolution of the Northern Adriatic Coastal Plain (Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 466, 265–278. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Campo, B.; Morelli, A. The Value of Pocket Penetration Tests for the High-Resolution Palaeosol Stratigraphy of Late Quaternary Deposits. Geol. J. 2015, 50, 670–682. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Cleveland, D.M.; Morelli, A.; Hong, W. Paleosols and Associated Channel-Belt Sand Bodies from a Continuously Subsiding Late Quaternary System (Po Basin, Italy): New Insights into Continental Sequence Stratigraphy. Bull. Geol. Soc. Am. 2017, 129, 449–463. [Google Scholar] [CrossRef]
- Meisch, C. Freshwater Ostracoda of Western and Central Europe; Süsswasserfauna von Mitteleuropa; Spektrum Akademischer Verlag: Heidelberg/Berlin, Germany, 2000; Volume 3. [Google Scholar]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Amorosi, A.; Bruno, L.; Cacciari, M.; Campo, B.; Rossi, V. Tracing Marine Flooding Surface Equivalents across Freshwater Peats and Other Wetland Deposits by Integrated Sedimentological and Pollen Data. Int. J. Coal Geol. 2021, 246, 103830. [Google Scholar] [CrossRef]
- Pint, A.; Frenzel, P. Ostracod Fauna Associated with Cyprideis torosa—An Overview. J. Micropalaeontol. 2017, 36, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Tibert, N.E.; Walker, L.J.; Patterson, W.P.; Hubeny, J.B.; Jones, E.; Cooper, O.R. A Centennial Record of Paleosalinity Change in the Tidal Reaches of the Potomac and Rappahannock Rivers, Tributaries to Chesapeake Bay. Va. J. Sci. 2012, 63, 111–128. [Google Scholar]
- Horne, D.J.; Benardout, G.; Whittaker, J.E. Cyprideis torosa (Jones, 1850) in Its Type Area and Stratigraphical Context: Potential for Mapping the Freshwater/Estuarine Boundaries of the Thames–Medway River System in the MIS 9 and MIS 11 Interglacials. J. Micropalaeontol. 2017, 36, 127–135. [Google Scholar] [CrossRef]
- FitzGerald, D.; Buynevich, I.; Hein, C. Morphodynamics and Facies Architecture of Tidal Inlets and Tidal Deltas. In Principles of Tidal Sedimentology; Davis, R.A., Dalrymple, R.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; ISSN 9789400701236. [Google Scholar]
- Zecchin, M.; Tosi, L.; Caffau, M.; Baradello, L.; Donnici, S. Sequence Stratigraphic Significance of Tidal Channel Systems in a Shallow Lagoon (Venice, Italy). Holocene 2014, 24, 646–658. [Google Scholar] [CrossRef]
- Aiello, G.; Barra, D.; Coppa, M.G.; Valente, A.; Zeni, F. Recent Infralittoral Foraminiferida and Ostracoda from the Porto Cesareo Lagoon (Ionian Sea, Mediterranean). Boll. Della Soc. Paleontol. Ital. 2006, 45, 1–14. [Google Scholar]
- Tsourou, T. Composition and Distribution of Recent Marine Ostracod Assemblages in the Bottom Sediments of Central Aegean Sea (SE Andros Island, Greece). Int. Rev. Hydrobiol. 2012, 97, 276–300. [Google Scholar] [CrossRef]
- Frenzel, P.; Keyser, D.; Viehberg, F.A. An Illustrated Key and (Palaeo)Ecological Primer for Postglacial to Recent Ostracoda (Crustacea) of the Baltic Sea. Boreas 2010, 39, 567–575. [Google Scholar] [CrossRef]
- Serandrei Barbero, R.; Albani, A.D.; Bonardi, M. Ancient and Modern Salt Marshes in the Lagoon of Venice. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 202, 229–244. [Google Scholar] [CrossRef]
- Stefani, M.; Vincenzi, S. The Interplay of Eustasy, Climate and Human Activity in the Late Quaternary Depositional Evolution and Sedimentary Architecture of the Po Delta System. Mar. Geol. 2005, 222–223, 19–48. [Google Scholar] [CrossRef]
- Raymo, M.E.; Mitrovica, J.X. Collapse of Polar Ice Sheets during the Stage 11 Interglacial. Nature 2012, 483, 453–456. [Google Scholar] [CrossRef] [Green Version]
- Spratt, R.M.; Lisiecki, L.E. A Late Pleistocene Sea Level Stack. Clim. Past 2016, 12, 1079–1092. [Google Scholar] [CrossRef] [Green Version]
- Antonioli, F. Sea Level Change in Western-Central Mediterranean since 300 Kyr: Comparing Global Sea Level Curves with Observed Data. Alp. Mediterr. Quat. 2012, 25, 15–23. [Google Scholar]
- Rovere, A.; Raymo, M.E.; Vacchi, M.; Lorscheid, T.; Stocchi, P.; Gómez-Pujol, L.; Harris, D.L.; Casella, E.; O’Leary, M.J.; Hearty, P.J. The Analysis of Last Interglacial (MIS 5e) Relative Sea-Level Indicators: Reconstructing Sea-Level in a Warmer World. Earth-Sci. Rev. 2016, 159, 404–427. [Google Scholar] [CrossRef] [Green Version]
- Bini, M.; Zanchetta, G.; Drysdale, R.N.; Giaccio, B.; Stocchi, P.; Vacchi, M.; Hellstrom, J.C.; Couchoud, I.; Monaco, L.; Ratti, A.; et al. An End to the Last Interglacial Highstand before 120 Ka: Relative Sea-Level Evidence from Infreschi Cave (Southern Italy). Quat. Sci. Rev. 2020, 250, 106658. [Google Scholar] [CrossRef]
- Rasmussen, T.L.; Thomsen, E. Foraminifera and paleoenvironment of the Plio-Pleistocene Kallithea Bay Section, Rhodes, Greece: Evidence for cyclic sedimentation and shallow-water sapropels. In Lagoon to Deep-Water Formaminifera and Ostracods from the Plio-Pleistocene Kallithea Bay Section, Rhodes, Greece; Rasmussen, T.L., Hastrup, A., Thomsen, E., Eds.; Cushman Foundation for Foraminiferal Research: Fredericksburg, VR, USA, 2005; Volume 39, pp. 15–51. ISBN 9781970168341. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, V.; Amorosi, A.; Barbieri, G.; Vaiani, S.C.; Germano, M.; Campo, B. A Long-Term Record of Quaternary Facies Patterns and Palaeonvironmental Trends from the Po Plain (NE Italy) as Revealed by Bio-Sedimentary Data. Geosciences 2021, 11, 401. https://doi.org/10.3390/geosciences11100401
Rossi V, Amorosi A, Barbieri G, Vaiani SC, Germano M, Campo B. A Long-Term Record of Quaternary Facies Patterns and Palaeonvironmental Trends from the Po Plain (NE Italy) as Revealed by Bio-Sedimentary Data. Geosciences. 2021; 11(10):401. https://doi.org/10.3390/geosciences11100401
Chicago/Turabian StyleRossi, Veronica, Alessandro Amorosi, Giulia Barbieri, Stefano Claudio Vaiani, Matteo Germano, and Bruno Campo. 2021. "A Long-Term Record of Quaternary Facies Patterns and Palaeonvironmental Trends from the Po Plain (NE Italy) as Revealed by Bio-Sedimentary Data" Geosciences 11, no. 10: 401. https://doi.org/10.3390/geosciences11100401
APA StyleRossi, V., Amorosi, A., Barbieri, G., Vaiani, S. C., Germano, M., & Campo, B. (2021). A Long-Term Record of Quaternary Facies Patterns and Palaeonvironmental Trends from the Po Plain (NE Italy) as Revealed by Bio-Sedimentary Data. Geosciences, 11(10), 401. https://doi.org/10.3390/geosciences11100401