
Assessing the Climatic Effects on Vegetation Dynamics in the Mekong River Basin


Abstract
:1. Introduction
2. Materials and Methods
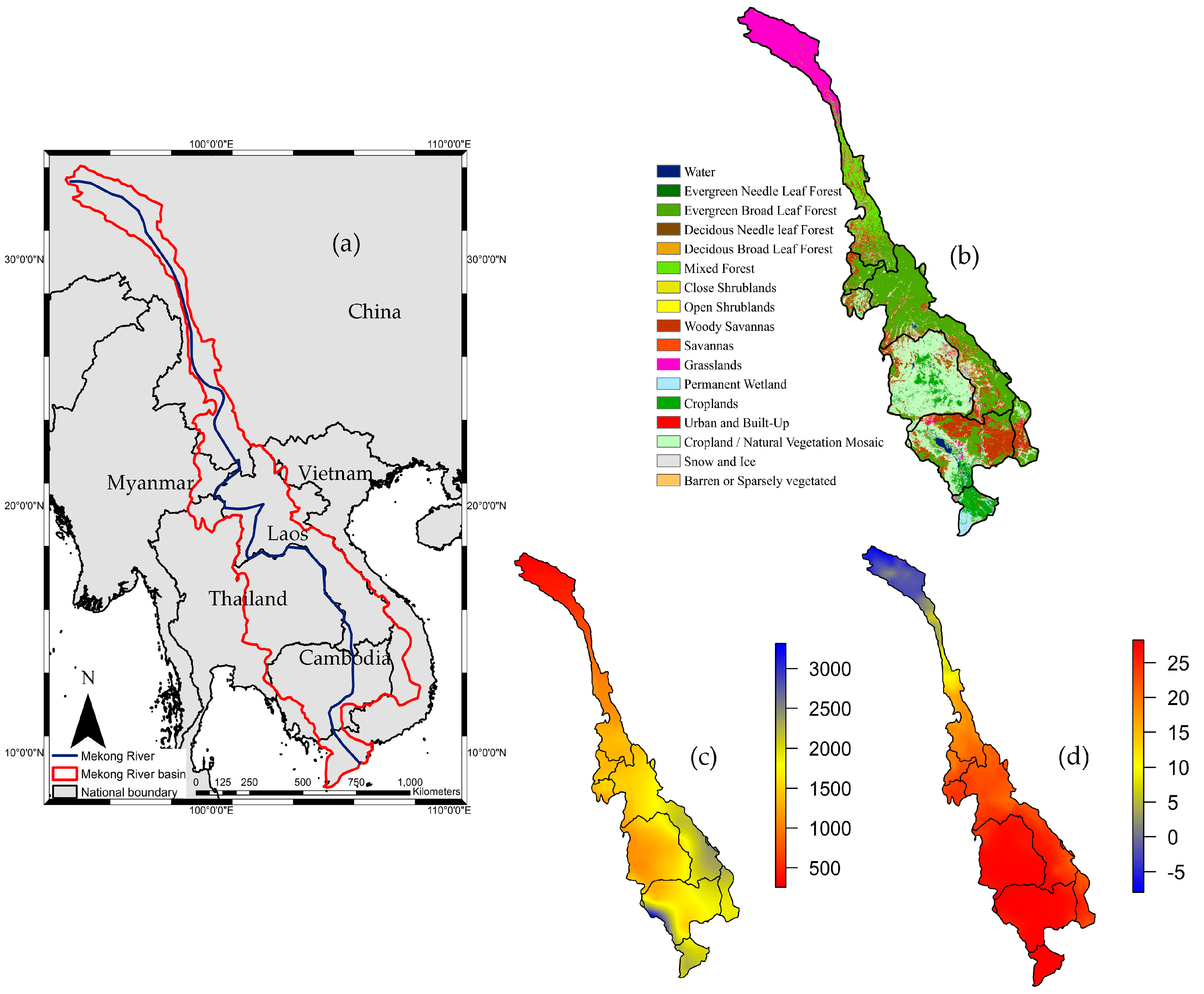
2.1. Study Area Description
2.2. Data Source and Pre-Processing
2.2.1. Land Cover Map
2.2.2. NDVI Data Set
2.2.3. Climate Data
2.3. Methods
2.3.1. Coefficient of Variation
2.3.2. Sen’s Slope Analysis
2.3.3. Mann–Kendall Trend Test
2.3.4. Correlation Analysis
2.3.5. Partial Least Square Regression (PLSR) Analysis
4. Results and Discussion
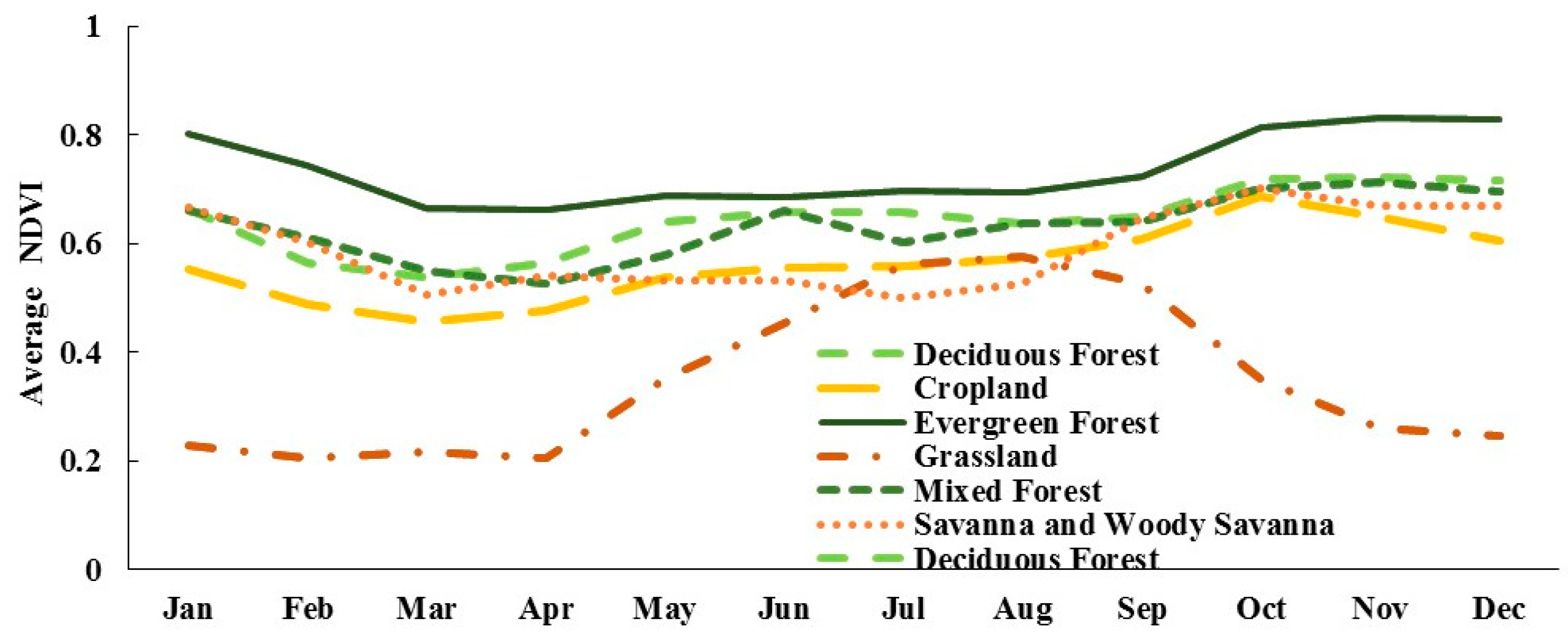
4.1. Seasonal Vegetation Dynamics
4.2. Interannual Changes in NDVI and Climate Variables for Different Vegetation Types
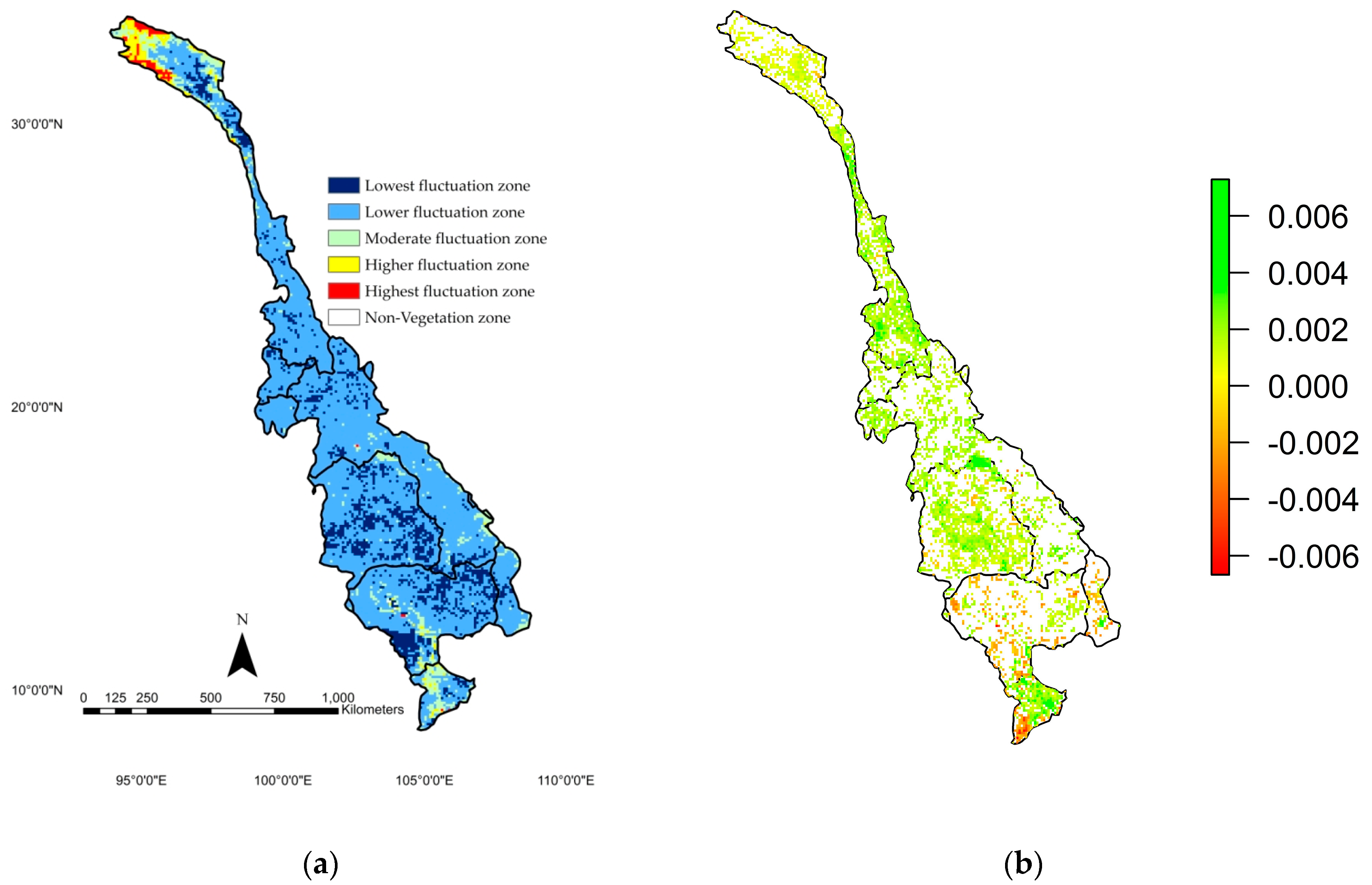
4.3. Spatial Annual NDVI Fluctuations and Trend Analyses
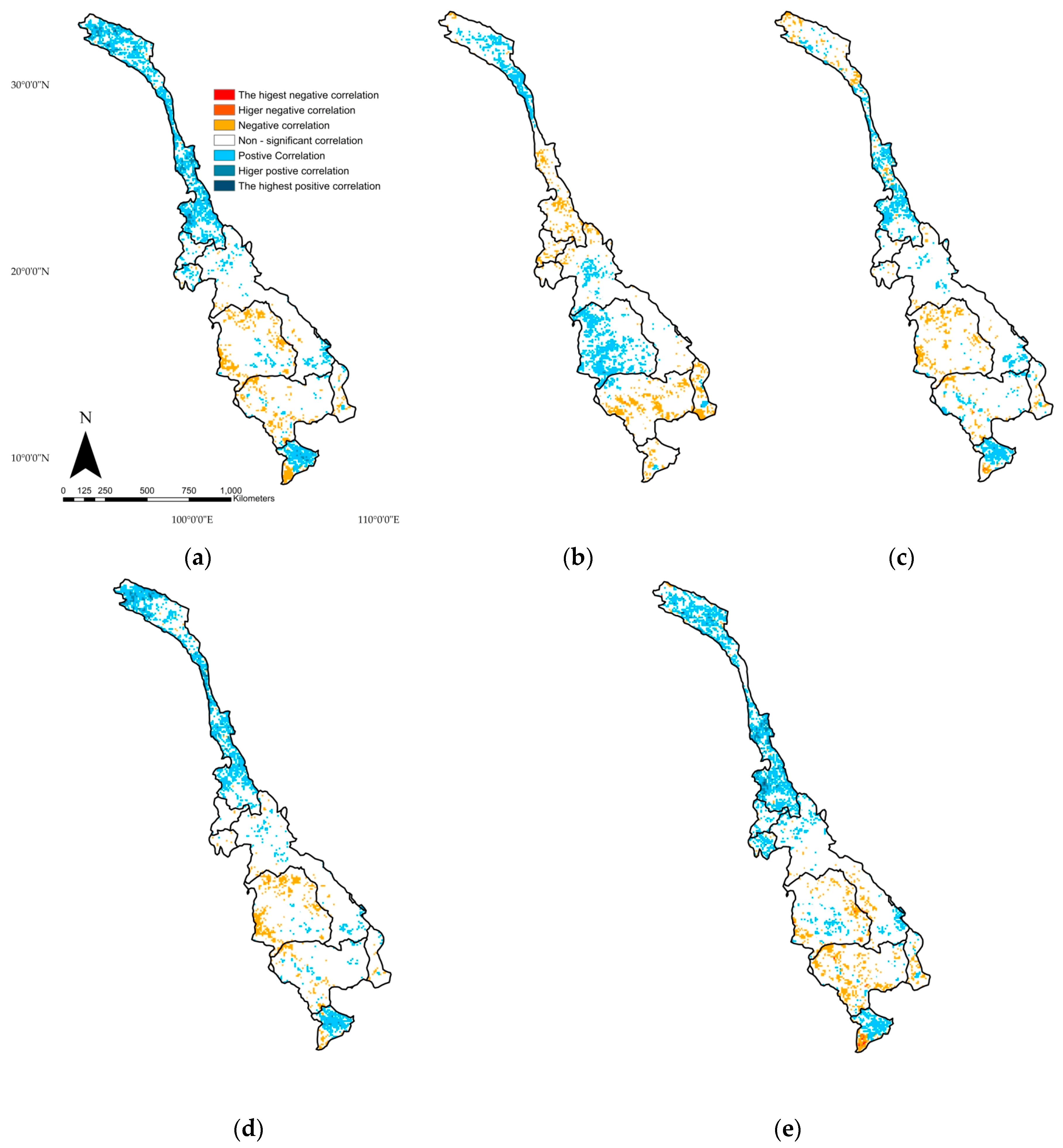
4.4. Correlation Analysis between NDVI and Climate Variables
4.5. Climate Driving Factors of Vegetation Dynamics
5. Conclusions
- (1)
- The patterns of seasonal vegetation dynamics vary depending on vegetation types. The highest seasonality was shown by grassland, which is logic being annual plants, while evergreen forest was the one showing less seasonality. The average trends of NDVI seasonal dynamics were similar in the majority of vegetation types, with the main exception being grasslands, which displayed a significantly different pattern. However, there needs to be more focus on elevation and other topographic factors in future research.
- (2)
- NDVI, air temperature, precipitation, and PET were shown to have increasing temporal trends at all locations. Moreover, the spatial trend of NDVI confirmed that NDVI at the basin scale was found to be mostly increasing relative to the lowest and lower fluctuation levels during the last three decades.
- (3)
- Climate variables, air temperature, precipitation, and PET were shown to have positive correlations with vegetation in the upper part of the basin. In the lower part of the basin, evergreen forest demonstrated non-significant relationships with these variables. Cropland was only found to have a positive correlation with precipitation, while the savanna and woody savanna ecosystems were shown to have significant negative relationships with precipitation.
- (4)
- The climatic factors driving NDVI were dependent on vegetation types. Air temperature and PET governed the greenness of cropland, while the different types of forest show different controlling climatic factors; the evergreen forest biome is controlled by air temperature, while air temperature and precipitation are crucial for deciduous forests. The average rise in temperature drives greenness in a mixed forest. PET was important for the grassland biome in order to promote greenness, while precipitation could reduce its greenness. For savanna and woody savanna, precipitation was the sole negative contributor.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Overpeck, J.T.; Rind, D.; Goldberg, R. Climate-induced changes in forest disturbance and vegetation. Nature 1990, 343, 51–53. [Google Scholar] [CrossRef]
- Prasad, V.K.; Basarinath, K.V.S.; Eaturu, A. Effects of precipitation, temperature and topographic parameters on evergreen vegetation greenery in the Western Ghats, India. Int. J. Clomatol. 2008, 28, 1807–1819. [Google Scholar] [CrossRef]
- Yu, H.; Xu, J.; Okuto, E.; Luedeling, E. Seasonal response of grassland to climate change on the Tibetan Plateau. PLoS ONE. 2012, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chuai, X.W.; Huang, X.J.; Wang, W.J.; Bao, G. NDVI, temperature and precipitation changes and their relationships with different vegetation types during 1998–2007 in Inner Mongolia, China. Int. J. Clomatol. 2013, 33, 1696–1706. [Google Scholar] [CrossRef]
- Thavorntam, W.; Tantemsapya, N. Vegetation greenness modeling in response to climate change for Northeast Thailand. J. Geogr. 2012, 23, 1052–1068. [Google Scholar] [CrossRef]
- Cao, R.; Jiang, W.; Yuan, L.; Wang, W.; Lv, A.; Chen, A. Inter-annual variations in vegetation and their response to climatic factors in the upper catchments of the Yellow River from 2000 to 2010. J. Geogr. 2014, 24, 963–979. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, X.; Tao, J.; Wu, J.; Wang, J.; Shi, P.; Zhang, Y.; Yu, C. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agr. Forest. Meteorol. 2014, 189–190, 11–18. [Google Scholar] [CrossRef]
- Wang, H.; Chen, A.; Wang, Q.; He, B. Drought dynamics and impacts on vegetation in China from 1982 to 2011. Ecol. Eng. 2015, 75, 303–307. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends. Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.J.; Justice, C.O.; Prince, S.D. Monitoring the grasslands of the Sahel 1984–1985. Int. J. Remote Sens. 1986, 7, 1571–1581. [Google Scholar] [CrossRef]
- Chamaille-Jammes, S.; Fritz, H.; Murindagomo, F. Spatial patterns of the NDVI–rainfall relationship at the seasonal and interannual time scales in an African savanna. Int. J. Remote Sens. 2006, 27, 5185–5200. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X. Detecting Climate Effects on Vegetation in Northern Mixed Prairie Using NOAA AVHRR 1-km Time-Series NDVI Data. Remote Sens. 2012, 4, 120–134. [Google Scholar] [CrossRef]
- Potter, C. Vegetation cover change in the Upper Kong River basin of the Sierra Nevada detected using Landsat satellite image analysis. Climatic Change. 2015, 131, 635–647. [Google Scholar] [CrossRef]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Tucker, C.J.; Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Myneni, R.B.; Taylor, M.G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int. J. Biometeorol. 2001, 45, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, J.; Zhou, L.; Tucker, C.J.; Myneni, R.B.; Ceulemans, R. Evidence fora persistent and extensive greening trend in Eurasia inferred from satellite vegetation index data. J. Geophys. Res.: Atmos. 2002, 107, 1–14. [Google Scholar] [CrossRef]
- Huang, C.; Li, Y.; Liu, G.; Zhang, H.; Liu, Q. Recent climate variability and its impact on precipitation, temperature, and vegetation dynamics in the Lancang River headwater area of China. Int. J. Remote Sens. 2014, 38, 2822–2834. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Guo, H.; Leinenkugel, P.; Zhou, Y.; Li, L.; Shen, Q. Drought impact on vegetation productivity in the Lower Mekong Basin. Int. J. Remote Sens. 2014, 35, 2835–2856. [Google Scholar] [CrossRef]
- Guo, L.; Wu, S.; Zhao, D.; Yin, Y.; Leng, G.; Zhang, Q. NDVI-based vegetation change in Inner Mongolia from 1982 to 2006 and its relationship to climate at biome scale. Adv. Meteoro. 2014. [Google Scholar] [CrossRef]
- Mekong River Commission (MRC). Overview of the Hydrology of the Mekong Basin; Mekong River Commission: Vientiane, Laos, 2005; pp. 12–14. [Google Scholar]
- Kite, G. Modelling the Mekong: Hydrological simulation for environmental impact studies. J. Hydrol. 2001, 335, 1–13. [Google Scholar] [CrossRef]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS collection 5 global land cover: Algorithm refinements and characterization of new dataset. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple methods for reconstructing a high-quality NDVI time series data set based on the Savitzky-Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Geographical Data Analysis and Modeling, R Packages Version 2.5-8. Available online: https://cran.r-project.org/web/packages/raster/ (accessed on 2 July 2016).
- Harris, I.; Jones, T.; Osborn, T.; Lister, D.H. Update high-resolution grids of monthly climatic observation —the CRU TS 3.10 Dataset. Int. J. Climatol. 2013, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Milich, L.; Weiss, E. GAC NDVI interannual coefficient of variation (CoV) images: Ground truth sampling of the Sahel along north-south transects. Int. J. Remote Sens. 2000, 21, 235–260. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Oxford University Press: London, UK, 1975. [Google Scholar]
- Sen, P.K. Estimate of the regression coefficient based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 379–1389. [Google Scholar] [CrossRef]
- Fensholt, R.; Langanke, T.; Rasmussen, K.; Reenbreg, A.; Prince, S.D.; Tucker, C.; Scholes, R.J.; Le, Q.B.; Bondeau, A.; Eastman, R.; et al. Greenness in semi-arid areas across the globe 1981–2007—An Earth Observing Satellite based analysis of trends and drivers. Remote Sens. Environ. 2012, 121, 144–158. [Google Scholar] [CrossRef]
- Wold, H. The Fixed-Point Approach to Interdependent System; Elsevier Science Ltd.: Amsterdam, The Netherlands, 1981. [Google Scholar]
- Wold, H. Partial least squares. In Encyclopedia of Statistical Sciences; Kotz, S., Johnson, N.L., Eds.; John Wiley and Sons: New York, NY, USA, 1985. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar]
- Garcίa, M.; Litago, J.; Palacios-Orueta, A.; Pinzόn, J.E.; Ustin, S.L. Short-term propagation of rainfall perturbation on terrestrial ecosystems in the central California. Appl. Veg. Sci. 2010, 13, 146–162. [Google Scholar] [CrossRef]
- Van Leeuwen, W.J. D.; Hartfield, K.; Miranda, M.; Meza, F.J. Trends and ENSO/AAO driven variability in NDVI derived productivity and phenology alongside the Andes Mountains. Remote Sens. 2013, 5, 117–1203. [Google Scholar] [CrossRef]
- Rundell, P.W. Vegetation in the Mekong Basin. In The Mekong: Biophysical Environment of an International River Basin; Cambel, I.C., Ed.; Academic Press: Amsterdam, The Netherland, 2009. [Google Scholar]
- Vaiphasa, C.; Piamduaytham, S.; Viphasa, T.; Skidmore, A.K. A Normalized Difference Vegetation index (NDVI) Time-series of idle agriculture lands: A preliminary study. Eng. J. 2011, 15, 9–16. [Google Scholar] [CrossRef]
- Chitpaiboon, C. Farmer’s Livelihood and the Existence of Agriculture Sector in Bang Pla Ma District, Suphan Buri Province. Master’s Thesis, Kasetsart University, Bangkok, Thailand, 31 May 2013. [Google Scholar]
- Zhan, Z.Z.; Liu, B.B.; Li, H.M.; Wu, W.; Zhong, B. The Relationship between NDVI and Terrain factors. Procedia. Environ. Sci. 2012, 12, 765–771. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.T.; Laprise, R.; et al. Regional climate projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, A., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Lacombe, G.; Hoanh, C.T.; Smakhtin, V. Multi-year variability or unidirectional trends? Mapping long-term precipitation and temperature changes in continental Southeast Asia using PRECIS regional climate model. Clim. Chang. 2012, 113, 258–299. [Google Scholar] [CrossRef]
- Future Climate in World Regions: An Intercomparison of Model-Based Projections for the New IPCC Emissions Scenarios. Available online: http://www.ipcc-data.org/documents/scatter_plot_report.pdf (accessed on 15 August 2016).
- Liang, S.; Chen, J.; Jin, X.; Wan, L.; Gong, B. Regularity of vegetation coverage changes in the Tibetan plateau over the last 21 years. Adv. Earth Sci. 2007, 22, 33–38. [Google Scholar]
- Ding, M.; Zhang, Y.; Liu, L.; Zhang, W.; Wang, Z.; Bai, W. The relationship between NDVI and precipitation on the Tibetan Plateau. J. Geogr. 2007, 17, 259–268. [Google Scholar] [CrossRef]
- Cui, X.; Graf, H.F. Recent land cover changes on the Tibetan Plateau: A review. Clim. Chang. 2009, 94, 47–61. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Li, S.; Motesharrei, S. Spatial and Temporal Patterns of Global NDVI Trends: Correlations with Climate and Human Factors. Remote Sens. 2015, 7, 13233–13250. [Google Scholar] [CrossRef]
- Peng, J.; Liu, Z.; Liu, Y.; Wu, J.; Han, Y. Trend analysis of vegetation dynamics in Qinghai-Tibet Plateau using Hurst Exponent. Ecol. Indic. 2012, 14, 28–39. [Google Scholar] [CrossRef]
- Liu, S.; Yan, D.; Shi, X.; Wang, G.; Yuan, Z.; Yin, J. Grassland NDVI response to climate change factors in different vegetation regionalization in China. In Proceedings of International Symposium GRMSE 2013, Wuhan, China, 8–11 November 2013.
- Fang, J.; Piao, S.; Zhou, L.; He, J.; Wei, F.; Myneni, R.B.; Tacker, C.J.; Tan, K. Precipitation patterns alter growth of temperate vegetation. Geophys. Res. Lett. 2005, 32, L21411. [Google Scholar] [CrossRef]
- Tao, F.; Yokosuka, M.; Xu, Y.; Hayashi, Y.; Zhang, Z. Climate changes and trends in phenology and yields of field crops in China, 1981–2000. Agr. Forest Meteorol. 2006, 138, 82–92. [Google Scholar] [CrossRef]
- Lobell, D.B.; Field, C.B. Global scale climate crop-yield relationship and the impact of recent warming. Environ. Res. Lett. 2007, 2, 014002. [Google Scholar] [CrossRef]
- Schlenker, W.; Roberts, M.J. Nonlinear temperature effects indicate severe damages to U.S. crop yields under climate change. PNAS 2009, 106, 15594–15598. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Wang, Z.; Luo, L.; Ren, C. Integrating AVHRR and MODIS data to monitor NDVI changes and their relationship with climatic parameters in Northeast China. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 528–536. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, F.; Guo, M.; Tani, H.; Matsuoka, N.; Matsumura, S. Spatial and Temporal relationship among NDVI, Climate factors and Land cover change in the North Asia from1982 to 2009. GISci. Remote Sens. 2013, 48, 371–393. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types | Climate Zone |
|---|---|
| Cropland | Aw |
| Evergreen forest | Cwa |
| Deciduous forest | Aw |
| Savanna and woody savanna | Aw |
| Mixed forest | Cwb |
| Grassland | ET |
| Vegetation Types | NDVI | T | TMN | TMX | Prec | PET |
|---|---|---|---|---|---|---|
| Cropland | 0.007 * | 0.017 * | 0.020 * | 0.014 | 3.512 | 0.997 |
| Evergreen forest | 0.002 | 0.021 * | 0.026 * | 0.020 * | 8.627 * | 1.272 |
| Deciduous forest | 0.004 | 0.022 * | 0.025 * | 0.014 * | 5.377 | 1.481 * |
| Mixed forest | 0.010 * | 0.037 * | 0.039 * | 0.036 * | −1.576 | 2.861 * |
| Grassland | 0.002 | 0.031 * | 0.033 * | 0.028 * | 0.116 | 1.381 * |
| Savanna and woody savanna | 0.003 | 0.020 * | 0.027 * | 0.015 * | 0.015 * | 1.515 * |
| Vegetation Types | T | TMN | TMX | Prec | PET |
|---|---|---|---|---|---|
| VIP score, PLSR coeff. | VIP score, PLSR coeff. | VIP score, PLSR coeff. | VIP score, PLSR coeff. | VIP score, PLSR coeff. | |
| Cropland | 1.174, 0.790 | 1.140, –0.427 | 1.160, –0.442 | 0.277, 0.0002 | 0.950, –0.0006 |
| Evergreen forest | 1.372, 0.344 | 1.489, 0.534 | 0.872, 0.019 | 0.144, –0.481 | 0.347, –0.464 |
| Deciduous forest | 1.077, –0.130 | 0.935, –0.077 | 1.168, –0.178 | 1.065, 0.348 | 0.685, 0.0009 |
| Mixed forest | 1.095, 0.452 | 1.088, –0.296 | 1.101, –0.129 | 0.672, –0.003 | 0.977, –0.012 |
| Grassland | 0.768, –0.117 | 0.486, –0.021 | 0.833, –0.091 | 0.914, 0.016 | 1.626, –0.056 |
| Savanna and woody savanna | 0.470, 1.271 | 0.656, –0.540 | 0.416, –0.742 | 1.898, –0.0008 | 0.756, –0.0031 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na-U-Dom, T.; Mo, X.; Garcίa, M. Assessing the Climatic Effects on Vegetation Dynamics in the Mekong River Basin. Environments 2017, 4, 17. https://doi.org/10.3390/environments4010017
Na-U-Dom T, Mo X, Garcίa M. Assessing the Climatic Effects on Vegetation Dynamics in the Mekong River Basin. Environments. 2017; 4(1):17. https://doi.org/10.3390/environments4010017
Chicago/Turabian StyleNa-U-Dom, Tawatchai, Xingguo Mo, and Monica Garcίa. 2017. "Assessing the Climatic Effects on Vegetation Dynamics in the Mekong River Basin" Environments 4, no. 1: 17. https://doi.org/10.3390/environments4010017
APA StyleNa-U-Dom, T., Mo, X., & Garcίa, M. (2017). Assessing the Climatic Effects on Vegetation Dynamics in the Mekong River Basin. Environments, 4(1), 17. https://doi.org/10.3390/environments4010017