
The Tetrapod Fossil Record from the Uppermost Maastrichtian of the Ibero-Armorican Island: An Integrative Review Based on the Outcrops of the Western Tremp Syncline (Aragón, Huesca Province, NE Spain)

 , , , , ,
, , , , ,
Abstract
:1. Introduction
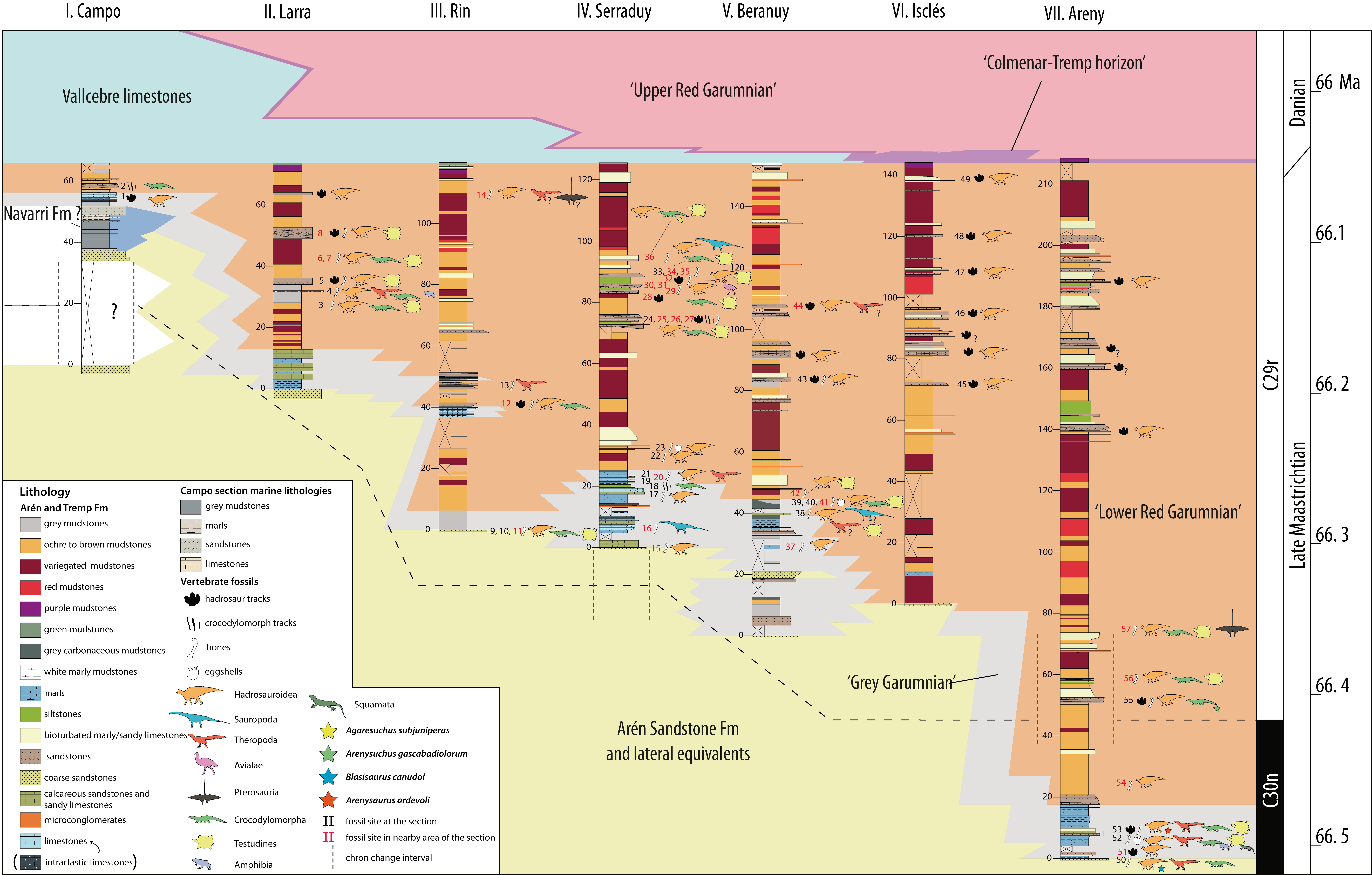
2. The Geological and Stratigraphic Framework of the Western Tremp Syncline (Aragonese Outcrops of the Tremp Fm)

3. The Upper Maastrichtian Tetrapod Fossil Record of the Western Tremp Syncline and Its Integration within the Ibero-Armorican Island Record
3.1. Dinosauria
3.1.1. Hadrosauroidea
3.1.2. Sauropoda
3.1.3. Theropoda
3.2. Pterosauria
3.3. Crocodylomorpha
3.4. Testudines
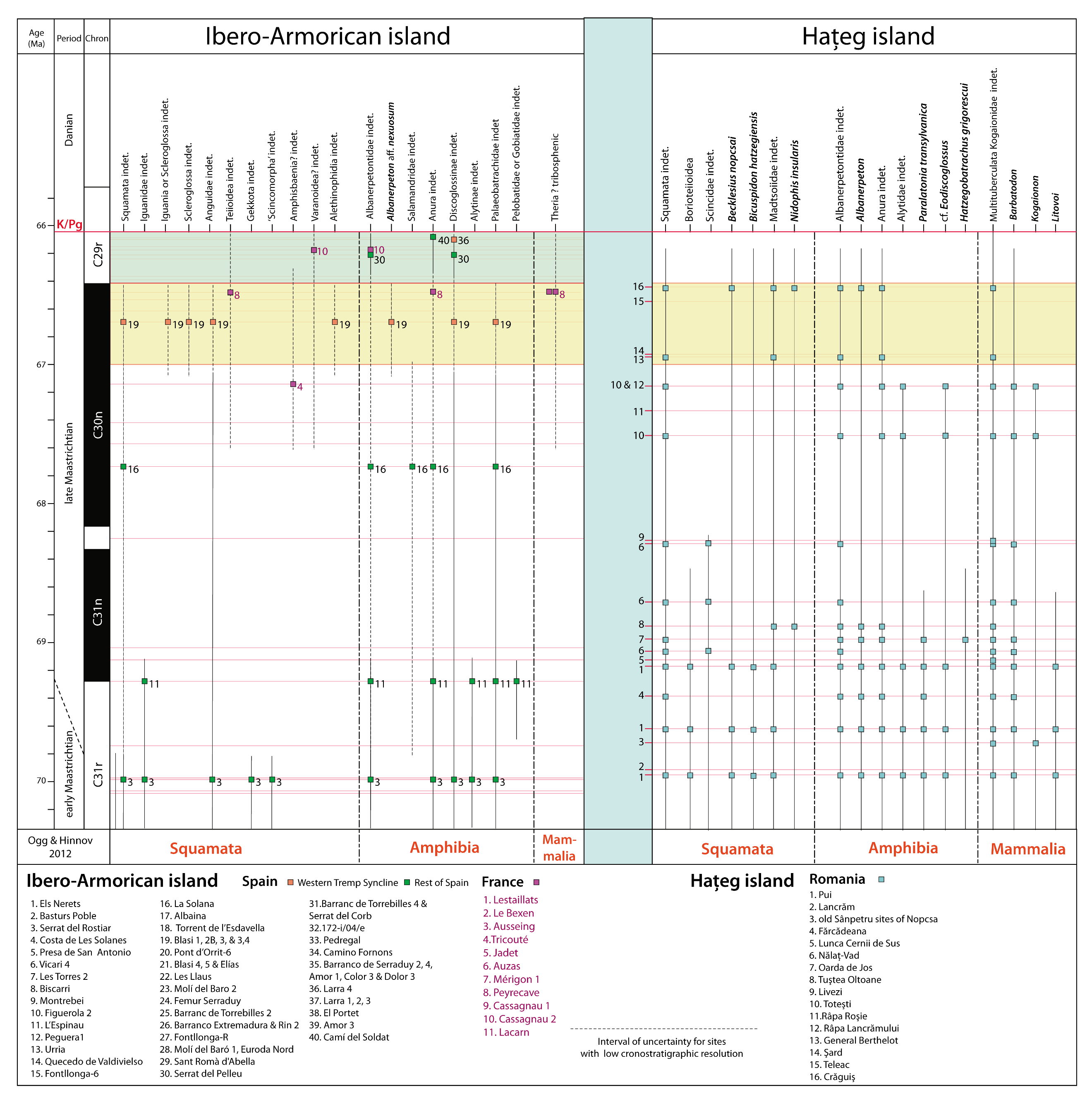
3.5. Amphibia and Squamata
3.6. The Tetrapod Fossil Record from the Upper Maastrichtian of the Ibero-Armorican Island
4. Discussion
4.1. Comparison with the Upper Maastrichtian Vertebrate Assemblage from the Hațeg Island
4.1.1. Dinosauria
4.1.2. Pterosauria
4.1.3. Crocodylomorpha
4.1.4. Testudines
4.1.5. Amphibia and Squamata
4.1.6. Mammalia
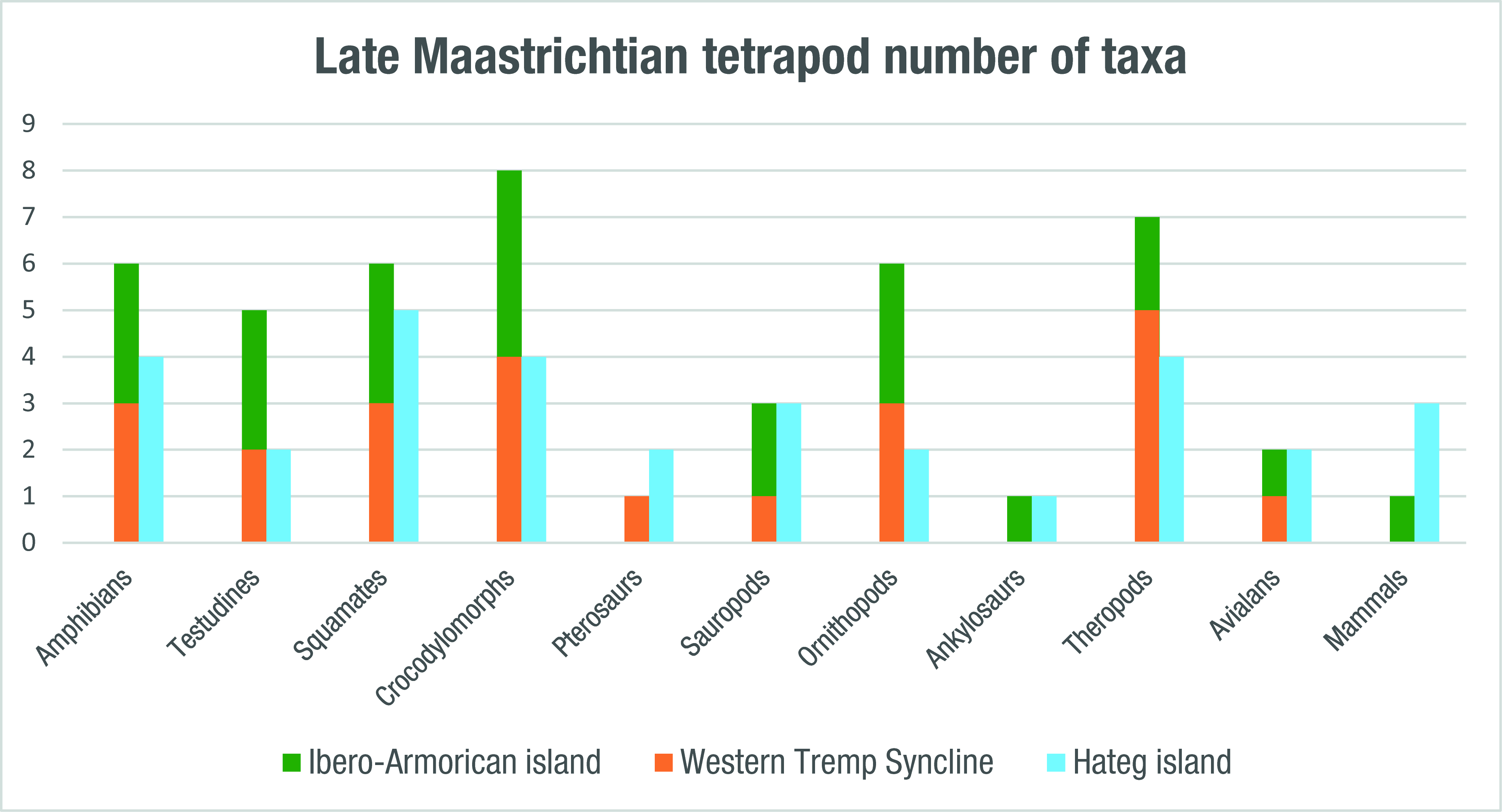
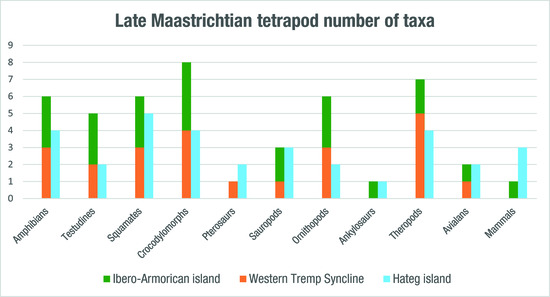
4.2. Evaluation of the Tetrapod Diversity of the Ibero-Armorican Island and Its Biases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alvarez, L.W.; Alvarez, W.; Asaro, F.; Michel, H.V. Extraterrestrial cause for the cretaceous-tertiary extinction. Science 1980, 208, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.G.; Kominz, M.A.; Browning, J.V.; Wright, J.D.; Mountain, G.S.; Katz, M.E.; Sugarman, P.J.; Cramer, B.S.; Christie-Blick, N.; Peka, S.F. The Phanerozoic Record of Global Sea-Level Change. Science 2005, 310, 1293–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Keller, G. Maastrichtian climate, productivity and faunal turnovers in planktic foraminifera in South Atlantic DSDP sites 525A and 21. Mar. Micropaleontol. 1998, 33, 55–86. [Google Scholar] [CrossRef]
- Barnet, J.S.K.; Littler, K.; Kroon, D.; Leng, M.J.; Westerhold, T.; Röhl, U.; Zachos, J.C. A new high-resolution chronology for the late Maastrichtian warming event: Establishing robust temporal links with the onset of Deccan volcanism. Geology 2018, 46, 147–150. [Google Scholar] [CrossRef] [Green Version]
- Courtillot, V.; Féraud, G.; Maluski, H.; Vandamme, D.; Moreau, M.G.; Besse, J. Deccan flood basalts and the Cretaceous/Tertiary boundary. Nature 1988, 333, 843–846. [Google Scholar] [CrossRef]
- Courtillot, V.E.; Renne, P.R. On the ages of flood basalt events. Comptes Rendus Geosci. 2003, 335, 113–140. [Google Scholar] [CrossRef] [Green Version]
- Tobin, T.S.; Ward, P.D.; Steig, E.J.; Olivero, E.B.; Hilburn, I.A.; Mitchell, R.N.; Diamond, M.R.; Raub, T.D.; Kirschvink, J.L. Extinction patterns, δ18 O trends, and magnetostratigraphy from a southern high-latitude Cretaceous–Paleogene section: Links with Deccan volcanism. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 180–188. [Google Scholar] [CrossRef]
- Schoene, B.; Eddy, M.P.; Samperton, K.M.; Keller, C.B.; Keller, G.; Adatte, T.; Khadri, S.F.R. U-Pb constraints on pulsed eruption of the Deccan Traps across the end-Cretaceous mass extinction. Science 2019, 363, 862–866. [Google Scholar] [CrossRef]
- Hildebrand, A.R.; Penfield, G.T.; Kring, D.A.; Pilkington, M.; Camargo, Z.A.; Jacobsen, S.B.; Boynton, W.V. Chicxulub Crater: A possible Cretaceous/Tertiary boundary impact crater on the Yucatán Peninsula, Mexico. Geology 1991, 19, 867. [Google Scholar] [CrossRef]
- Renne, P.R.; Arenillas, I.; Arz, J.A.; Vajda, V.; Gilabert, V.; Bermúdez, H.D. Multi-proxy record of the Chicxulub impact at the Cretaceous-Paleogene boundary from Gorgonilla Island, Colombia. Geology 2018, 46, 547–550. [Google Scholar] [CrossRef]
- Maruoka, T. Mass Extinction at the cretaceous–paleogene (K–Pg) boundary. In Astrobiology; Springer: Singapore, 2019; pp. 303–320. [Google Scholar]
- Lyons, S.L.; Karp, A.T.; Bralower, T.J.; Grice, K.; Schaefer, B.; Gulick, S.P.S.; Morgan, J.V.; Freeman, K.H. Organic matter from the Chicxulub crater exacerbated the K–Pg impact winter. Proc. Natl. Acad. Sci. USA 2020, 117, 25327–25334. [Google Scholar] [CrossRef]
- Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; et al. The chicxulub asteroid impact and mass extinction at the cretaceous-paleogene boundary. Science 2010, 327, 1214–1218. [Google Scholar] [CrossRef] [Green Version]
- Witts, J.D.; Whittle, R.J.; Wignall, P.B.; Crame, J.A.; Francis, J.E.; Newton, R.J.; Bowman, V.C. Macrofossil evidence for a rapid and severe Cretaceous–Paleogene mass extinction in Antarctica. Nat. Commun. 2016, 7, 11738. [Google Scholar] [CrossRef]
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17084–17093. [Google Scholar] [CrossRef]
- Dzombak, R.M.; Sheldon, N.D.; Mohabey, D.M.; Samant, B. Stable climate in India during Deccan volcanism suggests limited influence on K–Pg extinction. Gondwana Res. 2020, 85, 19–31. [Google Scholar] [CrossRef]
- Raup, D.M.; Sepkoski, J.J. Mass extinctions in the marine fossil record. Science 1982, 215, 1501–1503. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, D. Extinctions in the fossil record. Philos. Trans. R. Soc. London Ser. B Biol. Sci. 1994, 344, 11–17. [Google Scholar] [CrossRef]
- Bardet, N. Extinction events among Mesozoic marine reptiles. Hist. Biol. 1994, 7, 313–324. [Google Scholar] [CrossRef]
- Longrich, N.R.; Tokaryk, T.; Field, D.J. Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. Proc. Natl. Acad. Sci. USA 2011, 108, 15253–15257. [Google Scholar] [CrossRef] [Green Version]
- Longrich, N.R.; Martill, D.M.; Andres, B. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLOS Biol. 2018, 16, e2001663. [Google Scholar] [CrossRef] [Green Version]
- Brusatte, S.L.; Butler, R.J.; Barrett, P.M.; Carrano, M.T.; Evans, D.C.; Lloyd, G.T.; Mannion, P.D.; Norell, M.A.; Peppe, D.J.; Upchurch, P.; et al. The extinction of the dinosaurs. Biol. Rev. 2015, 90, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Puértolas-Pascual, E.; Blanco, A.; Brochu, C.A.; Canudo, J.I. Review of the late cretaceous-early paleogene crocodylomorphs of Europe: Extinction patterns across the K-PG boundary. Cretac. Res. 2016, 57, 565–590. [Google Scholar] [CrossRef]
- Pearson, D.A.; Schaefer, T.; Johnson, K.R.; Nichols, D.J.; Hunter, J.P. Vertebrate biostratigraphy of the Hell Creek Formation in southwestern North Dakota and northwestern South Dakota. Spec. Pap. Geol. Soc. Am. 2002, 361, 145–167. [Google Scholar] [CrossRef]
- Lyson, T.R.; Longrich, N.R. Spatial niche partitioning in dinosaurs from the latest Cretaceous (Maastrichtian) of North America. Proc. R. Soc. B Biol. Sci. 2011, 278, 1158–1164. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.R.; Nichols, D.J.; Hartman, J.H. Hell Creek Formation: A 2001 synthesis. Geol. Soc. Am. Spec. Pap. 2002, 361, 503–510. [Google Scholar]
- LeCain, R.; Clyde, W.C.; Wilson, G.P.; Riedel, J. Magnetostratigraphy of the Hell Creek and lower Fort Union Formations in northeastern Montana. In Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas; Geological Society of America: Boulder, CO, USA, 2014. [Google Scholar]
- Sprain, C.J.; Renne, P.R.; Clemens, W.A.; Wilson, G.P. Calibration of chron C29r: New high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana. GSA Bull. 2018, 130, 1615–1644. [Google Scholar] [CrossRef]
- Fowler, D. The Hell Creek Formation, Montana: A stratigraphic review and revision based on a sequence stratigraphic approach. Geosciences 2020, 10, 435. [Google Scholar] [CrossRef]
- Csiki-Sava, Z.; Buffetaut, E.; Ősi, A.; Pereda-Suberbiola, X.; Brusatte, S.L. Island life in the Cretaceous—Faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. Zookeys 2015, 469, 1–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canudo, J.I.; Oms, O.; Vila, B.; Galobart, À.; Fondevilla, V.; Puértolas-Pascual, E.; Sellés, A.G.; Cruzado-Caballero, P.; Dinarès-Turell, J.; Vicens, E.; et al. The upper Maastrichtian dinosaur fossil record from the southern Pyrenees and its contribution to the topic of the Cretaceous-Palaeogene mass extinction event. Cretac. Res. 2016, 57, 540–551. [Google Scholar] [CrossRef]
- Csiki-Sava, Z.; Vremir, M.; Vasile, Ş.; Brusatte, S.L.; Dyke, G.; Naish, D.; Norell, M.A.; Totoianu, R. The east side story—The Transylvanian latest Cretaceous continental vertebrate record and its implications for understanding Cretaceous–Paleogene boundary events. Cretac. Res. 2016, 57, 662–698. [Google Scholar] [CrossRef]
- Fondevilla, V.; Riera, V.; Vila, B.; Sellés, A.G.; Dinarès-Turell, J.; Vicens, E.; Gaete, R.; Oms, O. Galobart Chronostratigraphic synthesis of the latest Cretaceous dinosaur turnover in south-western Europe. Earth Sci. Rev. 2019, 191, 168–189. [Google Scholar] [CrossRef]
- Vila, B.; Sellés, A.G.; Brusatte, S.L. Diversity and faunal changes in the latest Cretaceous dinosaur communities of southwestern Europe. Cretac. Res. 2016, 57, 552–564. [Google Scholar] [CrossRef]
- Gómez-Gras, D.; Roigé, M.; Fondevilla, V.; Oms, O.; Boya, S.; Remacha, E. Provenance constraints on the Tremp Formation paleogeography (southern Pyrenees): Ebro Massif VS Pyrenees sources. Cretac. Res. 2016, 57, 414–427. [Google Scholar] [CrossRef]
- Puigdefàbregas, C.; Muñoz, J.A.; Marzo, M. Thrust Belt Development in the Eastern Pyrenees and Related Depositional Sequences in the Southern Foreland Basin; Wiley: Hoboken, NJ, USA, 1986. [Google Scholar]
- Muñoz, J.A. Evolution of a continental collision belt: ECORS-Pyrenees crustal balanced cross-section. In Thrust Tectonics; Springer: Dordrecht, The Netherlands, 1992; pp. 235–246. [Google Scholar]
- Teixell, A. Crustal structure and orogenic material budget in the west central Pyrenees. Tectonics 1998, 17, 395–406. [Google Scholar] [CrossRef]
- Teixell, A. Estructura de los Pirineos: Generalidades. In Geología de España; SGE-IGME: Madrid, Spain, 2004; pp. 321–323. [Google Scholar]
- Costa, E.; Garcés, M.; López-Blanco, M.; Beamud, E.; Gómez-Paccard, M.; Larrasoaña, J.C. Closing and continentalization of the South Pyrenean foreland basin (NE Spain): Magnetochronological constraints. Basin Res. 2009, 36, 349–364. [Google Scholar] [CrossRef]
- Ardévol, L.; Klimowitz, J.; Malagón, J.; Nagtegaal, P.J.C. Depositional sequence response to foreland deformation in the upper Cretaceous of the Southern Pyrenees, Spain. Am. Assoc. Pet. Geol. Bull. 2000, 84, 566–588. [Google Scholar] [CrossRef]
- Oms, O.; Fondevilla, V.; Riera, V.; Marmi, J.; Vicens, E.; Estrada, R.; Vila, B. Transitional environments of the lower Maastrichtian South-Pyrenean Basin (Catalonia, Spain): The Fumanya Member tidal flat. Cretac. Res. 2016, 57, 428–442. [Google Scholar] [CrossRef]
- Rosell, J.; Linares, R.; Llompart, C. El “garumniense” prepirenaico. Rev. Soc. Geol. España 2001, 14, 47–56. [Google Scholar]
- Mey, P.H.W.; Nagtegaal, P.J.C.; Roberti, K.J.; Hartevelt, J.J.A. Lithostratigraphic subdivision of Post-Hercynian deposits in the South-Central Pyrenees, Spain. Leidse Geol. Meded. 1968, 41, 221–228. [Google Scholar]
- Nagtegaal, P.J.C.; Van Vliet, A.; Brouwer, J. Syntectonic coastal offlap and concurrent turbidite deposition: The Upper Cretaceous Aren sandstone in the South-Central Pyrenees, Spain. Sediment. Geol. 1983, 34, 185–218. [Google Scholar] [CrossRef]
- Mutti, E.; Sgavetti, M. Sequence stratigraphy of the Upper Cretaceous Aren strata in the Aren–Orcau region, south-central Pyrenees, Spain: Distinction between eustatically and tectonically controlled depositional sequences. Ann. Univ. Ferrara 1987, 1, 1–22. [Google Scholar]
- Nagtegaal, P.J.C. Depositional history and clay minerals of the Upper Cretaceous basin in the south-central Pyrenees, Spain. Leidse Geol. Meded. 1972, 47, 251–275. [Google Scholar]
- Mutti, E.; Rosell, J.; Ghibaudo, G.; Obrador, A. The Upper Cretaceous Aren Sandstone in its type-area. In Proceedings of the 9th International Congress International Association of Sedimentologists, Nice, France, 1 January–30 November 1975; pp. 7–15. [Google Scholar]
- Díaz-Molina, M.; Kälin, O.; Benito, M.I.; Lopez-Martinez, N.; Vicens, E. Depositional setting and early diagenesis of the dinosaur eggshell-bearing Aren Fm at Bastus, Late Campanian, south-central Pyrenees. Sediment. Geol. 2007, 199, 205–221. [Google Scholar] [CrossRef] [Green Version]
- Leymerie, A. Présence de garumnien en Espagne. Bulletin de la Sociéte Géologique de France. Bull. Soc. Géol. Fr. 1868, 25, 906–911. [Google Scholar]
- Cuevas, J.L. Estratigrafía del «Garumniense» de la Conca de Tremp. Prepirineo de Lérida. Acta Geológica Hispánica 1992, 27, 95–108. [Google Scholar]
- Pujalte, V.; Schmitz, B. The stratigraphy of the Tremp Group revisited (Garumnian, Tremp-Graus basin, South Pyrenees). Geogaceta 2005, 38, 79–82. [Google Scholar]
- Díaz-Molina, M. Sedimentación sintectónica asociada a una subida relativa del nivel del mar durante el Cretácico Superior (Fm. Tremp, provincia de Lérida). Estud. Geol. Núm. Extraordin. Galve Tremp. 1987, 43, 69–93. [Google Scholar]
- Eichenseer, H. Facies Geology of Late Maastrichtian to Early Eocene Coastal and Shallow Marine Sediments (Tremp-Graus Basin, Northeastern Spain); Universität Tübingen: Tübingen, Germany, 1988. [Google Scholar]
- Riera, V.; Oms, O.; Gaete, R.; Galobart, À. The end-Cretaceous dinosaur succession in Europe: The Tremp Basin record (Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 283, 160–171. [Google Scholar] [CrossRef]
- Díez-Canseco, D.; Arz, J.A.; Benito, M.I.; Díaz-Molina, M.; Arenillas, I. Tidal influence in redbeds: A palaeoenvironmental and biochronostratigraphic reconstruction of the Lower Tremp Formation (South-Central Pyrenees, Spain) around the Cretaceous/Paleogene boundary. Sediment. Geol. 2014, 312, 31–49. [Google Scholar] [CrossRef]
- Blanco, A.; Szabó, M.; Blanco-Lapaz, À.; Marmi, J. Late Cretaceous (Maastrichtian) Chondrichthyes and Osteichthyes from northeastern Iberia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 465, 278–294. [Google Scholar] [CrossRef]
- Ghinassi, M.; Oms, O.; Cosma, M.; Finotello, A.; Munari, G. Reading tidal processes where their signature is cryptic: The Maastrichtian meandering channel deposits of the Tremp Formation (Southern Pyrenees, Spain). Sedimentology 2020. [Google Scholar] [CrossRef]
- Villalba-Breva, S.; Martín-Closas, C. Upper Cretaceous paleogeography of the Central Southern Pyrenean Basins (Catalonia, Spain) from microfacies analysis and charophyte biostratigraphy. Facies 2013, 59, 319–345. [Google Scholar] [CrossRef]
- Vicente, A.; Martín-Closas, C.; Arz, J.A.; Oms, O. Maastrichtian-basal Paleocene charophyte biozonation and its calibration to the Global Polarity Time Scale in the southern Pyrenees (Catalonia, Spain). Cretac. Res. 2015, 52, 268–285. [Google Scholar] [CrossRef]
- Vicente, A.; Villalba-Breva, S.; Ferràndez-Cañadell, C.; Martín-Closas, C. Revision of the Maastrichtian-Palaeocene charophyte biostratigraphy of the Fontllonga reference section (Southern Pyrenees, Catalonia, Spain). Geol. Acta 2016, 14, 349–362. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Canudo, J.I.; Cruzado-Caballero, P.; Barco, J.L.; López-Martínez, N.; Oms, O.; Ruiz-Omeñaca, J.I. The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain). Comptes Rendus Palevol 2009, 8, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Fondevilla, V.; Dinarès-Turell, J.; Oms, O. The chronostratigraphic framework of the South-Pyrenean Maastrichtian succession reappraised: Implications for basin development and end-Cretaceous dinosaur faunal turnover. Sediment. Geol. 2016, 337, 55–68. [Google Scholar] [CrossRef]
- Puértolas-Pascual, E.; Arenillas, I.; Arz, J.A.; Calvín, P.; Ezquerro, L.; García-Vicente, C.; Pérez-Pueyo, M.; Sánchez-Moreno, E.M.; Villalaín, J.J.; Canudo, J.I. Chronostratigraphy and new vertebrate sites from the upper Maastrichtian of Huesca (Spain), and their relation with the K/Pg boundary. Cretac. Res. 2018, 89, 36–59. [Google Scholar] [CrossRef] [Green Version]
- López-Martínez, N.; Vicens, E. A new peculiar dinosaur egg, Sankofa pyrenaica oogen. nov. oosp. nov. from the Upper Cretaceous coastal deposits of the Aren Formation, south-central Pyrenees, Lleida, Catalonia, Spain. Palaeontology 2012, 55, 325–339. [Google Scholar] [CrossRef]
- Riera, V. Estudio Integrado (Geología y Paleontología) de la Sucesión de Dinosaurios (Maastrichtiense) de la Vertiente Surpirenaica; Universitat Autónoma de Barcelona: Barcelona, Spain, 2010. [Google Scholar]
- López-Martínez, N.; Arribas, M.E.; Robador, A.; Vicens, E.; Ardévol, L. Los carbonatos danienses (Unidad 3) de la Fm Temp (Pirineos sur-centrales): Paleogeografía y relación con el límite Cretácico-Terciario. Rev. Soc. Geol. Esp. 2006, 19, 233–255. [Google Scholar]
- Robador, A.; Samsó, J.M.; Serra-Kiel, J.; Tosquella, J. Field guide. In Introduction to the early Paleogene of the south Pyrenean basin. Field trip Guidebook; Barnolas, A., Robador, A., Serra-Kiel, J., Caus, E., Eds.; IGME: Barcelona, Spain, 1990; pp. 131–159. [Google Scholar]
- Serra-Kiel, P.; Canudo, J.I.; Dinares, J.; Molina, E.; Ortiz, N.; Pascual, J.O.; Samso, J.M.; Tosquella, J. Cronoestratigrafía de los sedimentos marinos del Terciario inferior de la Cuenca de Tremp-Graus (Zona Central Surpirenaica). Rev. Soc. Esp. Geol. 1994, 7, 273–299. [Google Scholar]
- Pujalte, V.; Schmitz, B.; Baceta, J.I. Sea-level changes across the Paleocene–Eocene interval in the Spanish Pyrenees, and their possible relationship with North Atlantic magmatism. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 393, 45–60. [Google Scholar] [CrossRef]
- Vila, B.; Galobart, À.; Canudo, J.I.; Le Loeuff, J.; Dinarès-Turell, J.; Riera, V.; Oms, O.; Tortosa, T.; Gaete, R. The diversity of sauropod dinosaurs and their first taxonomic succession from the latest Cretaceous of southwestern Europe: Clues to demise and extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 19–38. [Google Scholar] [CrossRef]
- López-Martínez, N.; Canudo, J.I.; Ardevol, L.; Pereda-Suberbiola, X.; Orue-Etxebarria, X.; Cuenca-Bescós, G.; Ruiz-Omeñaca, J.I.; Murelaga, X.; Feist, M. New dinosaur sites correlated with upper Maastrichtian pelagic deposits in the Spanish Pyrenees: Implications for the dinosaur extinctio. Cretac. Res. 2001, 22, 41–61. [Google Scholar] [CrossRef] [Green Version]
- Garrido Mejías, A.; Ríos Aragües, L.M. Síntesis geológica del Secundario y Terciario entre los ríos Cinca y Segre. Bol. Inst. Geol. Min. Esp. 1972, 83, 1–47. [Google Scholar]
- Canudo, J.I. The Collection of Type Fossils of the Natural Science Museum of the University of Zaragoza (Spain). Geoheritage 2018, 10, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Pereda-Suberbiola, X.; Canudo, J.I.; Company, J.; Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I. Hadrosauroid dinosaurs from the latest Cretaceous of the Iberian Peninsula. J. Vertebr. Paleontol. 2009, 29, 946–951. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Review of the fossill record of spanish hadrosaur remains. In Proceedings of the VIII Encuentro de Jóvenes Investigadores en Paleontología, Enciso, Spain, 21–22 April 2010; Volume 30, pp. 99–105. [Google Scholar]
- Cruzado-Caballero, P.; Puértolas-Pascual, E.; Canudo, J.I.; Castanera, D.; Gasca, J.M.; Moreno-Azanza, M. New hadrosaur remains from the Late Maastrichtian of Huesca (NE Spain). In Proceedings of the 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; pp. 45–48. [Google Scholar]
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Gaete, R.; Riera, V.; Oms, O.; Canudo, J.I. A new hadrosaurid dentary from the latest Maastrichtian of the Pyrenees (north Spain) and the high diversity of the duck-billed dinosaurs of the Ibero-Armorican Realm at the very end of the Cretaceous. Hist. Biol. 2014, 26, 619–630. [Google Scholar] [CrossRef]
- Company, J.; Cruzado-Caballero, P.; Canudo, J.I. Presence of diminutive hadrosaurids (Dinosauria: Ornithopoda) in the Maastrichtian of the south-central Pyrenees (Spain). J. Iber. Geol. 2015, 41, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Márquez, A.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À. Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin Canardia garonnensis. PLoS ONE 2013, 8, e69835. [Google Scholar] [CrossRef] [Green Version]
- Conti, S.; Vila, B.; Sellés, A.G.; Galobart, À.; Benton, M.J.; Prieto-Márquez, A. The oldest lambeosaurine dinosaur from Europe: Insights into the arrival of Tsintaosaurini. Cretac. Res. 2020, 107, 104286. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (hadrosauridae) from the latest cretaceous of Arén (Huesca, Spain). Can. J. Earth Sci. 2010, 47, 1507–1517. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Canudo, J.I.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I. New material and phylogenetic position of Arenysaurus ardevoli, a lambeosaurine dinosaur from the late Maastrichtian of Arén (Northern Spain). J. Vertebr. Paleontol. 2013, 33, 1367–1384. [Google Scholar] [CrossRef]
- Longrich, N.R.; Suberbiola, X.P.; Pyron, R.A.; Jalil, N.-E. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretac. Res. 2020, 104678. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Canudo Sanagustín, J.; Ruiz-Omeñaca, J.I. Nuevas evidencias de la presencia de hadrosaurios lambeosaurios (Dinosauria) en el Maastrichtiense superior de la Península Ibérica (Arén, Huesca). Geogaceta 2005, 3, 47–50. [Google Scholar]
- Cruzado-Caballero, P.; Canudo, J.I.; Ruiz-Omeñaca, J.I. Los fémures de Blasi (Arén, Huesca, Spain): Una contribución a los hadrosauroideos Europeos del Maastrichtiense superior. In Proceedings of the Actas de las IV Jornadas Internacionales sobre Paleontologıa de Dinosaurios y su Entorno, Burgos, Spain, 5–9 September 2009; pp. 197–205. [Google Scholar]
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Evidencias de la coexistencia de dinosaurios hadrosaurinos y lambeosaurinos en el Maastrichtiano superior de la Península Ibérica (Arén, Huesca, España). Ameghiniana 2010, 47, 153–164. [Google Scholar] [CrossRef]
- Canudo, I.; Cruzado-Caballero, P.; Moreno-Azanza, M. Possible theropod predation evidence in hadrosaurid dinosaurs from the Upper Maastrichtian (Upper Cretaceous) of Arén (Huesca, Spain). Kaupia Darmstädter Beiträge zur Naturgeschichte 2005, 14, 9–13. [Google Scholar]
- Cruzado-Caballero, P.; Fortuny, J.; Llacer, S.; Canudo, J.I. Paleoneuroanatomy of the European lambeosaurine dinosaur Arenysaurus ardevoli. Peer J. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mayayo-Lainez, A. Paleohistología y Mineralogía de los Dinosaurios Hadrosaurios del Maastrichtiense de Arén (Huesca). Master’s Thesis, University of Zaragoza, Zaragoza, Spain, 2020. [Google Scholar]
- Mayayo-Lainez, A.; Alegre-Esteve, M.; Bauluz, B.; Canudo, J. First approach to the paleohistology of the hadrosaur dinosaurs from Blasi 2A (Tremp Formation, Maastrichtian, Huesca). Ciências Terra-Procedia 2021, 1, 38–41. [Google Scholar]
- Puértolas-Pascual, E.; Cruzado-Caballero, P.; Canudo, J.I.; Gasca, J.M.; Moreno-Azanza, M.; Castanera, D.; Parrillas, J.; Ezquerro, L. Nuevos yacimientos de vertebrados del Maastrichtiense superior (Cretácico Superior) de Huesca (España). In Proceedings of the VIII Geological Congress of Spain, Oviedo, Spain, 17–19 July 2012; pp. 269–272. [Google Scholar]
- Pérez Pueyo, M.; Puértolas-Pascual, E.; Bádenas, B. Larra 4: Desenterrando a los últimos vertebrados del maastrichtiense terminal del pirineo aragonés. Zubía 2019, 31, 159–163. [Google Scholar]
- Pérez-Pueyo, M.; Gilabert, V.; Moreno-Azanza, M.; Puertolas-Pascual, E.; Bádenas, B.; Canudo, J.I. Late Maastrichtian fossil assemblage of Veracruz 1 site (Beranuy, NE Spain): Wildfires and bones in a transitional environment. In Proceedings of the VIII Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Burgos, Spain, 5–9 September 2019; pp. 111–113. [Google Scholar]
- Barco, J.L.; Ardevol, L.; Canudo, J.I. Descripción de los primeros rastros asignados a Hadrosauridae (Ornithopoda, Dinosauria) del Maastrichtiense de la Península Ibérica (Areny, Huesca). Geogaceta 2001, 30, 235–238. [Google Scholar]
- Vila, B.; Oms, O.; Fondevilla, V.; Gaete, R.; Galobart, À.; Riera, V.; Canudo, J.I. The Latest Succession of Dinosaur Tracksites in Europe: Hadrosaur Ichnology, Track Production and Palaeoenvironments. PLoS ONE 2013, 8, e72579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellés, A.G.; Via, B.; Galobart, À. Spheroolithus europaeus, oosp. nov. (late Maastrichtian, Catalonia), the youngest oological record of hadrosauroids in Eurasia. J. Vertebr. Paleontol. 2014, 34, 725–729. [Google Scholar] [CrossRef]
- Pérez-Pueyo, M.; Moreno-Azanza, M.; Núñez-Lahuerta, C.; Puértolas-Pascual, E.; Bádenas, B.; Canudo, J.I. Eggshell association of the Late Maastrichtian (Late Cretaceous) at Blasi 2B fossil site: A scrambled of vertebrate diversity. Ciências da Terra-Procedia 2021, 1, 58–61. [Google Scholar]
- Vila, B.; Sellés, A.G. Re-evaluation of the age of some dinosaur localities from the southern Pyrenees by means of megaloolithid oospecies. J. Iber. Geol. 2015, 41. [Google Scholar] [CrossRef] [Green Version]
- Canudo, J.I. Descripcion de un fragmento proximal de fémur de Titanosauridae (Dinosauria, Sauropoda) del Maastrichtiense superior de Serraduy (Huesca). In Proceedings of the XVII Jornadas de la Sociedad Española de Paleontología, Albarracín, Spain, 18–20 October 2001; pp. 255–262. [Google Scholar]
- Sellés, A.G.; Marmi, J.; Llácer, S.; Blanco, A. The youngest sauropod evidence in Europe. Hist. Biol. 2016, 28, 930–940. [Google Scholar] [CrossRef]
- Fondevilla, V.; Vila, B.; Oms, O.; Galobarp, À. Skin impressions of the last European dinosaurs. Geol. Mag. 2017, 154, 393–398. [Google Scholar] [CrossRef]
- Díez Díaz, V.; Pereda Suberbiola, X.; Sanz, J.L. The axial skeleton of the titanosaur Lirainosaurus astibiae (Dinosauria: Sauropoda) from the latest Cretaceous of Spain. Cretac. Res. 2013, 43, 145–160. [Google Scholar] [CrossRef]
- Salgado, L.; Coria, R.A.; Calvo, J.O. Evolution of titanosaurid sauropods: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 1997, 34, 3–32. [Google Scholar]
- Upchurch, P.; Barrett, P.M.; Dodson, P. 13. Sauropoda. In The Dinosauria, 2nd ed.; University of California Press: Berkeley, CA, USA, 2019; pp. 259–322. [Google Scholar]
- Calvo, J.O.; Bonaparte, J.F. Andesaurus delgadoi gen. et. sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquen, Argentina. Ameghiniana 1991, 28, 303–310. [Google Scholar]
- Mannion, P.D.; Calvo, J.O. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: Implications for titanosaur systematics. Zool. J. Linn. Soc. 2011, 155–181. [Google Scholar] [CrossRef]
- Torices, A.; Currie, P.J.; Canudo, J.I.; Pereda-Suberbiola, X. Theropod dinosaurs from the upper cretaceous of the south pyrenees basin of Spain. Acta Palaeontol. Pol. 2015, 60, 611–626. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, A.; Ortega, F.; Bolet, A.; Escaso, F.; Houssaye, A.; Martínez-Salanova, J.; de Miguel Chaves, C.; Mocho, P.; Narváez, I.; Segura, M.; et al. A review of the upper Campanian vertebrate site of Armuña (Segovia Province, Spain). Cretac. Res. 2016, 57, 591–623. [Google Scholar] [CrossRef]
- Pérez-Pueyo, M.; Puértolas-Pascual, E.; Moreno-Azanza, M.; Cruzado-Caballero, P.; Gasca, J.M.; Núñez-Lahuerta, C.; Canudo, J.I. First record of a giant bird (Ornithuromorpha) from the uppermost Maastrichtian of the Southern Pyrenees, NE Spain. J. Vertebr. Paleontol. 2021, 41, e1900210. [Google Scholar]
- López-Martínez, N.; Canudo, J.I.; Cuenca-Bescós, G. Latest cretaceous eggshells from Arén (Southern Pyrenees, Spain). In Proceedings of the First International Symposium on Dinosaur Eggs and Babies, Isona, Spain, 23–26 September 1999; pp. 35–36. [Google Scholar]
- Núñez-Lahuerta, C.; Moreno-Azanza, M.; Pérez-Pueyo, M. First approach for a taphonomic key for fossil eggs and eggshells accumulations using optic microscopy: The case of Blasi-2B (Upper Cretaceous, Spain). Ciências Terra Procedia 2021, 1, 42–45. [Google Scholar]
- Choi, S.; Moreno-Azanza, M.; Csiki-Sava, Z.; Prondvai, E.; Lee, Y.N. Comparative crystallography suggests maniraptoran theropod affinities for latest Cretaceous European ‘geckoid’ eggshell. Pap. Palaeontol. 2020, 6, 265–292. [Google Scholar] [CrossRef]
- Dalla Vecchia, F.M.; Riera, F.M.; Oms, J.O.; Dinarès-Turell, J.; Gaete, R.; Galobart, A. The last pterosaurs: First record from the uppermost Maastrichtian of the Tremp Syncline (Northern Spain). Acta Geol. Sin. 2013, 87, 1198–1227. [Google Scholar] [CrossRef]
- Puértolas, E.; Canudo, J.I.; Cruzado-Caballero, P. A new crocodylian from the late Maastrichtian of Spain: Implications for the initial radiation of crocodyloids. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Blanco, A.; Puértolas-Pascual, E.; Marmi, J.; Vila, B.; Sellés, A.G. Allodaposuchus palustris sp. nov. from the Upper Cretaceous of Fumanya (South-Eastern Pyrenees, Iberian Peninsula): Systematics, Palaeoecology and Palaeobiogeography of the Enigmatic Allodaposuchian Crocodylians. PLoS ONE 2014, 9, e115837. [Google Scholar] [CrossRef]
- Blanco, A.; Fortuny, J.; Vicente, A.; Luján, À.H.; García-Marçà, J.A.; Sellés, A.G. A new species of Allodaposuchus (Eusuchia, Crocodylia) from the Maastrichtian (Late Cretaceous) of Spain: Phylogenetic and paleobiological implications. Peer J. 2015, 3, e1171. [Google Scholar] [CrossRef] [Green Version]
- Narváez, I.; Brochu, C.A.; Escaso, F.; Pérez-García, A.; Ortega, F. New Crocodyliforms from Southwestern Europe and Definition of a Diverse Clade of European Late Cretaceous Basal Eusuchians. PLoS ONE 2015, 10, e0140679. [Google Scholar] [CrossRef]
- Narváez, I.; Brochu, C.A.; Escaso, F.; Pérez-García, A.; Ortega, F. New Spanish Late Cretaceous eusuchian reveals the synchronic and sympatric presence of two allodaposuchids. Cretac. Res. 2016, 65, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Mateus, O.; Puértolas-Pascual, E.; Callapez, P.M. A new eusuchian crocodylomorph from the Cenomanian (Late Cretaceous) of Portugal reveals novel implications on the origin of Crocodylia. Zool. J. Linn. Soc. 2019, 186, 501–528. [Google Scholar] [CrossRef] [Green Version]
- Puértolas-Pascual, E.; Canudo, J.I.; Moreno-Azanza, M. The eusuchian crocodylomorph Allodaposuchus subjuniperus sp. nov., a new species from the latest Cretaceous (upper Maastrichtian) of Spain. Hist. Biol. 2014, 26, 91–109. [Google Scholar] [CrossRef]
- Blanco, A.; Puértolas-Pascual, E.; Marmi, J.; Moncunill-Solé, B.; Llácer, S.; Rössner, G.E. Late Cretaceous (Maastrichtian) crocodyliforms from north-eastern Iberia: A first attempt to explain the crocodyliform diversity based on tooth qualitative traits. Zool. J. Linn. Soc. 2020, 189, 584–617. [Google Scholar] [CrossRef]
- Moreno-Azanza, M.; Bauluz, B.; Canudo, J.I.; Puértolas-Pascual, E.; Sellés, A.G. A re-evaluation of aff. Megaloolithidae eggshell fragments from the uppermost Cretaceous of the Pyrenees and implications for crocodylomorph eggshell structure. Hist. Biol. 2014, 26, 195–205. [Google Scholar] [CrossRef]
- Hirsch, K.F. Fossil crocodilian eggs from the eocene of Colorado. J. Paleontol. 1985, 3, 531–542. [Google Scholar]
- Vila, B.; Castanera, D.; Marmi, J.; Canudo, J.I.; Galobart, À. Crocodile swim tracks from the latest Cretaceous of Europe. Lethaia 2015, 48, 256–266. [Google Scholar] [CrossRef]
- Pérez-Pueyo, M.; Castanera, D.; Bádenas, B.; Canudo, J.I. New evidences of Crocodylomorpha swim tracks in the Maastrichtian of Beranuy (Huesca, Spain). In Proceedings of the XVI Encuentro de Jóvenes Investigadores en Paleontología, Zarautz, Spain, 11–14 April 2018; pp. 137–140. [Google Scholar]
- De Lapparent de Broin, F.; Murelaga, X. Turtles from Upper Cretaceous of Lano (Iberian Peninsula). Comptes Rendus l’Académie Sci. 1996, 323, 729–735. [Google Scholar]
- Murelaga, X.; Canudo, J.I. Descripción de los restos de quelonios del Maastrichtiense superior de Aren y Serraduy (Huesca). Geogaceta 2005, 28, 51–54. [Google Scholar]
- Blain, H.A.; Canudo, J.I.; Cuenca-Bescós, G.; López-Martínez, N. Amphibians and squamate reptiles from the latest Maastrichtian (Upper Cretaceous) of Blasi 2 (Huesca, Spain). Cretac. Res. 2010, 31, 433–446. [Google Scholar] [CrossRef]
- Berreteaga, A.; Pereda Suberbiola, X.; Floquet, M.; Olivares, M.; Etxebarria, N.; Iriarte, E.; Badiola Kortabitarte, A.; Elorza, J.; Astibia Ayerra, H. Datos sedimentológicos y tafonómicos de enclaves finicretácicos con fósiles de vertebrados de la Formación Sobrepeña (Burgos, Región Vasco-Cantábrica). Geo-Temas 2008, 10, 1278–1280. [Google Scholar]
- Corral, J.-C.; Pueyo, E.L.; Berreteaga, A.; Rodríguez-Pintó, A.; Sánchez, E.; Pereda-Suberbiola, X. Magnetostratigraphy and lithostratigraphy of the Laño vertebrate-site: Implications in the uppermost Cretaceous chronostratigraphy of the Basque-Cantabrian Region. Cretac. Res. 2016, 57, 473–489. [Google Scholar] [CrossRef]
- Company, J. Vertebrados continentales del Cretácico superior (Campaniense- Maatrichtiense) de Valencia. Ph.D. Thesis, Universidad de Valencia, Valencia, Spain, 2004. [Google Scholar]
- Company, J.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. Last Cretaceous dinosaur faunas from Eastern Iberia into its regional paleogeographic context. Faunal composition and palaeobiogeographical implications. In Proceedings of the IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno Salas de los Infantes, Burgos, Spain, 5–9 September 2009; pp. 17–44. [Google Scholar]
- Laurent, Y.; Bilotte, M.; Le Loeuff, J. Late Maastrichtian continental vertebrates from southwestern France: Correlation with marine fauna. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 187, 121–135. [Google Scholar] [CrossRef]
- Le Loeuff, J.; Buffetaut, E.; Martin, M. The last stages of dinosaur faunal history in Europe: A succession of Maastrichtian dinosaur assemblages from the Corbières (southern France). Geol. Mag. 1994, 131, 625–630. [Google Scholar] [CrossRef]
- Prieto-Márquez, A.; Fondevilla, V.; Sellés, A.G.; Wagner, J.R.; Galobart, À. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European archipelago. Cretac. Res. 2019, 96, 19–37. [Google Scholar] [CrossRef]
- Casanovas-Cladellas, M.L.; Santafé-Llopis, J.V.; Isidro-Llorens, A. Pararhabdodon isonensis n. gen. n. sp. (Dinosauria). Estudio mofológico, radio-tomográfico y consideraciones biomecanicas 26-27:121-131. Paleontol. Evol. 1993, 26–27, 121–132. [Google Scholar]
- Serrano, J.F.; Sellés, A.G.; Vila, B.; Galobart, À.; Prieto-Márquez, A. The osteohistology of new remains of Pararhabdodon isonensis sheds light into the life history and paleoecology of this enigmatic European lambeosaurine dinosaur. Cretac. Res. 2021, 118, 104677. [Google Scholar] [CrossRef]
- Prieto-Marquez, A.; Wagner, J.R. Pararhabdodon isonensis and Tsintaosaurus spinorhinus: A new clade of lambeosaurine hadrosaurids from Eurasia. Cretac. Res. 2009, 30, 1238–1246. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Marquez, A.; Gaete, R.; Rivas, G.; Galobart, À.; Boada, M. Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et sp. nov. J. Vertebr. Paleontol. 2006, 26, 929–943. [Google Scholar] [CrossRef]
- Blanco, A.; Prieto-Márquez, A.; De Esteban-Trivigno, S. Diversity of hadrosauroid dinosaurs from the Late Cretaceous Ibero-Armorican Island (European Archipelago) assessed from dentary morphology. Cretac. Res. 2015, 56, 447–457. [Google Scholar] [CrossRef]
- Fondevilla, V.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À.; Moncunill-Solé, B.; Köhler, M. Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, Tremp Syncline, Spain). PLoS ONE 2018, 13, e0206287. [Google Scholar] [CrossRef]
- Company, J.; Galobart Lorente, À.; Gaete, R. First data on the hadrosaurid dinosaurs (Ornithischia, Dinosauria) from the Upper Cretaceous of Valencia, Spain. Oryctos 1998, 1, 121–126. [Google Scholar]
- Pereda-Suberbiola, X.; Pérez-García, A.; Corral, J.C.; Murelaga, X.; Martin, G.; Larrañaga, J.; Bardet, N.; Berreteaga, A.; Company, J. First dinosaur and turtle remains from the latest Cretaceous shallow marine deposits of Albaina (Laño quarry, Iberian Peninsula). Comptes Rendus Palevol. 2015, 14, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Santafé, J.V.; Casanovas, M.L.; Llompart, C. Els Dinosaures i el Seu Entorn Geològic; Diputació de Lleida: Lleida, Spain, 1997. [Google Scholar]
- López-Martínez, N.; Moratalla, J.J.; Sanz, J.L. Dinosaurs nesting on tidal flats. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 160, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Baiano, M.; Galobart, À.; Dalla, F.M.; Vila, B. Aplicación de la microtomografía en el estudio de dientes aislados de dinosaurios terópodos. In Proceedings of the XII Encuentro de Jóvenes investigadores en Paleontología, Boltaña, Spain, 9–12 April 2014; pp. 57–59. [Google Scholar]
- Marmi, J.; Blanco, A.; Fondevilla, V.; Dalla Vecchia, F.M.; Sellés, A.G.; Vicente, A.; Martín-Closas, C.; Oms, O.; Galobart, À. The Molí del Baró-1 site, a diverse fossil assemblage from the uppermost Maastrichtian of the southern Pyrenees (north-eastern Iberia). Cretac. Res. 2016, 57, 519–539. [Google Scholar] [CrossRef]
- Sellés, A.G.; Vila, B.; Brusatte, S.L.; Currie, P.J.; Galobart, À. A fast-growing basal troodontid (Dinosauria: Theropoda) from the latest Cretaceous of Europe. Sci. Rep. 2021, 11, 4855. [Google Scholar] [CrossRef]
- Sellés, A.G.; Vila, B.; Galobart, À. Diversity of theropod ootaxa and its implications for the latest cretaceous dinosaur turnover in southwestern Europe. Cretac. Res. 2014, 49, 45–54. [Google Scholar] [CrossRef]
- Vianey-Liaud, M.; Lopez-Martinez, N. Late Cretaceous Dinosaur Eggshells from the Tremp Basin, Southern Pyrenees, Lleida, Spain. J. Paleontol. 1997, 71, 1157–1171. [Google Scholar] [CrossRef]
- Laurent, Y. Les faunes de vertébrés continentaux du Maastrichtien supérieur d’Europe: Systématique et biodiversité. Strat Série 2003, 41, 1–81. [Google Scholar]
- Ogg, J.G.; Hinnov, L.A.; Huang, C. Cretaceous. In The Geologic Time Scale; Elsevier: Amsterdam, The Netherlands, 2012; pp. 793–853. [Google Scholar]
- Buffetaut, E.; Clarke, J.B.; Le Loeuff, J. A terminal Cretaceous pterosaur from the Corbieres (southem France) and the problem of pterosaur extinction. Bull. Soc. Geol. Fr. 1996, 167, 753–759. [Google Scholar]
- Buffetaut, E.; Laurent, Y.; Le Loeuff, J.; Bilotte, M. A terminal Cretaceous giant pterosaur from the French Pyrenees. Geol. Mag. 1997, 134, 553–556. [Google Scholar] [CrossRef]
- Buffetaut, E. Late Cretaceous pterosaurs from France: A review. Zitteliana 2008, 28, 249–255. [Google Scholar]
- Company, J.; Ruiz-Omeñaca, J.I.; Suberbiola, X.P. A long-necked pterosaur (Pterodactyloidea, Azhdarchidae) from the Upper Cretaceous of Valencia, Spain. Geol. Mijnbouw/Netherlands J. Geosci. 1999, 78, 319–333. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Company, J.; Ruiz-Omeñaca, J.I. Azhdarchid pterosaurs from the Late Cretaceous (Campanian-Maastrichtian) of the Iberian Peninsula. In Proceedings of the Munich Flugsaurier, The Wellnhofer Pterosaur Meeting, Munich, Germany, 10–14 September 2007; p. 27. [Google Scholar]
- Blanco, A.; Méndez, J.M.; Marmi, J. The fossil record of the uppermost Maastrichtian Reptile Sandstone (Tremp Formation, northeastern Iberian Peninsula). Span. J. Paleontol. 2015, 30, 147–160. [Google Scholar] [CrossRef]
- Laurent, Y.; Buffetaut, E.; Le Loeuff, J. Un crane de thoracosaurine (Crocodylia, Crocodylidae) dans le Maastrichtien Supérieur du sud de la France. Oryctos 2000, 3, 19–27. [Google Scholar]
- Brochu, C.A. A new Late Cretaceous gavialoid crocodylian from eastern North America and the phylogenetic relationships of thoracosaurs. J. Vertebr. Paleontol. 2004, 24, 610–633. [Google Scholar] [CrossRef]
- Le Loeuff, J.; Buffetaut, E.; Cavin, L.; Laurent, Y.; Martin, M.; Martin, V.; Tong, H. Les hadrosaures des Corbières et des Petites Pyrenées. Bull. Soc. Etudes Sci. Aude 1994, 94, 19–21. [Google Scholar]
- Buffetaut, E.; Le Loeuff, J. Late Cretaceous dinosaurs from the foothills of the Pyrenees. Geol. Today 1997, 13, 60–68. [Google Scholar] [CrossRef]
- Tennant, J.P.; Mannion, P.D.; Upchurch, P. Evolutionary relationships and systematics of Atoposauridae (Crocodylomorpha: Neosuchia): Implications for the rise of Eusuchia. Zool. J. Linn. Soc. 2016, 177, 854–936. [Google Scholar] [CrossRef] [Green Version]
- Murelaga, X.; García Garmilla, F.; Pereda Suberbiola, X. Primeros restos de vertebrados del Cretácico Superior de Quecedo de Valdivieso (Burgos). Geogaceta 2005, 195–198. [Google Scholar]
- Berreteaga, A. Estudio Estratigráfico, Sedimentológico y Paleontológico de los Yacimientos con Fósiles de Vertebrados del Cretácico Final de la Región Vasco-Cantábrica. Ph.D. Thesis, University of the Basque Country, Bilbao, Spain, 2008. [Google Scholar]
- Laurent, Y.; Tong, H.; Claude, J. New side-necked turtle (Pleurodira: Bothremydidae) from the Upper Maastrichtian of the Petites-Pyrénées (Haute-Garonne, France). Cretac. Res. 2002, 23, 465–471. [Google Scholar] [CrossRef]
- Marmi, J.; Luján, Á.H.; Riera, V.; Gaete, R.; Oms, O.; Galobart, À. The youngest species of Polysternon: A new bothremydid turtle from the uppermost Maastrichtian of the southern Pyrenees. Cretac. Res. 2012, 35, 133–142. [Google Scholar] [CrossRef]
- Pérez-García, A. Las tortugas mesozoicas de la Peninsula Ibérica. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 2012. [Google Scholar]
- Murelaga, X.; Pereda Suberbiola, X.; Astibia, H.; Lapparent, F.D. Primeros datos sobre los quelonios del Cretácico superior de Lleida. Geogaceta 1998, 24, 239–242. [Google Scholar]
- Blanco, A.; Bolet, A.; Blain, H.-A.; Fondevilla, V.; Marmi, J. Late Cretaceous (Maastrichtian) amphibians and squamates from northeastern Iberia. Cretac. Res. 2016, 57, 624–638. [Google Scholar] [CrossRef]
- Szentesi, Z.; Company, J. Late Maastrichtian small-sized herpetofauna from Valencia province, eastern Spain. Hist. Biol. 2017, 29, 43–52. [Google Scholar] [CrossRef]
- Pol, C.; Buscalioni, A.D.; Carballeira, J.; Francés, V.; López-Martínez, N.; Marandat, B.; Moratalla, J.; Sanz, J.L.; Sigé, B.; Villatte, J. Reptiles and mammals from the Late Cretaceous new locality Quintanilla del Coco (Burgos Province, Spain). Neues Jahrb. Geol. Paläontol. Abh. 1992, 184, 279–314. [Google Scholar]
- Gheerbrant, E.; Astibia, H. Addition to the Late Cretaceous Laño mammal faunule (Spain) and to the knowledge of European “Zhelestidae” (Lainodontinae nov.). Bull. Soc. Géol. Fr. 2012, 183, 537–546. [Google Scholar] [CrossRef]
- Tabuce, R.; Tortosa, T.; Vianey-Liaud, M.; Garcia, G.; Lebrun, R.; Godefroit, P.; Dutour, Y.; Berton, S.; Valentin, X.; Cheylan, G. New eutherian mammals from the Late Cretaceous of Aix-en-Provence Basin, south-eastern France. Zool. J. Linn. Soc. 2013, 169, 653–672. [Google Scholar] [CrossRef]
- Peláez-Campomanes, P.; López-Martínez, N.; Álvarez-Sierra, M.A.; Daams, R. The earliest mammal of the European Paleocene: The multituberculate Hainina. J. Paleontol. 2000, 74, 701–711. [Google Scholar] [CrossRef] [Green Version]
- López-Martínez, N.; Pelaez-Campomanes, P. New mammals from south-central Pyrenees (Tremp Formation, Spain) and their bearing on late Paleocene marine-continental correlations. Bull. Soc. Géol. Fr. 1999, 170, 681–696. [Google Scholar]
- Gheerbrant, E.; Abrial, C.; Cappetta, H. Nouveaux sites a microvertébrés continentaux du Crétacé terminal des Petites Pyrénées (Haute-Garonne et Ariège, France). Geobios 1997, 30, 257–269. [Google Scholar] [CrossRef]
- Nopcsa, F. Über das Vorkommen der Dinosaurier in Siebenbürgen. Verhandlungen der Zool Gesellschaft 1914, 54, 12–14. [Google Scholar]
- Benton, M.J.; Csiki, Z.; Grigorescu, D.; Redelstorff, R.; Sander, P.M.; Stein, K.; Weishampel, D.B. Dinosaurs and the island rule: The dwarfed dinosaurs from Haţeg Island. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 293, 438–454. [Google Scholar] [CrossRef]
- Godefroit, P.; Codrea, V.; Weishampel, D.B. Osteology of Zalmoxes shqiperorum (Dinosauria, Ornithopoda), based on new specimens from the Upper Cretaceous of Nălaţ-Vad (Romania). Geodiversitas 2009, 31, 525–553. [Google Scholar] [CrossRef]
- Weishampel, D.B.; Jianu, C.; Csiki, Z.; Norman, D.B. Osteology and phylogeny of Zalmoxes (n. g.), an unusual Euornithopod dinosaur from the latest Cretaceous of Romania. J. Syst. Palaeontol. 2003, 1, 65–123. [Google Scholar] [CrossRef]
- Weishampel, D.B.; Norman, D.B.; Grigorescu, D. Telmatosaurus transsylvanicus from the late Cretaceous of Romania: The most basal hadrosaurid dinosaur. Palaeontology 1993, 36, 361–385. [Google Scholar]
- Dalla Vecchia, F.M. An overview of the latest Cretaceous hadrosauroid record in Europe. In Hadrosaurs; Eberth, D.A., Evans, D.C., Eds.; Indiana University Press: Bloomington, IN, USA, 2014; pp. 268–297. [Google Scholar]
- Codrea, V.; Smith, T.; Dica, P.; Folie, A.; Garcia, G.; Godefroit, P.; Van Itterbeeck, J. Dinosaur egg nests, mammals and other vertebrates from a new Maastrichtian site of the Haţeg Basin (Romania). Comptes Rendus Palevol 2002, 1, 173–180. [Google Scholar] [CrossRef]
- Vasile, S.; Csiki, Z.; Grigorescu, D. The first report of continental fossil remains from Crǎguiş (Haţeg Basin, Romania), and their stratigraphical significance. In Proceedings of the Eighth Romanian Symposium on Paleontology, Bucharest, Romania, 29–30 September 2011; pp. 127–128. [Google Scholar]
- Ősi, A.; Codrea, V.; Prondvai, E.; Csiki-Sava, Z. New ankylosaurian material from the Upper Cretaceous of Transylvania. Ann. Paléontol. 2014, 100, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Nopcsa, F. Die dinosaurier der siebenburgischen landesteile ungarns. Mitteilungen aus dem Jahrb. der königlich Ung. Geol. Reichsanstalt 1915, 23, 3–24. [Google Scholar]
- Csiki, Z.; Codrea, V.; Jipa-Murzea, C.; Godefroit, P. A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nalat-Vad, Hateg Basin, Romania. Neues Jahrb. Geol. Paläontol. Abhandlungen 2010, 258, 297–324. [Google Scholar] [CrossRef]
- Codrea, V.; Vremir, M.; Jipa, C.; Godefroit, P.; Csiki, Z.; Smith, T.; Fărcaş, C. More than just Nopcsa’s Transylvanian dinosaurs: A look outside the Haţeg Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 293, 391–405. [Google Scholar] [CrossRef]
- Vremir, M.M. New faunal elements from the late Cretaceous (Maastrichtian) continental deposits of Sebeş area (Transylvania). Terra Sebus 2010, 2, 635–684. [Google Scholar]
- Csiki, Z.; Vremir, M. A large-sized (?)Late Maastrichtian titanosaur from Rapa Rosie, Sebes. In Proceedings of the 8th Romanian Symposium of Paleontology, Bucharest, Romania, 29–30 September 2011; pp. 28–29. [Google Scholar]
- Mannion, P.D.; Díez Díaz, V.; Csiki-Sava, Z.; Upchurch, P.; Cuff, A.R. Dwarfs among giants: Resolving the systematics of the titanosaurian sauropod dinosaurs from the Late Cretaceous of Romania. In Proceedings of the XVII Conference of the EAVP, Brussels, Belgium, 1–6 July 2019; p. 65. [Google Scholar]
- Erickson, G.M.; Krick, B.A.; Hamilton, M.; Bourne, G.R.; Norell, M.A.; Lilleodden, E.; Sawyer, W.G. Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science 2012, 338, 98–101. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, A.R.H.; Reisz, R.R.; Evans, D.C.; Bailleul, A.M. Ontogeny reveals function and evolution of the hadrosaurid dinosaur dental battery. BMC Evol. Biol. 2016, 16, 152. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.; Feldmann, R.M.; Tashman, J.N. Consumption of crustaceans by megaherbivorous dinosaurs: Dietary flexibility and dinosaur life history strategies. Sci. Rep. 2017, 7, 11163. [Google Scholar] [CrossRef]
- Vremir, M.; Dyke, G.; Totoianu, R. Repertoire of the late cretaceous vertebrate localities from Sebescedil; area, Alba county (Romania). Terra Sebus 2015, 7, 695–724. [Google Scholar]
- Văcărescu, M.R.; Sava, Z.C.; Bucur, M.; Roman, A.; Vasile, Ș. Theropod dental diversity in the Maastrichtian of the Hațeg Basin, Romania Hațeg Basin have yielded a diverse and unique assemblage of vertebrate. In Proceedings of the 11th Romanian Symposium of Paleontology, Bucharest, Romania, 27–28 September 2017; pp. 122–123. [Google Scholar]
- Brusatte, S.L.; Vremir, M.; Csiki-Sava, Z.; Turner, A.H.; Watanabe, A.; Erickson, G.M.; Norell, M.A. The Osteology of Balaur bondoc, an Island-Dwelling Dromaeosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Romania. Bull. Am. Museum Nat. Hist. 2013, 374, 1–100. [Google Scholar] [CrossRef]
- Csiki, Z.; Vremir, M.; Brusatte, S.L.; Norell, M.A. An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. Proc. Natl. Acad. Sci. USA 2010, 107, 15357–15361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Dyke, G.J.; Codrea, V.; Godefroit, P.; Smith, T. A Euenantiornithine Bird from the Late Cretaceous Ha eg Basin of Romania. Acta Palaeontol. Pol. 2011, 56, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Mayr, G.; Codrea, V.; Solomon, A.; Bordeianu, M.; Smith, T. A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic. Cretac. Res. 2020, 106, 104271. [Google Scholar] [CrossRef]
- Sallam, H.M.; Gorscak, E.; O’Connor, P.M.; El-Dawoudi, I.A.; El-Sayed, S.; Saber, S.; Kora, M.A.; Sertich, J.J.W.; Seiffert, E.R.; Lamanna, M.C. New Egyptian sauropod reveals Late Cretaceous dinosaur dispersal between Europe and Africa. Nat. Ecol. Evol. 2018, 2, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, T.; Buffetaut, E.; Vialle, N.; Dutour, Y.; Turini, E.; Cheylan, G. A new abelisaurid dinosaur from the Late Cretaceous of southern France: Palaeobiogeographical implications. Ann. Paléontol. 2014, 100, 63–86. [Google Scholar] [CrossRef]
- Buffetaut, E.; Grigorescu, D.; Csiki, Z. A new giant pterosaur with a robust skull from the latest Cretaceous of Romania. Naturwissenschaften 2002, 89, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Naish, D.; Witton, M.P. Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators. Peer J. 2017, 5, e2908. [Google Scholar] [CrossRef] [Green Version]
- Solomon, A.A.; Codrea, V.A.; Venczel, M.; Grellet-Tinner, G. A new species of large-sized pterosaur from the Maastrichtian of Transylvania (Romania). Cretac. Res. 2020, 110, 104316. [Google Scholar] [CrossRef]
- Rabi, M.; Delfino, M. A reassessment of the “alligatoroid” eusuchian from the Late Cretaceous of Hungary and its taxonomic implications. In Proceedings of the 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; pp. 203–206. [Google Scholar]
- Martin, J.E.; Rabi, M.; Csiki, Z. Survival of Theriosuchus (Mesoeucrocodylia: Atoposauridae) in a Late Cretaceous archipelago: A new species from the Maastrichtian of Romania. Naturwissenschaften 2010, 97, 845–854. [Google Scholar] [CrossRef]
- Martin, J.E.; Rabi, M.; Csiki-Sava, Z.; Vasile, Ş. Cranial morphology of Theriosuchus sympiestodon (Mesoeucrocodylia, Atoposauridae) and the widespread occurrence of Theriosuchus in the Late Cretaceous of Europe. J. Paleontol. 2014, 88, 444–456. [Google Scholar] [CrossRef]
- Rabi, M.; Sebők, N. A revised Eurogondwana model: Late Cretaceous notosuchian crocodyliforms and other vertebrate taxa suggest the retention of episodic faunal links between Europe and Gondwana during most of the Cretaceous. Gondwana Res. 2015, 28, 1197–1211. [Google Scholar] [CrossRef]
- Sellés, A.G.; Blanco, A.; Vila, B.; Marmi, J.; López-Soriano, F.J.; Llácer, S.; Frigola, J.; Canals, M.; Galobart, À. A small Cretaceous crocodyliform in a dinosaur nesting ground and the origin of sebecids. Sci. Rep. 2020, 10, 15293. [Google Scholar] [CrossRef]
- Rabi, M.; Vremir, M.; Tong, H. Preliminary Overview of Late Cretaceous Turtle Diversity in Eastern Central Europe (Austria, Hungary, and Romania). In Morphology and Evolution of Turtles. Vertebrate Paleobiology and Paleoanthropology; Brinkman, D., Holroyd, P., Gardner, J., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 307–336. [Google Scholar]
- Pérez-García, A.; Codrea, V. New insights on the anatomy and systematics of Kallokibotion Nopcsa, 1923, the enigmatic uppermost Cretaceous basal turtle (stem Testudines) from Transylvania. Zool. J. Linn. Soc. 2018, 182, 419–443. [Google Scholar] [CrossRef]
- Venczel, M.; Gardner, J.D.; Codrea, V.A.; Csiki-Sava, Z.; Vasile, Ş.; Solomon, A.A. New insights into Europe’s most diverse Late Cretaceous anuran assemblage from the Maastrichtian of western Romania. Palaeobiodivers. Palaeoenviron. 2016, 96, 61–95. [Google Scholar] [CrossRef]
- Grigorescu, D.; Venczel, M.; Csiki, Z.; Limberea, R. New microvertebrate fossil assemblages from the Uppermost Cretaceous of the Haţeg Basin (Romania). Geol. Mijnb. 1999, 78, 301–314. [Google Scholar] [CrossRef]
- Folie, A.; Codrea, V. New lissamphibians and squamates from the Maastrichtian of Haţeg Basin, Romania. Acta Palaeontol. Pol. 2005, 50, 57–71. [Google Scholar]
- Codrea, V.A.; Venczel, M.; Solomon, A. A new family of teiioid lizards from the Upper Cretaceous of Romania with notes on the evolutionary history of early teiioids. Zool. J. Linn. Soc. 2017. [Google Scholar] [CrossRef]
- Vasile, Ş.; Csiki-Sava, Z.; Venczel, M. A new madtsoiid snake from the Upper Cretaceous of the Haţeg Basin, western Romania. J. Vertebr. Paleontol. 2013, 33, 1100–1119. [Google Scholar] [CrossRef]
- Venczel, M.; Vasile, Ş.; Csiki-Sava, Z. A Late Cretaceous madtsoiid snake from Romania associated with a megaloolithid egg nest—Paleoecological inferences. Cretac. Res. 2015, 55, 152–163. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Corral, J.C.; Astibia, H.; Badiola, A.; Bardet, N.; Berreteaga, A.; Buffetaut, E.; Buscalioni, A.D.; Cappetta, H.; Cavin, L.; et al. Late Cretaceous continental and marine vertebrate assemblages from the Laño quarry (Basque-Cantabrian Region, Iberian Peninsula): An update. J. Iber. Geol. 2015, 41. [Google Scholar] [CrossRef] [Green Version]
- Csiki, Z.; Grigorescu, D.; Rücklin, M. A new multituberculate specimen from the Maastrichtian of Pui, Romania and a reassessment of affinities of Barbatodon. Acta Palaeontol. Rom. 2005, 5, 73–86. [Google Scholar]
- Smith, T.; Codrea, V. Red iron-pigmented tooth enamel in a multituberculate mammal from the Late Cretaceous Transylvanian “Haţeg Island”. PLoS ONE 2015, 10, e0132550. [Google Scholar] [CrossRef]
- Csiki-Sava, Z.; Vremir, M.; Meng, J.; Brusatte, S.L.; Norell, M.A. Dome-headed, small-brained island mammal from the Late Cretaceous of Romania. Proc. Natl. Acad. Sci. USA 2018, 115, 4857–4862. [Google Scholar] [CrossRef] [Green Version]
- De Bast, E.; Smith, T. The oldest Cenozoic mammal fauna of Europe: Implication of the Hainin reference fauna for mammalian evolution and dispersals during the Paleocene. J. Syst. Palaeontol. 2017, 15, 741–785. [Google Scholar] [CrossRef]
- Brocklehurst, N.; Kammerer, C.F.; Fröbisch, J. The early evolution of synapsids, and the influence of sampling on their fossil record. Paleobiology 2013, 39, 470–490. [Google Scholar] [CrossRef]
- Purnell, M.A.; Donoghue, P.C.J. Between death and data: Biases in interpretation of the fossil record of conodonts—The Palaeontological Association. Spec. Pap. Palaeontol. 2005, 25, 7–25. [Google Scholar]
- Benton, M.J.; Dunhill, A.M.; Lloyd, G.T.; Marx, F.G. Assessing the quality of the fossil record: Insights from vertebrates. Geol. Soc. Lond. Spec. Publ. 2011, 358, 63–94. [Google Scholar] [CrossRef] [Green Version]
- Benton, M.J.; Pearson, P.N. Speciation in the fossil record. Trends Ecol. Evol. 2001, 16, 405–411. [Google Scholar] [CrossRef]
- Grigorescu, D.; Csiki-Sava, Z.; Vasile, Ş.; Butiseacă, G.-A. Taphonomic biases in macro- and microvertebrate assemblages from the Maastrichtian of the Hațeg Basin (Romania) and their relevance in the reconstruction of a fossil ecosystem. In Proceedings of the 9th Annual Meeting of the European Association of Vertebrate Palaeontologists, Heraklion, Greece, 14–19 June 2011; pp. 28–29. [Google Scholar]
- Chiarenza, A.A.; Mannion, P.D.; Lunt, D.J.; Farnsworth, A.; Jones, L.A.; Kelland, S.-J.; Allison, P.A. Ecological niche modelling does not support climatically-driven dinosaur diversity decline before the Cretaceous/Paleogene mass extinction. Nat. Commun. 2019, 10, 1091. [Google Scholar] [CrossRef] [Green Version]
- Teixell, A.; Labaume, P.; Lagabrielle, Y. The crustal evolution of the west-central Pyrenees revisited: Inferences from a new kinematic scenario. Comptes Rendus Geosci. 2016, 348, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Fondevilla, V.; Vila, B.; Galobart, À.; Oms, O. Taphonomic modes in the dinosaur-bearing Tremp Formation (Maastrichtian, Southern Pyrenees). In Proceedings of the XIV Annual Meeting of the European Association of Vertebrate Palaeontologists, Haarlem, The Netherlands, 6–10 July 2016; p. 223. [Google Scholar]
- Upchurch, P.; Mannion, P.D.; Benson, R.B.J.; Butler, R.J.; Carrano, M.T. Geological and anthropogenic controls on the sampling of the terrestrial fossil record: A case study from the Dinosauria. Geol. Soc. Lond. Spec. Publ. 2011, 358, 209–240. [Google Scholar] [CrossRef]
- Le Loeuff, J. Paleobiogeography and biodiversity of Late Maastrichtian dinosaurs: How many dinosaur species went extinct at the Cretaceous-Tertiary boundary? Bull. Soc. Géol. Fr. 2012, 183, 547–559. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Ibero-Armorican Island | Western Tremp Syncline | Hațeg Island |
|---|---|---|---|
| Amphibia | 6 | 3 | 4 |
| Anura | 4 | 2 | 3 |
| Albanerpetontidae | 1 | 1 | 1 |
| Salamandridae | 1 | − | − |
| Squamata | 6 + 2? | 3 + 2? | 5 |
| ‘Scincomorpha’ | 1 | 1? | 1 |
| Anguimorpha | 1 | 1 | 1 |
| Teiioidea | 1 | − | 1 |
| Borioteiioidea | − | − | 1 |
| Scleroglossa | 1 | 1 | − |
| Iguanidae | 1 | 1? | − |
| Amphisbaenia | 1? | − | − |
| Varanoidea | 1? | − | − |
| Alethinophidia | 1 | 1 | 1 |
| Testudines | 5 | 2 | 2 |
| Meiolaniformes | − | − | 1 |
| Pan-Pleurodira | 3 | 1 | 1 |
| Pan-Cryptodira | 2 | 1 | − |
| Crocodylomorpha | 8 | 4 | 4 |
| Notosuchia (Doratodon) | 1 | − | 1 |
| Neosuchia (‘Atoposauridae’) | 1 | - | 1 |
| Basal Eusuchia (Allodaposuchidae) | 4 | 2 | 1 |
| Basal Eusuchia (Hylaeochampsidae, cf. Acynodon) | 1 | 1 | 1 |
| Eusuchia (Gavialoidea) | 1 | 1 | − |
| Pterosauria | 1 | 1 | 2 |
| Azhdarchidae | 1 | 1 | 2 |
| Dinosauria | 18 + 1? (6) | 10 (1) | 12 + 1? (3) |
| Sauropoda | 3 | 1 | 3 |
| Titanosauria | 3 | 1 | 3 |
| Theropoda | 8 + 1? | 6 | 6 + 1? (3) |
| Alvarezsauridae | − | - | 1? |
| Abelisauroidea | 1 | 1 | − |
| Coelurosauria indet. | 1 | 1 | − |
| Maniraptora | 5 | 3 | 3 (2) |
| Paraves with uncertain affinities (Balaur, Elopteryx) | − | − | 1 (1) |
| Enantiornithes | 1? | − | 1 |
| Ornithuromorpha | 1 | 1 | 1 |
| Ornithopoda | 6 (6) | 3 (1) | 2 |
| Rhabdodontidae | − | − | 1 |
| Hadrosauroidea | 6 (6) | 3 (1) | 1 |
| Ankylosauria | 1 | - | 1 |
| Nodosauridae | 1 | − | 1 |
| Mammalia | 1? | 3 | |
| Multituberculata | − | − | 3 |
| Theria | 1? | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Pueyo, M.; Cruzado-Caballero, P.; Moreno-Azanza, M.; Vila, B.; Castanera, D.; Gasca, J.M.; Puértolas-Pascual, E.; Bádenas, B.; Canudo, J.I. The Tetrapod Fossil Record from the Uppermost Maastrichtian of the Ibero-Armorican Island: An Integrative Review Based on the Outcrops of the Western Tremp Syncline (Aragón, Huesca Province, NE Spain). Geosciences 2021, 11, 162. https://doi.org/10.3390/geosciences11040162
Pérez-Pueyo M, Cruzado-Caballero P, Moreno-Azanza M, Vila B, Castanera D, Gasca JM, Puértolas-Pascual E, Bádenas B, Canudo JI. The Tetrapod Fossil Record from the Uppermost Maastrichtian of the Ibero-Armorican Island: An Integrative Review Based on the Outcrops of the Western Tremp Syncline (Aragón, Huesca Province, NE Spain). Geosciences. 2021; 11(4):162. https://doi.org/10.3390/geosciences11040162
Chicago/Turabian StylePérez-Pueyo, Manuel, Penélope Cruzado-Caballero, Miguel Moreno-Azanza, Bernat Vila, Diego Castanera, José Manuel Gasca, Eduardo Puértolas-Pascual, Beatriz Bádenas, and José Ignacio Canudo. 2021. "The Tetrapod Fossil Record from the Uppermost Maastrichtian of the Ibero-Armorican Island: An Integrative Review Based on the Outcrops of the Western Tremp Syncline (Aragón, Huesca Province, NE Spain)" Geosciences 11, no. 4: 162. https://doi.org/10.3390/geosciences11040162
APA StylePérez-Pueyo, M., Cruzado-Caballero, P., Moreno-Azanza, M., Vila, B., Castanera, D., Gasca, J. M., Puértolas-Pascual, E., Bádenas, B., & Canudo, J. I. (2021). The Tetrapod Fossil Record from the Uppermost Maastrichtian of the Ibero-Armorican Island: An Integrative Review Based on the Outcrops of the Western Tremp Syncline (Aragón, Huesca Province, NE Spain). Geosciences, 11(4), 162. https://doi.org/10.3390/geosciences11040162