
Exposure of Zebrafish Embryos to Urea Affects NOS1 Gene Expression in Neuronal Cells

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Urea Treatment
2.3. RNA Purification and Reverse Transcription
2.4. Quantitative Real-Time PCR
2.5. Synthesis of Riboprobes for NOS1, NOS2a, and NOS2b
2.6. In Situ Hybridization
2.7. Immunofluorescence Staining for Anti-GFP
2.8. In Situ Hybridization Combined with TUNEL Assay on Paraffin Section
2.9. Diaminofluorescein-FM Diacetate (DAF-FM DA)
2.10. Statistical Analysis
3. Results
3.1. Quantitative Analysis of NOS Genes during Brain Development in Zebrafish Embryos
3.2. Spatiotemporal Expression Pattern of nos1, nos2a, and nos2b Genes during Embryonic Development in Zebrafish
3.3. Nos1 Gene Is Specifically Expressed in Neuronal Cells during Zebrafish Embryonic Development
3.4. Urea Toxicity Monitoring in Zebrafish Embryos
3.5. Urea Exposure Affects Neuronal nos1 Expression during Development in Zebrafish Embryos
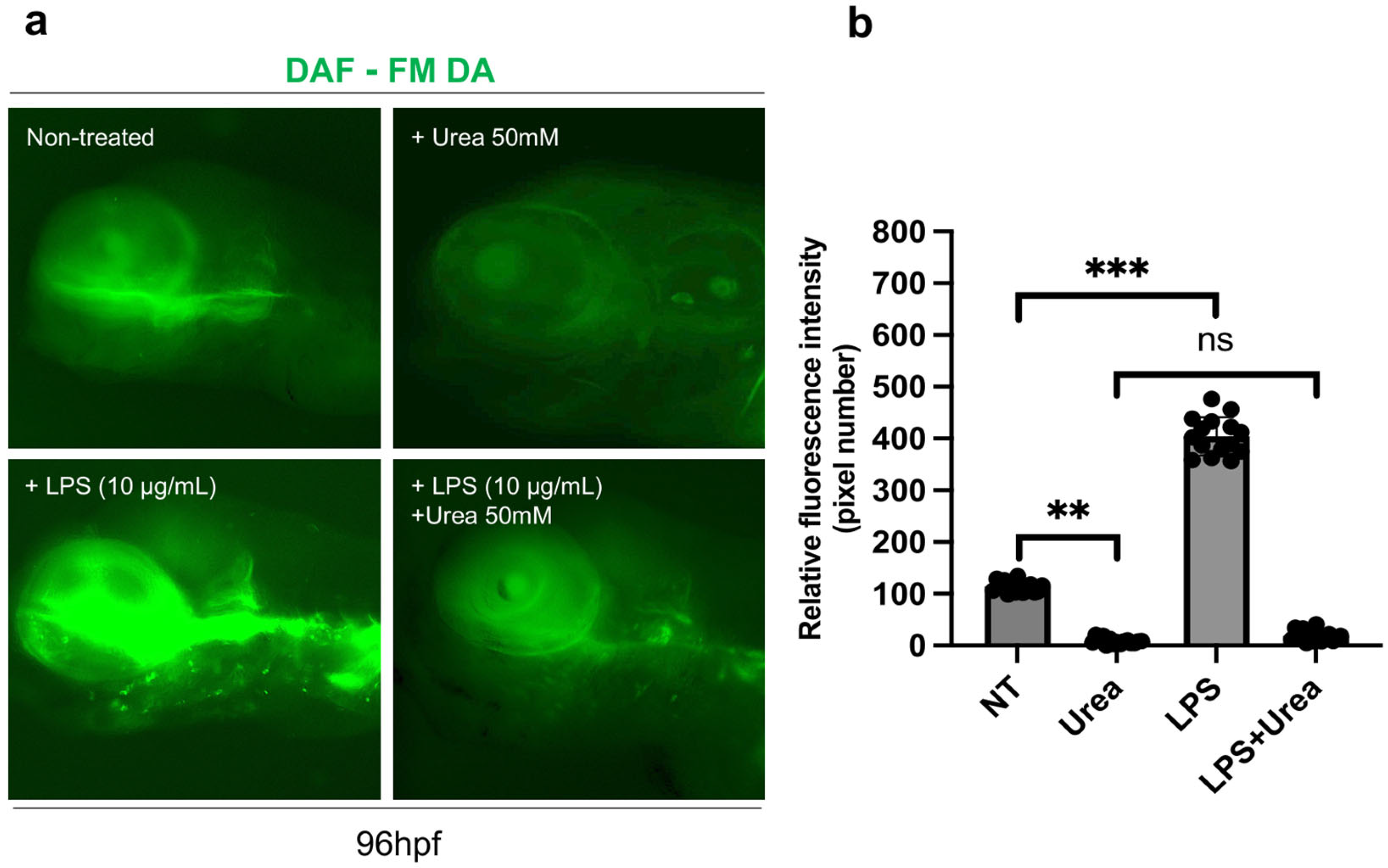
3.6. Effects of Urea Treatment on Proliferation, Cell Death and NO Production in Zebrafish Embryos
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, D.; Liu, Y.; Yang, G.; Zhang, A. Water- and Fertilizer-Integrated Hydrogel Derived from the Polymerization of Acrylic Acid and Urea as a Slow-Release N Fertilizer and Water Retention in Agriculture. J. Agric. Food Chem. 2018, 66, 5762–5769. [Google Scholar] [CrossRef]
- Ruthrof, K.X.; Steel, E.; Misra, S.; McComb, J.; O’Hara, G.; Hardy, G.; Howieson, J. Transitioning from phosphate mining to agriculture: Responses to urea and slow release fertilizers for Sorghum bicolor. Sci. Total Environ. 2018, 625, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Saggar, S.; Singh, J.; Giltrap, D.L.; Zaman, M.; Luo, J.; Rollo, M.; Kim, D.G.; Rys, G.; van der Weerden, T.J. Quantification of reductions in ammonia emissions from fertiliser urea and animal urine in grazed pastures with urease inhibitors for agriculture inventory: New Zealand as a case study. Sci. Total Environ. 2013, 465, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Witte, C.P. Urea metabolism in plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Tasistro, A.S.; Cabrera, M.L.; Ritz, C.W.; Kissel, D.E. Manipulating bedding materials and PLT to reduce NH3 emissions from broiler manure. Bioresour. Technol. 2008, 99, 1952–1960. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Ma, X.; Tang, Q.; Yang, J.; Li, G.; Schuchardt, F. Combined use of nitrification inhibitor and struvite crystallization to reduce the NH3 and N2O emissions during composting. Bioresour. Technol. 2016, 217, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Matczuk, D.; Siczek, A. Effectiveness of the use of urease inhibitors in agriculture: A review. Int. Agrophys. 2021, 35, 197–208. [Google Scholar] [CrossRef]
- Allende-Montalbán, R.; Martín-Lammerding, D.; Delgado, M.D.; Porcel, M.A.; Gabriel, J.L. Urease Inhibitors Effects on the Nitrogen Use Efficiency in a Maize-Wheat Rotation with or without Water Deficit. Agriculture 2021, 11, 684. [Google Scholar] [CrossRef]
- Amorim, M.D.; Toyosumi, I.D.; Lopes, W.D.; Camillo, L.R.; Ferreira, L.N.; Junior, D.S.R.; Soares, W.D.; Gesteira, A.D.; Costa, M.G.C.; Coelho, M.A.N. Partial rootzone drying and regulated deficit irrigation can be used as water-saving strategies without compromising fruit yield and quality in tropically grown sweet orange. J. Hortic. Sci. Biotechnol. 2021, 96, 663–672. [Google Scholar] [CrossRef]
- Silva, F.L.B.; Vieira, L.G.E.; Ribas, A.F.; Moro, A.L.; Neris, D.M.; Pacheco, A.C. Proline accumulation induces the production of total phenolics in transgenic tobacco plants under water deficit without increasing the G6PDH activity. Theor. Exp. Plant Phys. 2018, 30, 251–260. [Google Scholar] [CrossRef]
- Ray, A.; Forrestal, P.; Nkwonta, C.; Rahman, N.; Byrne, P.; Danaher, M.; Richards, K.; Hogan, S.; Cummins, E. Modelling potential human exposure to the urease inhibitor NBPT through the environment-food pathway. Environ. Int. 2023, 179, 108149. [Google Scholar] [CrossRef]
- Ray, A.; Nkwonta, C.; Forrestal, P.; Danaher, M.; Richards, K.; O’Callaghan, T.; Hogan, S.; Cummins, E. Current knowledge on urease and nitrification inhibitors technology and their safety. Rev. Environ. Health 2021, 36, 477–491. [Google Scholar] [CrossRef]
- Dickerson, A.S.; Lee, J.S.; Keshava, C.; Hotchkiss, A.; Persad, A.S. Assessment of Health Effects of Exogenous Urea: Summary and Key Findings. Curr. Environ. Health Rep. 2018, 5, 205–212. [Google Scholar] [CrossRef]
- Zala, N.; Kavishvar, A. Respiratory morbidity due to ammonia exposure among the employees of a urea manufacturing industry located in western part of India. Indian. J. Occup. Environ. Med. 2012, 16, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.H.; Bråtveit, M.; Moen, B.E. Exposure to ammonia and acute respiratory effects in a urea fertilizer factory. Int. J. Occup. Environ. Health 2007, 13, 153–159. [Google Scholar] [CrossRef]
- Quintana, A.; López-Doval, L.; Rodríguez, R.; Souto, J.A.; Casares, J.J. Risk analysis in the chemical industry: An urea-formaldehyde resins case. WIT Trans. Ecol. Environ. 2006, 91, 33. [Google Scholar] [CrossRef]
- Sands, J.M. Regulation of urea transporter proteins in kidney and liver. Mt. Sinai J. Med. 2000, 67, 112–119. [Google Scholar] [PubMed]
- Lu, H.Y.; Ning, X.Y.; Chen, Y.Q.; Han, S.J.; Chi, P.; Zhu, S.N.; Yue, Y. Predictive Value of Serum Creatinine, Blood Urea Nitrogen, Uric Acid, and β2-Microglobulin in the Evaluation of Acute Kidney Injury after Orthotopic Liver Transplantation. Chin. Med. J. 2018, 131, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr.; Moncman, C.L.; Kepka, D.M.; Nebes, V.L.; Diven, W.F.; Dizikes, G.J.; Cederbaum, S.D.; DeFranco, D. Effects of deletions in mouse chromosome 7 on expression of genes encoding the urea-cycle enzymes and phosphoenolpyruvate carboxykinase (GTP) in liver, kidney, and intestine. Biochem. Genet. 1988, 26, 769–781. [Google Scholar] [CrossRef]
- Takatsu, K.; Ishizaka, K. Reaginic antibody formation in the mouse. VI. Suppression of IgE and IgG antibody responses to ovalbumin following the administration of high dose urea-denatured antigen. Cell Immunol. 1975, 20, 276–289. [Google Scholar] [CrossRef]
- Ferrando, S.; Gallus, L.; Gambardella, C.; Marchesotti, E.; Ravera, S.; Franceschini, V.; Masini, M.A. Effects of urea on the molecules involved in the olfactory signal transduction: A preliminary study on Danio rerio. Fish. Physiol. Biochem. 2014, 40, 1793–1800. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Imbrogno, S.; Tota, B.; Gattuso, A. The evolutionary functions of cardiac NOS/NO in vertebrates tracked by fish and amphibian paradigms. Nitric Oxide 2011, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gomez, Y.; Vargas, J.P.; Lopez, J.C.; Portavella, M. Inhibition of brain NOS activity impair spatial learning acquisition in fish. Brain Res. Bull. 2020, 164, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.C.S.; Gayathry, R. Nitric oxide synthase (NOS) inhibitor L-NAME activates inducible NOS/NO system and drives multidimensional regulation of Na(+)/K(+) -ATPase in ionocyte epithelia of immersion-stressed air-breathing fish (Anabas testudineus Bloch). J. Exp. Zool. A Ecol. Integr. Physiol. 2021, 335, 396–416. [Google Scholar] [CrossRef]
- Gray, W.P.; Cheung, A. Nitric Oxide Regulation of Adult Neurogenesis. Nitric Oxide 2014, 96, 59–77. [Google Scholar] [CrossRef]
- Bernstein, H.G.; Keilhoff, G.; Steiner, J.; Dobrowolny, H.; Bogerts, B. Nitric oxide and schizophrenia: Present knowledge and emerging concepts of therapy. CNS Neurol. Disord. Drug Targets 2011, 10, 792–807. [Google Scholar] [CrossRef]
- Hannibal, L. Nitric Oxide Homeostasis in Neurodegenerative Diseases. Curr. Alzheimer Res. 2016, 13, 135–149. [Google Scholar] [CrossRef]
- Kotamraju, S.; Kalivendi, S.; Shang, T.; Kalyanaraman, B. Nitric oxide, proteasomal function, and iron homeostasis—Implications in aging and neurodegenerative diseases. Method. Enzymol. 2005, 396, 526–534. [Google Scholar] [CrossRef]
- Peña-Altamira, E.; Petazzi, P.; Contestabile, A. Nitric Oxide Control of Proliferation in Nerve Cells and in Tumor Cells of Nervous Origin. Curr. Pharm. Des. 2010, 16, 440–450. [Google Scholar] [CrossRef]
- Luo, C.X.; Zhu, D.Y. Research progress on neurobiology of neuronal nitric oxide synthase. Neurosci. Bull. 2011, 27, 23–35. [Google Scholar] [CrossRef]
- Carreira, B.P.; Santos, D.F.; Santos, A.I.; Carvalho, C.M.; Araújo, I.M. Nitric Oxide Regulates Neurogenesis in the Hippocampus following Seizures. Oxidative Med. Cell. Longev. 2015, 2015, 451512. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.X.; Jin, X.; Cao, C.C.; Zhu, M.M.; Wang, B.; Chang, L.; Zhou, Q.G.; Wu, H.Y.; Zhu, D.Y. Bidirectional Regulation of Neurogenesis by Neuronal Nitric Oxide Synthase Derived from Neurons and Neural Stem Cells. Stem Cells 2010, 28, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhu, D.Y. Neuronal nitric oxide synthase: Structure, subcellular localization, regulation, and clinical implications. Nitric Oxide 2009, 20, 223–230. [Google Scholar] [CrossRef]
- Park, H.C.; Kim, C.H.; Bae, Y.K.; Yee, S.Y.; Kim, S.H.; Hong, S.K.; Shin, J.; Yoo, K.W.; Hibi, M.; Hirano, T.; et al. Analysis of upstream elements in the promoter leads to the establishment of transgenic zebrafish with fluorescent neurons. Dev. Biol. 2000, 227, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Mailhe, M.P.; Wagner, I.; Merkler, D.; Golub, R.; Bertrand, J.Y. Synergistic prostaglandin E synthesis by myeloid and endothelial cells promotes fetal hematopoietic stem cell expansion in vertebrates. EMBO J. 2022, 41, e108536. [Google Scholar] [CrossRef] [PubMed]
- Mahony, C.B.; Cacialli, P.; Pasche, C.; Monteiro, R.; Savvides, S.N.; Bertrand, J.Y. Hapln1b, a central organizer of the ECM, modulates kit signaling to control developmental hematopoiesis in zebrafish. Blood Adv. 2021, 5, 4935–4948. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P. Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish. Vet. Sci. 2022, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Dogan, S.; Linnerz, T.; Pasche, C.; Bertrand, J.Y. Minichromosome maintenance protein 10 (mcm10) regulates hematopoietic stem cell emergence in the zebrafish embryo. Stem Cell Reports 2023, 18, 1534–1546. [Google Scholar] [CrossRef]
- Russo, B.; Borowczyk, J.; Cacialli, P.; Moguelet, P.; Truchetet, M.E.; Modarressi, A.; Brembilla, N.C.; Bertrand, J.; Boehncke, W.H.; Chizzolini, C. IL-25 participates in keratinocyte-driven dermal matrix turnover and is reduced in systemic sclerosis epidermis. Rheumatology 2022, 61, 4558–4569. [Google Scholar] [CrossRef]
- Yao, J.; Li, C.; Zhang, J.R.; Liu, S.K.; Feng, J.B.; Wang, R.J.; Li, Y.; Jiang, C.; Song, L.; Chen, A.L.; et al. Expression of nitric oxide synthase (NOS) genes in channel catfish is highly regulated and time dependent after bacterial challenges. Dev. Comp. Immunol. 2014, 45, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ni, L.; Fu, X.; Duan, D.; Xu, J.; Gao, X. A Sulfated Polysaccharide from Saccharina japonica Suppresses LPS-Induced Inflammation Both in a Macrophage Cell Model via Blocking MAPK/NF-kappaB Signal Pathways In Vitro and a Zebrafish Model of Embryos and Larvae In Vivo. Mar. Drugs 2020, 18, 593. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.Y.; Begho, T. Crop residue burning in South Asia: A review of the scale, effect, and solutions with a focus on reducing reactive nitrogen losses. J. Environ. Manag. 2022, 314, 115104. [Google Scholar] [CrossRef]
- Zielonka, M.; Probst, J.; Carl, M.; Hoffmann, G.F.; Kölker, S.; Okun, J.G. Bioenergetic dysfunction in a zebrafish model of acute hyperammonemic decompensation. Exp. Neurol. 2019, 314, 91–99. [Google Scholar] [CrossRef]
- Gil, C.; Hobbs, A.; Schmidlin, A.; Vogel, D.; Wiesinger, H. Generation of nitric oxide from urea cycle intermediates in brain cells. J. Neurochem. 1999, 73, S219. [Google Scholar]
- Gropman, A. Brain imaging in urea cycle disorders. Mol. Genet. Metab. 2010, 100, S20–S30. [Google Scholar] [CrossRef]
- Robinson, C.; Whitehead, M.; Ellenbogen, A.; Ludwig, W.; Gropman, A. Defining Brain Chemical Changes and Areas of the Brain Most Affected by High Ammonia Levels in Children with Urea Cycle Disorders. Ann. Neurol. 2016, 80, S345–S346. [Google Scholar]
- Fu, B.M.M.; Shin, D.W.; Fan, J.; Khadka, N.; Luu, E.; Khalid, W.; Xia, Y.F.; Bikson, M. Inhibition of Nitric Oxide Synthase (NOS) by NG-monomethyl-L-arginine (L-NMMA) Reduces Transient Increase in the Blood-Brain Barrier Solute Permeability in Rat Brain by Transcranial Direct Current Stimulation. FASEB J. 2018, 32, 922.2. [Google Scholar] [CrossRef]
- Chen, L.N.; Sun, J.; Yang, X.D.; Xiao, K.; Lv, Y.; Zhang, B.Y.; Zhou, W.; Chen, C.; Gao, C.; Shi, Q.; et al. The Brain NO Levels and NOS Activities Ascended in the Early and Middle Stages and Descended in the Terminal Stage in Scrapie-Infected Animal Models. Mol. Neurobiol. 2017, 54, 1786–1796. [Google Scholar] [CrossRef]
- Necchi, D.; Virgili, M.; Monti, B.; Contestabile, A.; Scherini, E. Regional alterations of the NO/NOS system in the aging brain: A biochemical, histochemical and immunochemical study in the rat. Brain Res. 2002, 933, 31–41. [Google Scholar] [CrossRef]
- da Silva Chaves, S.N.; Dutra Costa, B.P.; Vidal Gomes, G.C.; Lima-Maximino, M.; Pacheco Rico, E.; Maximino, C. NOS-2 participates in the behavioral effects of ethanol withdrawal in zebrafish. Neurosci. Lett. 2020, 728, 134952. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Fang, P.; Lin, J.F.; Pan, H.C.; Zhang, F.; Shen, Y.Q. Expression of gdnf and nos in adult zebrafish brain during the regeneration after spinal cord injury. Yi Chuan 2013, 35, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Annona, G.; Ferran, J.L.; De Luca, P.; Conte, I.; Postlethwait, J.H.; D’Aniello, S. Expression Pattern of in the Developing Nervous System of Ray-Finned Fish. Genes 2022, 13, 918. [Google Scholar] [CrossRef]
- Gutiérrez, H.C.; O’Leary, A.; Freudenberg, F.; Fedele, G.; Wilkinson, R.; Markham, E.; van Eeden, F.; Reif, A.; Norton, W.H.J. Nitric oxide interacts with monoamine oxidase to modulate aggression and anxiety-like behaviour. Eur. Neuropsychopharm. 2020, 30, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Husson, A.; Brasse-Lagnel, C.; Fairand, A.; Renouf, S.; Lavoinne, A. Argininosuccinate synthetase from the urea cycle to the citrulline-NO cycle. Eur. J. Biochem. 2003, 270, 1887–1899. [Google Scholar] [CrossRef] [PubMed]
- Javrushyan, H.; Avtandilyan, N.; Trchounian, A. The effects of NO on the urea cycle pathway in short-term intermittent hypobaric hypoxia in rats. Respir. Physiol. Neurobiol. 2021, 285, 103598. [Google Scholar] [CrossRef]
- Shekar Roy, H.; Neethu, K.M.; Rajput, S.; Sadhukhan, S.; Gowri, V.; Hassan Dar, A.; Monga, M.; Salaria, N.; Guha, R.; Chattopadhyay, N.; et al. Efficient Nitric Oxide Scavenging by Urea-Functionalized Push-Pull Chromophore Modulates NO-Mediated Diseases. Chemistry 2023, 29, e202301748. [Google Scholar] [CrossRef]
- Moeslinger, T.; Spieckermann, P.G. Urea-induced inducible nitric oxide synthase inhibition and macrophage proliferation. Kidney Int. Suppl. 2001, 78, S2–S8. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ef1a-QF | CTACCTACCCTCCTCTTGGT |
| ef1a-QR | CTTCTGTGCAGACTTTGTGA |
| nos1-QF | TTGGATAGTGCCACCCATGT |
| nos1-QR | GCGTCCCATTCACTCCTTTC |
| nos2a-QF | TGGAGAAATGCGCCAAGATG |
| noa2a-QR | CCGTTGGTGGCATACTTCAG |
| nos2b-QF | TGGCTCTCGGATGTATCCAC |
| nos2b-QR | CAGCTCATCTCCCTCTCCTG |
| pcna-QF | CAAGGAGGATGAAGCGGTAACA |
| pcna-QR | CTGCGGACATGCTAAGTGTG |
| nos1-F | CGCTCACAAACCAGATGAAG |
| nos1-R | GCTGCATACAGATCTCTGTAAG |
| nos2a-F | CCAGGACTCACTTCATCACA |
| nos2a-R | CCATCTGATAGCCAGCATACT |
| nos2b-F | CGTGCCATCAAGATGTGTTAC |
| nos2b-R | TAGCCAGCGTATCGTATGAG |
| pcna-F | CCTTAAGAAGGTCCTGGAG |
| pcna-R | CCACACAACTGTATTCCTGCTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacialli, P.; Ricci, S.; Frabetti, F.; Ferrando, S.; Franceschini, V. Exposure of Zebrafish Embryos to Urea Affects NOS1 Gene Expression in Neuronal Cells. Environments 2024, 11, 41. https://doi.org/10.3390/environments11030041
Cacialli P, Ricci S, Frabetti F, Ferrando S, Franceschini V. Exposure of Zebrafish Embryos to Urea Affects NOS1 Gene Expression in Neuronal Cells. Environments. 2024; 11(3):41. https://doi.org/10.3390/environments11030041
Chicago/Turabian StyleCacialli, Pietro, Serena Ricci, Flavia Frabetti, Sara Ferrando, and Valeria Franceschini. 2024. "Exposure of Zebrafish Embryos to Urea Affects NOS1 Gene Expression in Neuronal Cells" Environments 11, no. 3: 41. https://doi.org/10.3390/environments11030041
APA StyleCacialli, P., Ricci, S., Frabetti, F., Ferrando, S., & Franceschini, V. (2024). Exposure of Zebrafish Embryos to Urea Affects NOS1 Gene Expression in Neuronal Cells. Environments, 11(3), 41. https://doi.org/10.3390/environments11030041