
Potential Use of Biochar as a Mitigation Strategy for Salinity-Related Issues in Tomato Plants (Solanum lycopersicum L.)

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochar Production and Characterization
2.2. Plant Growth
2.3. Determination of the Concentration of Macro- and Micronutrients in Tomato Plants
2.4. Determination of Malondialdehyde Concentration
2.5. Determination of Non-Protein Thiol Concentration
2.6. Extraction and Determination of Guaiacol Peroxidase (GPX) Activity
2.7. Other Measurements
2.8. Statistical Analysis
3. Results
3.1. Biochar Characterization
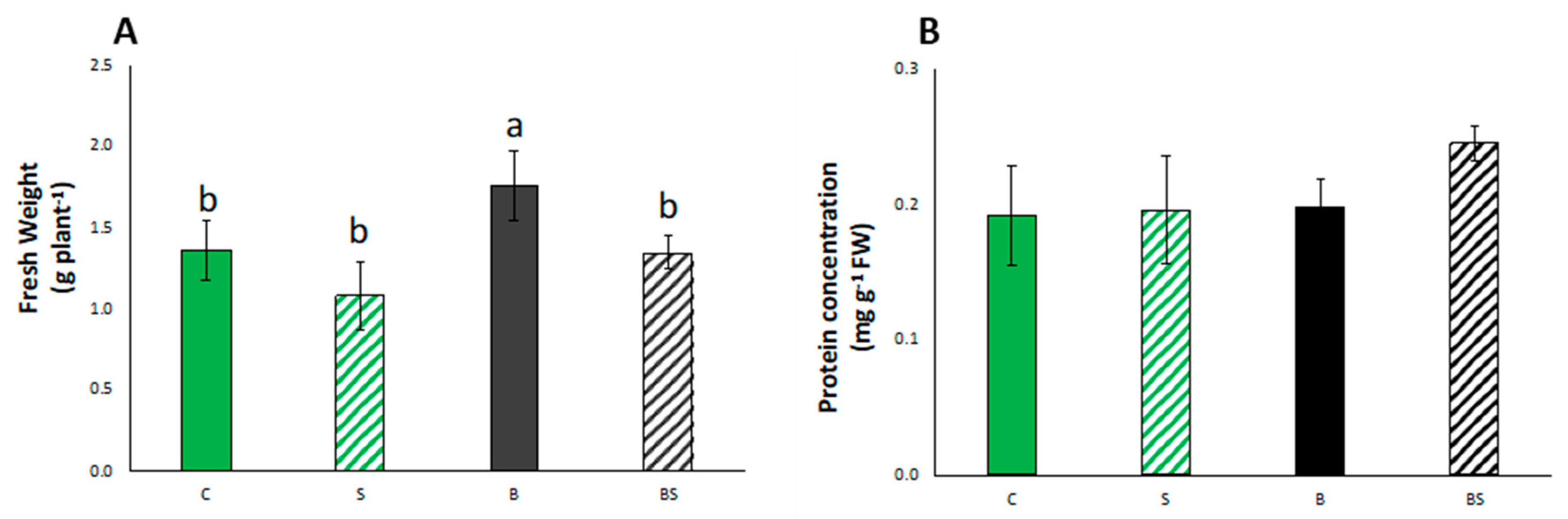
3.2. Plant Growth Parameters
3.3. Lipid Peroxidation
3.4. Non-Enzymatic and Enzymatic Antioxidant Response
3.5. Shoot Elemental Composition
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC. 2023: Summary for Policymakers. In Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; pp. 1–34. [Google Scholar]
- Atwoli, L.; Erhabor, G.E.; Gbakima, A.A.; Haileamlak, A.; Ntumba, J.-M.K.; Kigera, J.; Laybourn-Langton, L.; Mash, R.; Muhia, J.; Mulaudzi, F.M.; et al. COP27 Climate Change Conference: Urgent action needed for Africa and the world. Rev. Panam. Salud Publica 2022, 23, 1486–1488. [Google Scholar]
- Misra, A.K. Climate change and challenges of water and food security. Int. J. Sustain. Built Environ. 2014, 3, 153–165. [Google Scholar] [CrossRef]
- Scholes, R.J. The Future of Semi-Arid Regions: A Weak Fabric Unravels. Climate 2020, 8, 43. [Google Scholar] [CrossRef]
- Velmurugan, A.; Swarnam, P.; Subramani, T.; Meena, B.; Kaledhonkar, M.J. Water demand and salinity. In Desalination-Challenges and Opportunities, 1st ed.; Farahani, M.H.D.A., Vatanpour, V., Taheri, A., Eds.; IntechOpen: London, UK, 2020; pp. 1–11. [Google Scholar]
- Steduto, P.; Faurès, J.-M.; Hoogeveen, J.; Winpenny, J.; Burke, J. Coping with Water Scarcity, an Action Framework for Agricultural and Food Security; FAO: Rome, Italy, 2012. [Google Scholar]
- Yermiyahu, U.; Tal, A.; Ben-Gal, A.; Bar-Tal, A.; Tarchitzky, J.; Lahav, O. Rethinking desalinated water quality and agriculture. Science 2007, 318, 920–921. [Google Scholar] [CrossRef]
- Javeed, H.M.R.; Wang, X.; Ali, M.; Nawaz, F.; Qamar, R.; Rehman, A.U.; Shehzad, M.; Mubeen, M.; Shabbir, R.; Javed, T.; et al. Potential Utilization of Diluted Seawater for the Cultivation of Some Summer Vegetable Crops: Physiological and Nutritional Implications. Agronomy 2021, 11, 1826. [Google Scholar] [CrossRef]
- De Pascale, S.; Maggio, A.; Fogliano, V.; Ambrosino, P.; Ritieni, A. Irrigation with Saline Water Improves Carotenoids Content and Antioxidant Activity of Tomato. J. Hortic. Sci. Biotechnol. 2001, 76, 447–453. [Google Scholar] [CrossRef]
- Atzori, G.; Mancuso, S.; Masi, E. Seawater Potential Use in Soilless Culture: A Review. Sci. Hortic. 2019, 249, 199–207. [Google Scholar] [CrossRef]
- Sakamoto, K.; Kogi, M.; Yanagisawa, T. Effects of salinity and nutrients in seawater on hydroponic culture of red leaf lettuce. Environ. Control Biol. 2014, 52, 189–195. [Google Scholar] [CrossRef]
- Turhan, A.; Kuscu, H.; Ozmen, N.; Sitki Serbeci, M.; Osman Demir, A. Effect of different concentrations of diluted seawater on yield and quality of lettuce. Chil. J. Agric. Res. 2014, 74, 111–116. [Google Scholar] [CrossRef]
- Sgherri, C.; Kadlecová, Z.; Pardossi, A.; Navari-Izzo, F.; Izzo, R. Irrigation with diluted seawater improves the nutritional value of cherry tomatoes. J. Agric. Food Chem. 2008, 56, 3391–3397. [Google Scholar] [CrossRef]
- Sgherri, C.; Navari-Izzo, F.; Pardossi, A.; Soressi, G.P.; Izzo, R. The influence of diluted seawater and ripening stage on the content of antioxidants in fruits of different tomato genotypes. J. Agric. Food Chem. 2007, 55, 2452–2458. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.O.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef]
- Saifullah; Dahlawi, S.; Naeem, A.; Rengel, Z.; Naidu, R. Biochar application for the remediation of salt-affected soils: Challenges and opportunities. Sci. Total Environ. 2018, 625, 320–335. [Google Scholar] [CrossRef]
- Yu, H.; Zou, W.; Chen, J.; Chen, H.; Yu, Z.; Huang, J.; Tang, H.; Wei, X.; Gao, B. Biochar amendment improves crop production in problem soils: A review. J. Environ. Manag. 2019, 232, 8–21. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Biochar mitigates salinity stress in potato. J. Agron. Crop. Sci. 2015, 201, 368–378. [Google Scholar] [CrossRef]
- Kul, R.; Arjumend, T.; Ekinci, M.; Yildirim, E.; Turan, M.; Argin, S. Biochar as an organic soil conditioner for mitigating salinity stress in tomato. Soil Sci. Plant Nutr. 2021, 67, 693–706. [Google Scholar] [CrossRef]
- Lashari, M.S.; Liu, Y.; Li, L.; Pan, W.; Fu, J.; Pan, G.; Zheng, J.; Zheng, J.; Zhang, X.; Yu, X. Effects of amendment of biochar-manure compost in conjunction with pyroligneous solution on soil quality and wheat yield of a salt-stressed cropland from Central China Great Plain. Field Crop. Res. 2013, 144, 113–118. [Google Scholar] [CrossRef]
- Thomas, S.C.; Frye, S.; Gale, N.; Garmon, M.; Launchbury, R.; Machado, N.; Melamed, S.; Murray, J.; Petroff, A.; Winsborough, C. Biochar mitigates negative effects of salt additions on two herbaceous plant species. J. Environ. Manag. 2013, 129, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Yaashikaa, P.R.; Senthil Kumar, P.; Varjani, S.; Saravanan, A. A critical review on the biochar production techniques, characterization, stability and applications for circular bioeconomy. Biotechnol. Rep. 2020, 28, e00570. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J. Biochar for mitigating climate change: Carbon sequestration in the black. Forum Geoöekologie 2007, 18, 15–17. [Google Scholar]
- Garcia, B.; Alves, O.; Rijo, B.; Lourinho, G.; Nobre, C. Biochar: Production, applications, and market prospects in Portugal. Environments 2022, 9, 95. [Google Scholar] [CrossRef]
- Picchio, R.; Pari, L.; Venanzi, R.; Latterini, F.; Suardi, A.; Alfano, V.; Bergonzoli, S. A new mobile kiln prototype for charcoal production. In Proceedings of the 28th European Biomass Conference and Exhibition, Virtual, 6–9 July 2020; pp. 686–689. [Google Scholar]
- Al, M.I. Salt Tolerance of Some Tomato (Solanum lycoversicum L.) Cultivars for Salinity under Controlled Conditions. Am. J. Plant Physiol. 2018, 13, 58–64. [Google Scholar]
- Zaki, H.E.M.; Yokoi, S. A Comparative In Vitro Study of Salt Tolerance in Cultivated Tomato and Related Wild Species. Plant Biotechnol. 2016, 33, 361–372. [Google Scholar] [CrossRef]
- Roșca, M.; Mihalache, G.; Stoleru, V. Tomato Responses to Salinity Stress: From Morphological Traits to Genetic Changes. Front. Plant Sci. 2023, 14, 1118383. [Google Scholar] [CrossRef]
- Picchio, R. Manuale di campo sulla carbonizzazione con sistemi a pirolisi lenta. In Field Handbook on the Wood Charbonization with Slow Pyrolysis Systems, in Italian; Picchio, R., Ed.; Bioactam: Viterbo, Italy, 2023; 20p, ISBN 979-12-200-5330-3. [Google Scholar]
- Janiszewska, D.; Olchowski, R.; Nowicka, A.; Zborowska, M.; Marszałkiewicz, K.; Shams, M.; Giannakoudakis, D.A.; Anastopoulos, I. Activated biochars derived from wood biomass liquefaction residues for effective removal of hazardous hexavalent chromium from aquatic environments. GCB Bioenergy 2021, 13, 1247–1259. [Google Scholar] [CrossRef]
- ISO 17225-4; Solid Biofuels—Fuel Specifications and Classes. International Organization for Standardization: Geneva, Switzerland, 2021.
- ISO 18125: 2018; Solid Biofuels—Determination of Heating Value. International Organization for Standardization: Geneva, Switzerland, 2018.
- ISO 17828: 2016; Solid Biofuels—Determination of Bulk Density. International Organization for Standardization: Geneva, Switzerland, 2016.
- ISO 18134-1: 2015; Solid Biofuels—Determination of Moisture Content—Oven Dry Method. International Organization for Standardization: Geneva, Switzerland, 2015.
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on Ignition as a Method for Estimating Organic and Carbonate Content in Sediments: Reproducibility and Comparability of Results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Blue, C.R.; Rimstidt, J.D.; Dove, P.M. A Mixed Flow Reactor Method to Synthesize Amorphous Calcium Carbonate under Controlled Chemical Conditions. Methods Enzymol. 2013, 532, 557–568. [Google Scholar] [PubMed]
- Mehlich, A. Use of triethanolamine acetate-barium hydroxide buffer for the determination of some base exchange properties and lime requirement of soil. Soil Sci. Soc. Am. Proc. 1938, 29, 374–378. [Google Scholar] [CrossRef]
- Zuchi, S.; Cesco, S.; Varanini, Z.; Pinton, R.; Astolfi, S. Sulphur deprivation limits Fe-deficiency responses in tomato plants. Planta 2009, 230, 85–94. [Google Scholar] [CrossRef]
- Quagliata, G.; Celletti, S.; Coppa, E.; Mimmo, T.; Cesco, S.; Astolfi, S. Potential Use of Copper-Contaminated Soils for Hemp (Cannabis sativa L.). Cultivation. Environ. 2021, 8, 111. [Google Scholar] [CrossRef]
- Silvestri, C.; Celletti, S.; Cristofori, V.; Astolfi, S.; Ruggiero, B.; Rugini, E. Olive (Olea europaea L.) plants transgenic for tobacco osmotin gene are less sensitive to in vitro-induced drought stress. Acta Physiol. Plant. 2017, 39, 229. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Florence, T.M.; Farrar, Y.T. Spectrophotometric determination of chloride at the parts-per-billion level by the mercury (II) thiocyanate method. Anal. Chim. Acta 1971, 54, 373–377. [Google Scholar] [CrossRef]
- Askeland, M.; Clarke, B.; Paz-Ferreiro, J. Comparative characterization of biochars produced at three selected pyrolysis temperatures from common woody and herbaceous waste streams. PeerJ 2019, 7, e6784. [Google Scholar] [CrossRef]
- Das, O.; Sarmah, A.K. The love–hate relationship of pyrolysis biochar and water: A perspective. Sci. Total Environ. 2015, 512, 682–685. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Sánchez, M.E.; Mora, M.; Barrón, V. Slow pyrolysis of relevant biomasses in the Mediterranean basin. Part 2. Char characterisation for carbon sequestration and agricultural uses. J. Clean. Prod. 2016, 120, 191–197. [Google Scholar] [CrossRef]
- USGS. Science for a Changing World. Available online: https://www.usgs.gov/special-topics/water-science-school/science/why-ocean-salty (accessed on 6 July 2023).
- IBI-STD-01.1; Standardized Product Definition and Product Testing Guidelines for Biochar That Is Used in Soil. Version 2.1. International Biochar Initiative (IBI): Canandaigua, NY, USA, 2013.
- Ferraro, G.; Pecori, G.; Rosi, L.; Bettucci, L.; Fratini, E.; Casini, D.; Rizzo, A.M.; Chiaramonti, D. Biochar from lab-scale pyrolysis: Influence of feedstock and operational temperature. Biomass Convers. Biorefinery 2021, 1–11. [Google Scholar] [CrossRef]
- Tank, N.; Saraf, M. Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J. Plant Interact. 2010, 5, 51–58. [Google Scholar] [CrossRef]
- Benavente, I.; Gasco, G.; Plaza, C.; Paz-Ferreiro, J.; Mendez, A. Choice of pyrolysis parameters for urban wastes affects soil enzymes and plant germination in a Mediterranean soil. Sci. Total Environ. 2018, 634, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; O’Connor, D.; Zhang, J.; Peng, T.; Shen, Z.; Tsang, D.C.W.; Hou, D. Effect of Pyrolysis Temperature, Heating Rate, and Residence Time on Rapeseed Stem Derived Biochar. J. Clean. Prod. 2018, 174, 977–987. [Google Scholar] [CrossRef]
- IBI-STD-2.1; Standardized Product Definition and Product Testing Guidelines for Biochar that Is Used in Soil. Version 2.1. International Biochar Initiative (IBI): Canandaigua, NY, USA, 2015.
- Hilber, P.; Mayer, V.; Gouliarmou, S.E.; Hale, G.; Cornelissen, H.P.; Schmidt, T.D. Bucheli Bioavailability and bioaccessibility of polycyclic aromatic hydrocarbons from (post-pyrolytically treated) biochars. Chemosphere 2017, 174, 700–707. [Google Scholar] [CrossRef]
- Ghani, W.A.W.A.K.; Mohd, A.; de Silva, G.; Bachmann, R.T.; Taufiq-Yap, Y.H.; Rashid, U.; Al-Muhtaseb, A.H. Biochar production from waste rubber-wood-sawdust and its potential use in C sequestration: Chemical and physical characterization. Ind. Crop. Prod. 2013, 44, 18–24. [Google Scholar] [CrossRef]
- José, M.; Sánchez-Martín, Á.M.; Campos, P.; Miller, A.Z. Effect of pyrolysis conditions on the total contents of polycyclic aromatic hydrocarbons in biochars produced from organic residues: Assessment of their hazard potential. Sci. Total Environ. 2019, 667, 578–585. [Google Scholar]
- Yao, Y.; Gao, B.; Zhang, M.; Inyang, M.; Zimmerman, A.R. Effect of Biochar Amendment on Sorption and Leaching of Nitrate, Ammonium, and Phosphate in a Sandy Soil. Chemosphere 2012, 89, 1467–1471. [Google Scholar] [CrossRef]
- Quratulain Shafiq, M.; Capareda, S.C. Firdaus-e-Bareen Effect of Different Temperatures on the Properties of Pyrolysis Products of Parthenium Hysterophorus. J. Saudi Chem. Soc. 2021, 25, 101197. [Google Scholar] [CrossRef]
- Glaser, B.; Lehr, V.I. Biochar effects on phosphorus availability in agricultural soils: A meta-analysis. Sci. Rep. 2019, 9, 9338. [Google Scholar] [CrossRef]
- Gao, S.; DeLuca, T.H.; Cleveland, C.C. Biochar additions alter phosphorus and nitrogen availability in agricultural ecosystems: A meta-analysis. Sci. Total Environ. 2019, 654, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Clough, T.J.; Condron, L.M.; Kammann, C.; Müller, C. A review of biochar and soil nitrogen dynamics. Agronomy 2013, 3, 275–293. [Google Scholar] [CrossRef]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to Salt Stress in Lettuce: Changes in Chlorophyll Fluorescence Parameters, Phytochemical Contents, and Antioxidant Activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Van Hoorn, J.W.; Katerji, N.; Hamdy, A.; Mastrorilli, M. Effect of Salinity on Yield and Nitrogen Uptake of Four Grain Legumes and on Biological Nitrogen Contribution from the Soil. Agric. Water Manag. 2001, 51, 87–98. [Google Scholar] [CrossRef]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global Plant-responding Mechanisms to Salt Stress: Physiological and Molecular Levels and Implications in Biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef]
- Geilfus, C.M. Review on the significance of chlorine for crop yield and quality. Plant Sci. 2018, 270, 114–122. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Wang, X.F. Experimental Guidance of Plant Physiology; South China University of Technology Press: Guangzhou, China, 2022; pp. 35–36. [Google Scholar]
- Hasnain, M.; Munir, N.; Abideen, Z.; Zulfiqar, F.; Koyro, H.W.; El-Naggar, A.; Caçador, I.; Duarte, B.; Rinklebe, J.; Yong, J.W.H. Biochar-Plant Interaction and Detoxification Strategies under Abiotic Stresses for Achieving Agricultural Resilience: A Critical Review. Ecotoxicol. Environ. Saf. 2023, 249, 114408. [Google Scholar] [CrossRef] [PubMed]
- Abbas, G.; Chen, Y.; Khan, F.Y.; Feng, Y.; Palta, J.A.; Siddique, K.H.M. Salinity and Low Phosphorus Differentially Affect Shoot and Root Traits in Two Wheat Cultivars with Contrasting Tolerance to Salt. Agronomy 2018, 8, 155. [Google Scholar] [CrossRef]
- Giraud, E.; Ho, L.H.; Clifton, R.; Carroll, A.; Estavillo, G.; Tan, Y.F.; Howell, K.A.; Ivanova, A.; Pogson, B.J.; Millar, H.; et al. The absence of ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress. Plant Physiol. 2008, 147, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Iqbal, N.; Syeed, S.; Khan, N.A. Salicylic Acid Alleviates Decreases in Photosynthesis under Salt Stress by Enhancing Nitrogen and Sulfur Assimilation and Antioxidant Metabolism Differentially in Two Mungbean Cultivars. J. Plant Physiol. 2011, 168, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Ashfaque, F. Exogenously Applied H2O2 Promotes Proline Accumulation, Water Relations, Photosynthetic Efficiency and Growth of Wheat (Triticum aestivum L.) Under Salt Stress. Annu. Res. Rev. Biol. 2014, 4, 105–120. [Google Scholar] [CrossRef]
- Rennenberg, H. The fate of excess sulfur in higher plants. Annu. Rev. Plant Biol. 1984, 35, 121–153. [Google Scholar] [CrossRef]
- Banci, L. Structural properties of peroxidases. J. Biotechnol. 1997, 53, 253–263. [Google Scholar] [CrossRef]
- Alamri, S.; Siddiqui, M.H.; Mukherjee, S.; Kumar, R.; Kalaji, H.M.; Irfan, M.; Minkina, T.; Rajput, V.D. Molybdenum-Induced Endogenous Nitric Oxide (NO) Signaling Coordinately Enhances Resilience through Chlorophyll Metabolism, Osmolyte Accumulation and Antioxidant System in Arsenate Stressed-Wheat (Triticum aestivum L.) Seedlings. Environ. Pollut. 2022, 292, 118268. [Google Scholar] [CrossRef]
- Liu, P.; Yang, Y.S.; Xu, G.D.; Fang, Y.H.; Yang, Y.A. The response of antioxidant enzymes of three soybean varieties to molybdenum and boron in soil with a connection to plant quality. Plant Soil Environ. 2005, 51, 351. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, C.; Zhao, X.; Tan, Q.; Sun, X.; Cao, A.; Cui, M.; Zhang, Y. Molybdenum improves antioxidant and osmotic-adjustment ability against salt stress in Chinese cabbage (Brassica campestris L. ssp. Pekinensis). Plant soil. 2012, 355, 375–383. [Google Scholar] [CrossRef]
- Al-Issawi, M.; Rihan, H.Z.; Al-Shmgani, H.; Fuller, M.P. Molybdenum application enhances antioxidant enzyme activity and COR15a protein expression under cold stress in wheat. J. Plant Interact. 2016, 11, 5–10. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Xu, S.; Sun, X. Nitric Oxide Mediates Molybdenum-Induced Antioxidant Defense in Wheat under Drought Stress. Front. Plant Sci. 2017, 8, 1085. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Sairam, R.K.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ Response Mechanisms to Salinity Stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef]
- Hassan, N.A.; Drew, J.V.; Knudsen, D.; Olson, R.A. Influence of soil salinity on production of dry matter and uptake and distribution of nutrients in barley and corn: I. Barley (Hordeum vulgare L.). Agron. J. 1970, 62, 43–45. [Google Scholar] [CrossRef]
- Dahiya, S.S.; Singh, M. Effect of salinity, alkalinity and iron application on the availability of iron, manganese, phosphorus and sodium in pea (Pisum sativum L.) crop. Plant Soil. 1976, 44, 697–702. [Google Scholar] [CrossRef]
- Okçu, G.; Kaya, M.D.; Atak, M. Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. 2005, 29, 237–242. [Google Scholar]
- Yousfi, S.; Mahmoudi, H.; Abdelly, C.; Gharsalli, M. Effect of salt on physiological responses of barley to iron deficiency. Plant Physiol. Biochem. 2007, 45, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Sarani, S. Growth, biochemical components and ion content of Chamomile (Matricaria chamomilla L.) under salinity stress and iron deficiency. J. Saudi Soc. Agric. Sci. 2012, 11, 37–42. [Google Scholar] [CrossRef]
- Rabhi, M.; Barhoumi, Z.; Ksouri, R.; Abdelly, C.; Gharsalli, M. Interactive Effects of Salinity and Iron Deficiency in Medicago Ciliaris. Comptes Rendus Biol. 2007, 330, 779–788. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Average | SD | p-Value |
|---|---|---|---|
| Carbonization yield | 28% | 3.8% | >0.05 |
| Bulk density (kg m−3) | 138.2 | 11.8 | >0.05 |
| HHV (MJ kg−1) | 32.6 | 1.9 | >0.05 |
| Percentage of calorific value from pyrolysis | 80% | 12% | >0.05 |
| H | 1.29% | 0.52% | >0.05 |
| C | 84.20% | 11.18% | >0.05 |
| N | 0.84% | 0.11% | >0.05 |
| O | 1.33% | 0.69% | >0.05 |
| Carbonate as CO2 | 2.85% | 0.54% | >0.05 |
| Carbonate (organic) * | 74.1% | 11.2% | >0.05 |
| Sulfur (total) | 0.03% | 0.01% | >0.05 |
| H/C | 0.21 | 0.08 | >0.05 |
| O/C | 0.014 | 0.002 | >0.05 |
| C/N | 201.5 | 81.2 | >0.05 |
| Ash | 4.50% | 1.21% | >0.05 |
| pH | 8.79 | 1.1 | >0.05 |
| Specific Surface Area (m2 g−1) | 274 | 10.4 | >0.05 |
| Volatile matter | 4.1% | 0.14% | >0.05 |
| Electrical conductivity (µS cm−1) | 341 | 18.2 | >0.05 |
| Salt content (g kg−1) | 0.412 | 0.05 | >0.05 |
| Concentration (mg kg−1) | Average | SD | p-Value |
|---|---|---|---|
| P | 590 | 22 | >0.05 |
| Mg | 1300 | 95 | >0.05 |
| Ca | 12,500 | 250 | >0.05 |
| K | 5500 | 121 | >0.05 |
| Na | 320 | 10 | >0.05 |
| Fe | 1550 | 120 | >0.05 |
| Si | 9900 | 99 | >0.05 |
| S | 290 | 13 | >0.05 |
| Pb | 30 | 5 | >0.05 |
| Cd | 0.15 | 0.02 | >0.05 |
| Cu | 30 | 2.1 | >0.05 |
| Ni | 7 | 1.2 | >0.05 |
| Hg | 0.02 | 0.01 | >0.05 |
| Zn | 90 | 9.2 | >0.05 |
| Cr | 21 | 1.6 | >0.05 |
| B | 26 | 2.7 | >0.05 |
| Mn | 350 | 62 | >0.05 |
| Benzo(k)Fluoranthene | <0.1 | - | - |
| 7,12-Dimethylbenz(a)anthracene | <0.1 | - | - |
| Fluoranthene | <0.1 | - | - |
| Naphthalene | 0.4 | 0.01 | >0.05 |
| Phenanthrene | 0.2 | 0.02 | >0.05 |
| SUM PAHs (EPA) | 0.92 | 0.3 | >0.05 |
| MDA | GPX | |
|---|---|---|
| Mo | R2 0.764 (−1.117) | - |
| Thiols | - | R2 0.872 (−63.952) |
| Na | Cl | |
|---|---|---|
| Fe | R2 0.904 (−95.185) | R2 0.966 (−0.042) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppa, E.; Quagliata, G.; Venanzi, R.; Bruschini, A.; Bianchini, L.; Picchio, R.; Astolfi, S. Potential Use of Biochar as a Mitigation Strategy for Salinity-Related Issues in Tomato Plants (Solanum lycopersicum L.). Environments 2024, 11, 17. https://doi.org/10.3390/environments11010017
Coppa E, Quagliata G, Venanzi R, Bruschini A, Bianchini L, Picchio R, Astolfi S. Potential Use of Biochar as a Mitigation Strategy for Salinity-Related Issues in Tomato Plants (Solanum lycopersicum L.). Environments. 2024; 11(1):17. https://doi.org/10.3390/environments11010017
Chicago/Turabian StyleCoppa, Eleonora, Giulia Quagliata, Rachele Venanzi, Alessandro Bruschini, Leonardo Bianchini, Rodolfo Picchio, and Stefania Astolfi. 2024. "Potential Use of Biochar as a Mitigation Strategy for Salinity-Related Issues in Tomato Plants (Solanum lycopersicum L.)" Environments 11, no. 1: 17. https://doi.org/10.3390/environments11010017
APA StyleCoppa, E., Quagliata, G., Venanzi, R., Bruschini, A., Bianchini, L., Picchio, R., & Astolfi, S. (2024). Potential Use of Biochar as a Mitigation Strategy for Salinity-Related Issues in Tomato Plants (Solanum lycopersicum L.). Environments, 11(1), 17. https://doi.org/10.3390/environments11010017