
Effects of Stem Density and Reynolds Number on Fine Sediment Interception by Emergent Vegetation


Abstract
:1. Introduction
1.1. Background
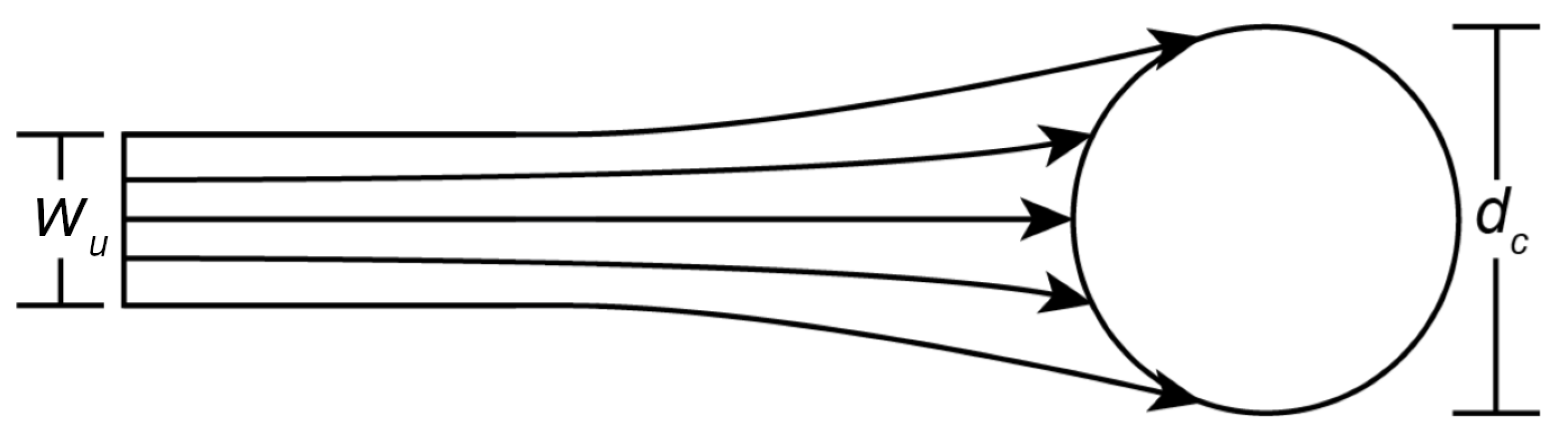
1.2. Particle Capture Models

2. Materials and Methods
2.1. Experimental Methods
2.1.1. Materials
- A flat-bedded array positioned flush with the neighboring channel bed, containing vertical, emergent collector stems.
- Two battery-operated peristaltic pumps (Cole-Parmer, Vernon Hills, IL) used to sample suspended particle concentration via three hose inlets each (inside diameter = 3.1 mm), which were suspended at a range of heights from the channel bed (5; 14; 27 cm); these were positioned 50 cm upstream and downstream of the test section and were sampled at a flow rate equivalent to the mean flume velocity.
- Sediment traps (n = 9) with 2.5 cm circular openings flush with the bed and collection filters (Whatman GF/F) on perforated filter holders recessed in a 5 cm deep cylindrical cavity (trap Reynolds number and aspect ratio chosen to minimize bias [46]), interspersed among the collectors in a grid-like pattern (Figure 3).
2.1.2. Suspended Particle Concentration Analysis
2.1.3. Estimating Collector-Induced Turbulence
2.1.4. Biofilm Growth
2.2. Sediment Transport and Particle Capture Model
2.2.1. Model Derivation
2.2.2. Model Execution
3. Results
3.1. Particle Capture
3.2. Turbulence
3.3. Biofilm Effects
3.4. Comparison to Previous Models of Capture Efficiency
4. Discussion
4.1. Inferred Mechanisms of Effect for Collector Density and Reynolds Number
4.2. Relative Importance of Biofilm
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| capture efficiency, dimensionless | |
| upstream width of streamlines that intersect a collector, cm | |
| collector diameter, cm | |
| effective capture efficiency (ECE), dimensionless | |
| probability of particle retention, dimensionless | |
| u | flow velocity, cm/s |
| collector Reynolds number, dimensionless | |
| kinematic viscosity, / | |
| particle diameter, | |
| collector solid volume fraction, dimensionless | |
| R | particle-collector diameter ratio, dimensionless |
| Peclet number, dimensionless | |
| D | diffusion coefficient, / |
| depth-averaged suspended sediment concentration, / | |
| t | time, s |
| h | water depth, cm |
| z | height above the bed, cm |
| suspended sediment concentration at height z, / | |
| concentration time-decay due to settling, s | |
| settling velocity, cm/s | |
| constant relating near-bed concentration to depth-averaged concentration, dimensionless | |
| entrainment rate, dimensionless | |
| concentration time-decay due to capture, s | |
| collector height density, / | |
| number of collectors, # | |
| V | test-section water volume, |
| sediment mass settled in test section, g | |
| sediment mass suspended at beginning of experiment, g | |
| T | total duration of experiment, s |
| k | total concentration time decay, s |
| background concentration time-decay, s | |
| power-law coefficient for parameter i, dimensionless |
Appendix A. Secondary Methodology
Appendix A.1. Particle Size

Appendix A.2. Flume Volume

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Estimated Volume (m3) | R2 |
|---|---|---|
| Linear | 2.4309 | 0.80 |
| Quadratic | 2.4277 | 0.90 |
| Cubic | 2.4373 | 0.94 |
| 4th-order | 2.4294 | 0.96 |

Appendix A.3. Concentration Profile

Appendix A.4. Heteroscedasticity Reduction

Appendix A.5. Results of Experimental Runs

References
- Thorne, K.; MacDonald, G.; Guntenspergen, G.; Ambrose, R.; Buffington, K.; Dugger, B.; Freeman, C.; Janousek, C.; Brown, L.; Rosencranz, J.; et al. US Pacific coastal wetland resilience and vulnerability to sea-level rise. Sci. Adv. 2018, 4, eaao3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowski, K.L.; Törnqvist, T.E.; Fernandes, A.M. Vulnerability of Louisiana’s coastal wetlands to present-day rates of relative sea-level rise. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirwan, M.L.; Guntenspergen, G.R.; D’Alpaos, A.; Morris, J.T.; Mudd, S.M.; Temmerman, S. Limits on the adaptability of coastal marshes to rising sea level. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef] [Green Version]
- Fagherazzi, S.; Kirwan, M.L.; Mudd, S.M.; Guntenspergen, G.R.; Temmerman, S.; D’Alpaos, A.; Van De Koppel, J.; Rybczyk, J.M.; Reyes, E.; Craft, C.; et al. Numerical models of salt marsh evolution: Ecological, geomorphic, and climatic factors. Rev. Geophys. 2012, 50. [Google Scholar] [CrossRef]
- Goodwin, T.H.; Young, A.R.; Holmes, M.G.; Old, G.H.; Hewitt, N.; Leeks, G.J.; Packman, J.C.; Smith, B.P. The temporal and spatial variability of sediment transport and yields within the Bradford Beck catchment, West Yorkshire. Sci. Total Environ. 2003, 314, 475–494. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Murray, A.B. A coupled geomorphic and ecological model of tidal marsh evolution. Proc. Natl. Acad. Sci. USA 2007, 104, 6118–6122. [Google Scholar] [CrossRef] [Green Version]
- Stumpf, R.P. The process of sedimentation on the surface of a salt marsh. Estuar. Coast. Shelf Sci. 1983, 17, 495–508. [Google Scholar] [CrossRef]
- Leonard, L.A.; Luther, M.E. Flow hydrodynamics in tidal marsh canopies. Limnol. Oceanogr. 1995, 40, 1474–1484. [Google Scholar] [CrossRef]
- Mudd, S.M.; D’Alpaos, A.; Morris, J.T. How does vegetation affect sedimentation on tidal marshes? Investigating particle capture and hydrodynamic controls on biologically mediated sedimentation. J. Geophys. Res. Earth Surf. 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Fauria, K.E.; Kerwin, R.E.; Nover, D.; Schladow, S.G. Suspended particle capture by synthetic vegetation in a laboratory flume. Water Resour. Res. 2015, 51, 9112–9126. [Google Scholar] [CrossRef]
- Spielman, L.A. Particle capture from low-speed laminar flows. Annu. Rev. Fluid Mech. 1977, 9, 297–319. [Google Scholar] [CrossRef]
- Rubenstein, D.I.; Koehl, M.A. The mechanisms of filter feeding: Some theoretical considerations. Am. Nat. 1977, 111, 981–994. [Google Scholar] [CrossRef]
- Christiansen, T.; Wiberg, P.; Milligan, T. Flow and sediment transport on a tidal salt marsh surface. Estuar. Coast. Shelf Sci. 2000, 50, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P. Turbulence effects on the settling of suspended particles. J. Sediment. Res. 1993, 63, 835–838. [Google Scholar]
- Jacobs, C.N.; Merchant, W.; Jendrassak, M.; Limpasuvan, V.; Gurka, R.; Hackett, E.E. Flow scales of influence on the settling velocities of particles with varying characteristics. PLoS ONE 2016, 11, e0159645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lam, K.M.; Lu, Y. Settling velocity of fine heavy particles in turbulent open channel flow. Phys. Fluids 2018, 30, 095106. [Google Scholar] [CrossRef]
- Yager, E.; Schmeeckle, M. The influence of vegetation on turbulence and bed load transport. J. Geophys. Res. Earth Surf. 2013, 118, 1585–1601. [Google Scholar] [CrossRef]
- Yang, J.Q.; Nepf, H.M. Impact of vegetation on bed load transport rate and bedform characteristics. Water Resour. Res. 2019, 55, 6109–6124. [Google Scholar] [CrossRef]
- Jordanova, A.A.; James, C. Experimental study of bed load transport through emergent vegetation. J. Hydraul. Eng. 2003, 129, 474–478. [Google Scholar] [CrossRef]
- D’Alpaos, A.; Lanzoni, S.; Marani, M.; Rinaldo, A. Landscape evolution in tidal embayments: Modeling the interplay of erosion, sedimentation, and vegetation dynamics. J. Geophys. Res. Earth Surf. 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Coco, G. Turbulence as the main driver of resuspension in oscillatory flow through vegetation. J. Geophys. Res. Earth Surf. 2018, 123, 891–904. [Google Scholar] [CrossRef]
- Dietrich, W.E. Settling velocity of natural particles. Water Resour. Res. 1982, 18, 1615–1626. [Google Scholar] [CrossRef]
- Yang, S.; Li, H.; Ysebaert, T.; Bouma, T.; Zhang, W.; Wang, Y.; Li, P.; Li, M.; Ding, P. Spatial and temporal variations in sediment grain size in tidal wetlands, Yangtze Delta: On the role of physical and biotic controls. Estuar. Coast. Shelf Sci. 2008, 77, 657–671. [Google Scholar] [CrossRef]
- Morris, J.T.; Sundareshwar, P.; Nietch, C.T.; Kjerfve, B.; Cahoon, D.R. Responses of coastal wetlands to rising sea level. Ecology 2002, 83, 2869–2877. [Google Scholar] [CrossRef]
- Braskerud, B. The influence of vegetation on sedimentation and resuspension of soil particles in small constructed wetlands. J. Environ. Qual. 2001, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Gleason, M.L.; Elmer, D.A.; Pien, N.C.; Fisher, J.S. Effects of stem density upon sediment retention by salt marsh cord grass, Spartina alterniflora Loisel. Estuaries 1979, 2, 271–273. [Google Scholar] [CrossRef]
- Tinoco, R.O.; Coco, G. A laboratory study on sediment resuspension within arrays of rigid cylinders. Adv. Water Resour. 2016, 92, 1–9. [Google Scholar] [CrossRef]
- Palmer, M.R.; Nepf, H.M.; Pettersson, T.J.; Ackerman, J.D. Observations of particle capture on a cylindrical collector: Implications for particle accumulation and removal in aquatic systems. Limnol. Oceanogr. 2004, 49, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Purich, A. The Capture of Suspended Particles by Aquatic Vegetation. Ph.D. Thesis, University of Western Australia, Perth, Australia, 2006. [Google Scholar]
- Tanino, Y.; Nepf, H.M. Laboratory investigation of lateral dispersion within dense arrays of randomly distributed cylinders at transitional Reynolds number. Phys. Fluids 2009, 21, 046603. [Google Scholar] [CrossRef]
- Wu, L.; Gao, B.; Muñoz-Carpena, R.; Pachepsky, Y.A. Single collector attachment efficiency of colloid capture by a cylindrical collector in laminar overland flow. Environ. Sci. Technol. 2012, 46, 8878–8886. [Google Scholar] [CrossRef]
- Peruzzo, P.; Defina, A.; Nepf, H. Capillary trapping of buoyant particles within regions of emergent vegetation. Water Resour. Res. 2012, 48. [Google Scholar] [CrossRef]
- Fuchs, N.A. The Mechanics of Aerosols; Pergamon Press: London, UK, 1964. [Google Scholar]
- Durham, W.M.; Climent, E.; Barry, M.; De Lillo, F.; Boffetta, G.; Cencini, M.; Stocker, R. Turbulence drives microscale patches of motile phytoplankton. Nat. Commun. 2013, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nepf, H.M. Drag, turbulence, and diffusion in flow through emergent vegetation. Water Resour. Res. 1999, 35, 479–489. [Google Scholar] [CrossRef]
- Wu, L.; Muñoz-Carpena, R.; Gao, B.; Yang, W.; Pachepsky, Y.A. Colloid filtration in surface dense vegetation: Experimental results and theoretical predictions. Environ. Sci. Technol. 2014, 48, 3883–3890. [Google Scholar] [CrossRef] [PubMed]
- Lamb, H. Hydrodynamics; University Press: Cambridge, UK, 1932. [Google Scholar]
- Langmuir, I.; Rodebush, W.; Lamer, V. Filtration of Aerosols and Development of Filter Materials; OSRD-865; Office of Scientific Research and Development: Washington, DC, USA, 1942. [Google Scholar]
- Wu, L.; Gao, B.; Muñoz-Carpena, R. Experimental Analysis of Colloid Capture by a Cylindrical Collector in Laminar Overland Flow. Environ. Sci. Technol. 2011, 45, 7777–7784. [Google Scholar] [CrossRef]
- Pang, S.; Zhang, S.; Lv, X.; Han, B.; Liu, K.; Qiu, C.; Wang, C.; Wang, P.; Toland, H.; He, Z. Characterization of bacterial community in biofilm and sediments of wetlands dominated by aquatic macrophytes. Ecol. Eng. 2016, 97, 242–250. [Google Scholar] [CrossRef]
- Crump, B.C.; Koch, E.W. Attached bacterial populations shared by four species of aquatic angiosperms. Appl. Environ. Microbiol. 2008, 74, 5948–5957. [Google Scholar] [CrossRef] [Green Version]
- DiCesare, E.W.; Hargreaves, B.; Jellison, K. Biofilm roughness determines Cryptosporidium parvum retention in environmental biofilms. Appl. Environ. Microbiol. 2012, 78, 4187–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searcy, K.E.; Packman, A.I.; Atwill, E.R.; Harter, T. Capture and retention of Cryptosporidium parvum oocysts by Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2006, 72, 6242–6247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, S.; Wingenroth, J.; Larsen, L. A functional form for particle interception in vegetated environments. Geosciences 2021. (in review). [Google Scholar]
- Discflo Disc Pumps: Installation, Operation, and Maintenance Manual; Discflo: Santee, CA, USA, 2003.
- Butman, C.A. Sediment trap biases in turbulent flows: Results from a laboratory flume study. J. Mar. Res. 1986, 44, 645–693. [Google Scholar] [CrossRef]
- Müller, M.; De Cesare, G.; Schleiss, A.J. Experiments on the effect of inflow and outflow sequences on suspended sediment exchange rates. Int. J. Sediment Res. 2017, 32, 155–170. [Google Scholar] [CrossRef]
- Jenzer Althaus, J.M.; Cesare, G.D.; Schleiss, A.J. Sediment evacuation from reservoirs through intakes by jet-induced flow. J. Hydraul. Eng. 2015, 141, 04014078. [Google Scholar] [CrossRef]
- Redding, T.; Devito, K. Particle densities of wetland soils in northern Alberta, Canada. Can. J. Soil Sci. 2006, 86, 57–60. [Google Scholar] [CrossRef]
- Baar, A.W.; de Smit, J.; Uijttewaal, W.S.; Kleinhans, M.G. Sediment transport of fine sand to fine gravel on transverse bed slopes in rotating annular flume experiments. Water Resour. Res. 2018, 54, 19–45. [Google Scholar] [CrossRef]
- Fisher, J.S.; Sill, B.L.; Clark, D.F. Organic detritus particles: Initiation of motion criteria on sand and gravel beds. Water Resour. Res. 1983, 19, 1627–1631. [Google Scholar] [CrossRef]
- Tlili, L. The Effect of External Controls on Channel-Belt Stacking Patterns in the Fluvio-Deltaic Domain of a Landscape Flume. Master’s Thesis, Utrecht University, Utrecht, The Netherlands, 2013. [Google Scholar]
- Nepf, H. Vegetated flow dynamics. Ecogeomorphol. Tidal Marshes 2004, 59, 137–163. [Google Scholar]
- Nikora, V.; Larned, S.; Nikora, N.; Debnath, K.; Cooper, G.; Reid, M. Hydraulic resistance due to aquatic vegetation in small streams: Field study. J. Hydraul. Eng. 2008, 134, 1326–1332. [Google Scholar] [CrossRef]
- Kadlec, R.H. Overland flow in wetlands: Vegetation resistance. J. Hydraul. Eng. 1990, 116, 691–706. [Google Scholar] [CrossRef]
- Lee, J.K.; Roig, L.C.; Jenter, H.L.; Visser, H.M. Drag coefficients for modeling flow through emergent vegetation in the Florida Everglades. Ecol. Eng. 2004, 22, 237–248. [Google Scholar] [CrossRef]
- Nepf, H.M. Flow and transport in regions with aquatic vegetation. Annu. Rev. Fluid Mech. 2012, 44, 123–142. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.; Hiatt, M.; Passalacqua, P. Hydrological connectivity in vegetated river deltas: The importance of patchiness below a threshold. Geophys. Res. Lett. 2018, 45, 10–416. [Google Scholar] [CrossRef]
- Hejduk, L.; Banasik, K. Variations in Suspended Sediment Grain Sizes in Flood Events of a Small Lowland River; IAHS Publication: Wallingford, UK, 2010; Volume 337, pp. 189–196. [Google Scholar]
- Noe, G.B.; Harvey, J.W.; Schaffranek, R.W.; Larsen, L.G. Controls of suspended sediment concentration, nutrient content, and transport in a subtropical wetland. Wetlands 2010, 30, 39–54. [Google Scholar] [CrossRef]
- Aiona, A.R. Can a Constructed Stormwater Facility Remove Fine Particles from Urban Runoff? Master’s Thesis, University of California, Davis, CA, USA, 2013. [Google Scholar]
- McKenzie, E.R.; Wong, C.M.; Green, P.G.; Kayhanian, M.; Young, T.M. Size dependent elemental composition of road-associated particles. Sci. Total Environ. 2008, 398, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Comprehensive Manual for Velocimeters; Nortek: Rud, Norway, 2018.
- Goring, D.G.; Nikora, V.I. Despiking acoustic Doppler velocimeter data. J. Hydraul. Eng. 2002, 128, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Ku, H.H. Notes on the use of propagation of error formulas. J. Res. Natl. Bureau Stand. 1966, 70, 263–273. [Google Scholar] [CrossRef]
- Schnurr, P.J.; Espie, G.S.; Allen, D.G. The effect of light direction and suspended cell concentrations on algal biofilm growth rates. Appl. Microbiol. Biotechnol. 2014, 98, 8553–8562. [Google Scholar] [CrossRef]
- Trulear, M.G.; Characklis, W.G. Dynamics of biofilm processes. J. Water Pollut. Control Fed. 1982, 54, 1288–1301. [Google Scholar]
- Liu, D.; Lau, Y.; Chau, Y.; Pacepavicius, G. Simple technique for estimation of biofilm accumulation. Bull. Environ. Contam. Toxicol. 1994, 53, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Characklis, W.G.; Trulear, M.; Bryers, J.; Zelver, N. Dynamics of biofilm processes: Methods. Water Res. 1982, 16, 1207–1216. [Google Scholar] [CrossRef]








| Parameter | This Study | Fauria et al. (2015) | Purich (2006) | Natural |
|---|---|---|---|---|
| Flow velocity ( /) | 2.0–6.0 | 1.8–6.1 | 1.0–10.2 | 0–25 [54] |
| Flow depth ( ) | 40 | 14–17 | 12 | 0–50 [55] |
| Reynolds number 1 | 8400–25,200 | 2520–10,400 | 1400–12,800 | 70–65,000 [8,56] |
| Collector shape | Cylindrical | Bladed | Cylindrical | Varies |
| Collector diameter ( ) | 0.318 | 0.3 | 0.6 | 0.1–1.2 [57,58] |
| Collector Reynolds number | 66–200 | 54–183 | 70–640 | 5–1000 [55] |
| Collector density () | 285–1487 | 2724–7209 | 1013–4053 | 10–2700 [58] |
| Solid volume fraction (%) | 0.22–1.17 | 0.82–2.16 | 2.86–11.5 | 0.1–1 [57] |
| Particle type | Walnut shell | Road dust | Pliolite® | Sediment |
| Particle density ( /) | 1.53 | 2.27–2.61 2 | 1.03 | 1.43–2.39 [49] |
| Average particle diameter, ( ) | 25.2 | ∼10–15 3 | 212–250 | 45–100 [59,60] |
| Suspended concentration ( /) | 5–55 | <9–50 | ∼110 | 2–25 [60,61] |
| Particle-collector diameter ratio, R | 0.0079 | 0.0004–0.083 3 | 0.037 | <0.25 |
| Mid-Water-Column TKE (mm2/s2) | ||
|---|---|---|
| Control | 67 | 2.48 |
| 134 | 2.98 | |
| 200 | 3.37 | |
| 0.22% | 67 | 1.23 |
| 134 | 5.26 | |
| 200 | 12.2 | |
| 0.64% | 67 | 9.13 |
| 134 | 35.9 | |
| 200 | 59.6 | |
| 1.17% | 67 | 11.1 |
| 134 | 29.8 | |
| 200 | 54.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wingenroth, J.; Yee, C.; Nghiem, J.; Larsen, L. Effects of Stem Density and Reynolds Number on Fine Sediment Interception by Emergent Vegetation. Geosciences 2021, 11, 136. https://doi.org/10.3390/geosciences11030136
Wingenroth J, Yee C, Nghiem J, Larsen L. Effects of Stem Density and Reynolds Number on Fine Sediment Interception by Emergent Vegetation. Geosciences. 2021; 11(3):136. https://doi.org/10.3390/geosciences11030136
Chicago/Turabian StyleWingenroth, Jordan, Candace Yee, Justin Nghiem, and Laurel Larsen. 2021. "Effects of Stem Density and Reynolds Number on Fine Sediment Interception by Emergent Vegetation" Geosciences 11, no. 3: 136. https://doi.org/10.3390/geosciences11030136
APA StyleWingenroth, J., Yee, C., Nghiem, J., & Larsen, L. (2021). Effects of Stem Density and Reynolds Number on Fine Sediment Interception by Emergent Vegetation. Geosciences, 11(3), 136. https://doi.org/10.3390/geosciences11030136