
Genetic Investigation of Four Beluga Sturgeon (Huso huso, L.) Broodstocks for its Reintroduction in the Po River Basin

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
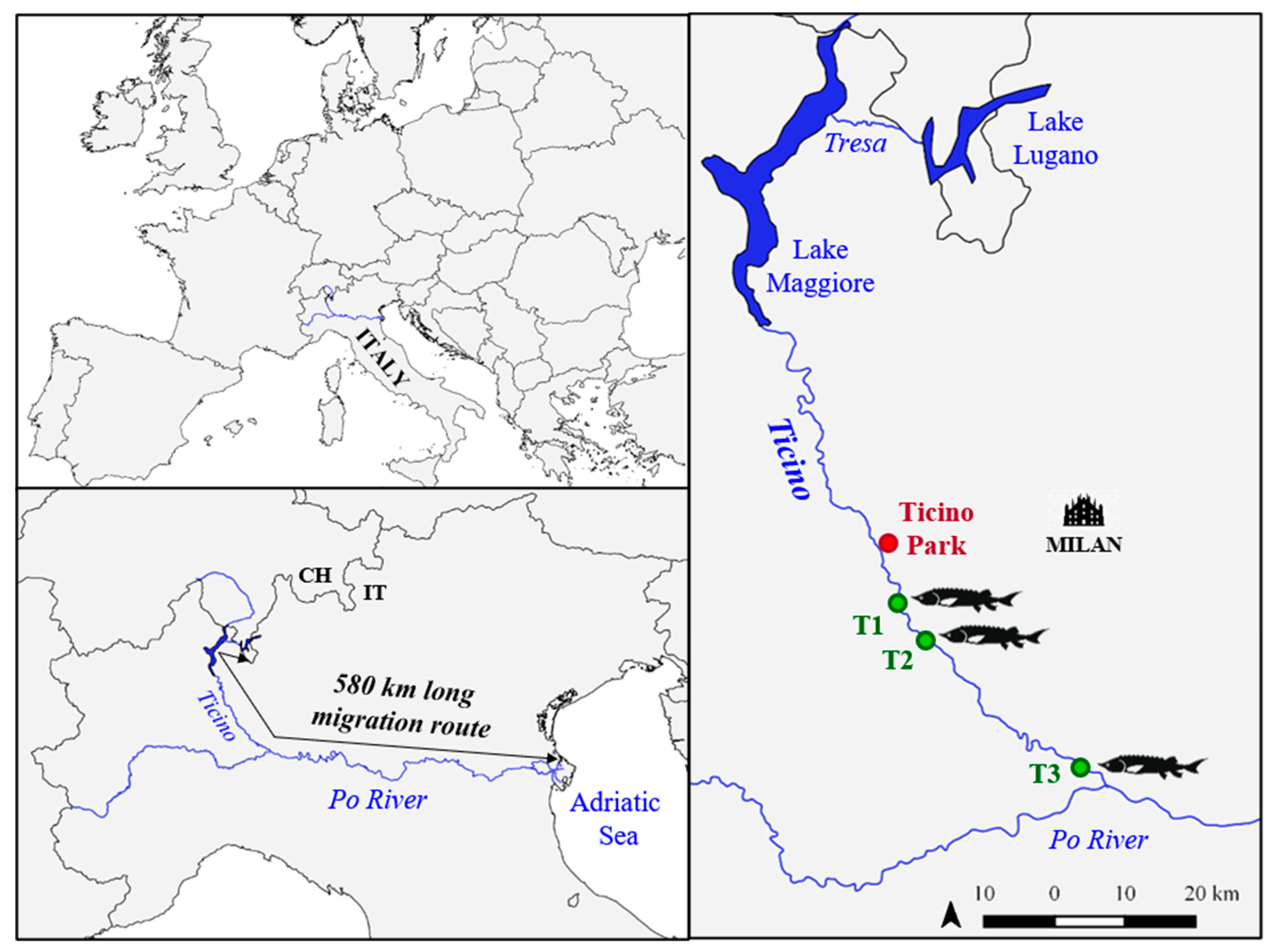
2.1. Broodstocks Description
2.2. Molecular Methods
2.3. Genetic Admixture and Substructure
2.4. Phylogenetic and Phylogeographic Relationship
3. Results
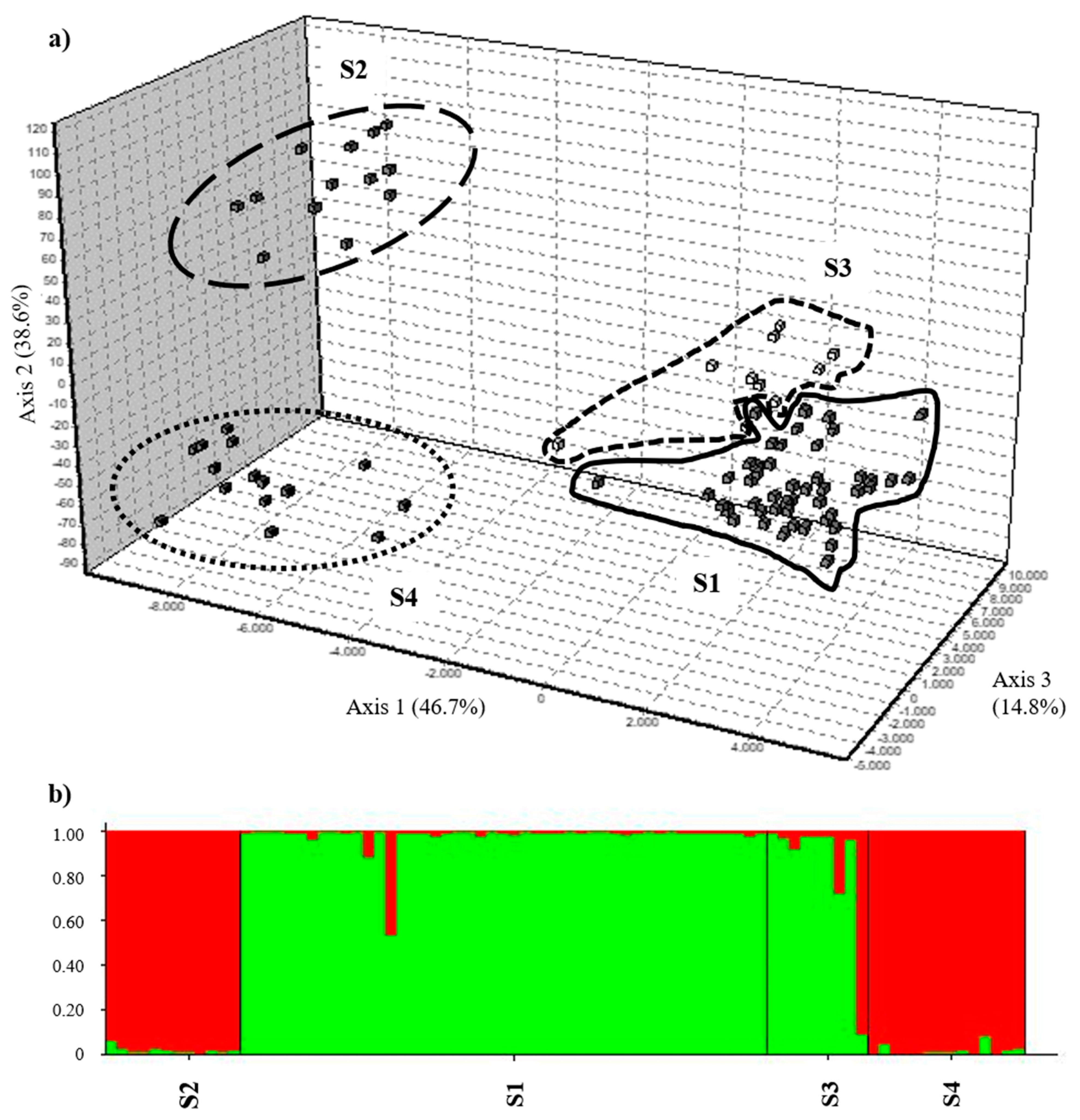
3.1. Genetic Admixture and Substructure
3.2. Mitochondrial Control Region Variability
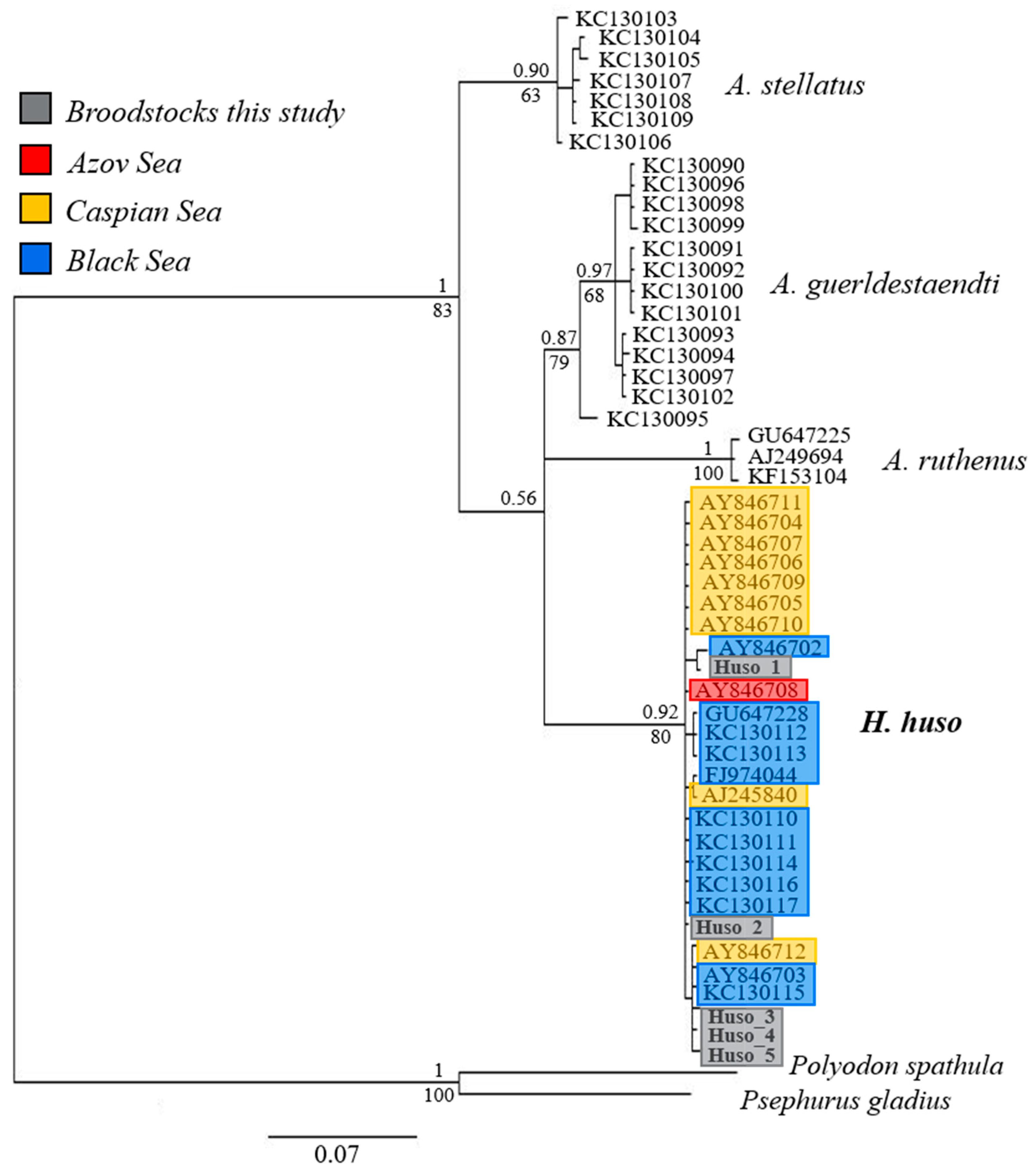
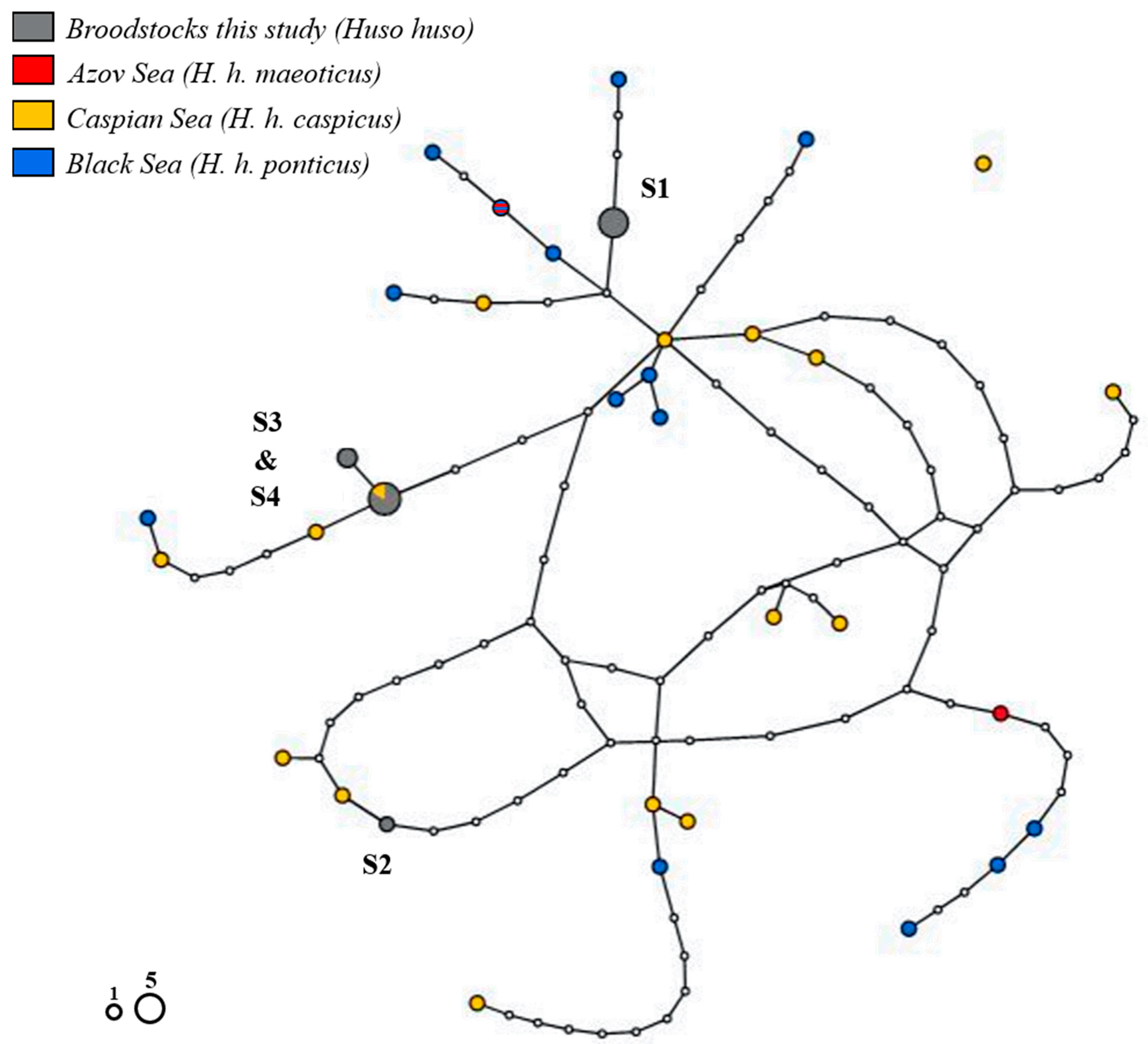
3.3. Phylogenetic and Phylogeographic Relationship
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grill, G.; Lehner, B.; Lumsdon, A.E.; MacDonald, G.K.; Zarfl, C.; Reidy, L.C. An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ. Res. Lett. 2015, 10, 015001. [Google Scholar] [CrossRef]
- Limburg, K.E.; Waldman, J.R. Dramatic Declines in North Atlantic Diadromous Fishes. BioScience 2009, 59, 955–965. [Google Scholar] [CrossRef]
- Branco, P.; Segurado, P.; Santos, J.M.; Ferreira, M.T. Prioritizing barrier removal to improve functional connectivity of rivers. J. Appl. Ecol. 2014, 51, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Segurado, P.; Branco, P.; Avelar, A.P.; Ferreira, M.T. Historical changes in the functional connectivity of rivers based on spatial network analysis and the past occurrences of diadromous species in Portugal. Aquat. Sci. 2015, 77, 427–440. [Google Scholar] [CrossRef]
- Seehausen, O.; Takimoto, G.; Roy, D.; Jokela, J. Speciation reversal and biodiversity dynamics with hybridization in changing environments. Mol. Ecol. 2008, 17, 30–44. [Google Scholar] [CrossRef]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland, 2007; ISBN 9782839902984. [Google Scholar]
- Dudu, A.; Georgescu, S.E.; Costache, M. Molecular analysis of phylogeographic Subspecies in three Ponto-Caspian sturgeon species. Genet. Mol. Biol. 2014, 37, 587–597. [Google Scholar] [CrossRef] [Green Version]
- IUCN 2010, Versions 2010; The IUCN Red List of Threatened Species. 2010.
- Boscari, E.; Marino, I.A.M.; Caruso, C.; Gessner, J.; Lari, M.; Mugue, N.; Barminsteva, A.; Suciu, R.; Onara, D.; Zane, L.; et al. Defining criteria for the reintroduction of locally extinct populations based on contemporary and ancient genetic diversity: The case of the Adriatic Beluga sturgeon (Huso huso). Divers. Distrib. 2021, 00, 1–12. [Google Scholar] [CrossRef]
- Zerunian, S. Piano D’azione Generale per la Conservazione dei Pesci D’acqua Dolce Italiani; Ministero dell’ambiente e della tutela del territorio, Direzione per la protezione della natura: Rome, Italy, 2003. [Google Scholar]
- Vecsei, P.; Suciu, R.; Peterson, D. Threatened fishes of the world: Huso huso (Linnaeus, 1758) (Acipenseridae). Environ. Biol. Fishes 2002, 65, 363–365. [Google Scholar] [CrossRef]
- Freyhof, J.; Bergner, L.; Ford, M. Threatened Freshwater Fishes of the Mediterranean Basin Biodiversity Hotspot: Distribution, Extinction Risk and the Impact of Hydropower; EuroNatur: Radolfzell, Germany, 2020. [Google Scholar] [CrossRef]
- Friedrich, T.; Reinarts, F.; Gessner, J. Sturgeon re-introduction in the Upper and Middle Danube River Basin. J. Appl. Ichthyol 2019, 35, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Çiftci, Y.; Eroğlu, O.; Firidin, Ş. Mitochondrial Cytochrome b Sequence Variation in Three Sturgeon Species (A. stellatus Pallas, 1771, A. gueldenstaedtii Brandt, 1833, H. huso Linnaeus, 1758) from the Black Sea Coasts of Turkey. Turk. J. Fish. Aquat. Sci. 2013, 13, 291–303. [Google Scholar] [CrossRef]
- IUCN 2020, Versions 2020-1; The IUCN Red List of Threatened Species. 2020.
- Council of the European Communities. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Puzzi, C.M.; Trasforini, S.; Sartorelli, M.; Tamborini, D. Ticino River ecological corridor restoring and monitoring. Ital. J. Fresh. Icht. 2017. [Google Scholar]
- Drauch, A.M.; Rhodes, O.E., Jr. Genetic Evaluation of the Lake Sturgeon Reintroduction Program in the Mississippi and Missouri Rivers. N. Am. J. Fish. Manag. 2007, 27, 434–442. [Google Scholar] [CrossRef]
- Frankham, R. Genetic adaptation to captivity in species conservation programs. Mol. Ecol. 2008, 17, 325–333. [Google Scholar] [CrossRef]
- Fraser, D.J. How well can captive breeding programs conserve biodiversity? A review of salmonids. Evol. Appl. 2008, 1, 535–586. [Google Scholar] [CrossRef]
- Pollard, H.A.; Flagg, T.A. Guidelines for use of captive broodstocks in recovery efforts for Pacific salmon. Am. Fish. Soc. Symp. 2004, 44, 333–345. [Google Scholar]
- Gessner, J.; Chebanov, M.; Freyhof, J. Huso huso. IUCN 2013, Version 2013.2; IUCN Red List of Threatened Species. 2010. [Google Scholar]
- Rosenthal, H.; Gessner, J.; Bronzi, P. VIENNA DECLARATION on Global Sturgeon Conservation; WSCS Special Publication No. 9; WSCS: Winston-Salem, NC, USA, 2018. [Google Scholar]
- Rosenthal, H.; Pourkazemi, M. Ramsar declaration on global sturgeon conservation. J. Appl. Ichthyol. 2006, 22, 5–11. [Google Scholar]
- Genovesi, P.; Occhipinti Ambrogi, A.; Amori, G. Linee Guida per L’immissione di Specie Faunistiche; Ministero dell’Ambiente: Rome, Italy, 2007. [Google Scholar]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high-quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- McQuown, E.C.; Sloss, B.L.; Sheehan, R.J.; Rodzen, J.; Tranah, G.J.; May, B. Microsatellite analysis of genetic variation in sturgeon: New primer sequences for Scaphirhynchus and Acipenser. Trans. Am. Fish. Soc. 2000, 129, 1380–1388. [Google Scholar] [CrossRef]
- Henderson-Arzapalo, A.; King, T.L. Novel microsatellite markers for Atlantic sturgeon (Acipenser oxyrinchus) population delineation and broodstock management. Mol. Ecol. Res. 2002, 2, 437–439. [Google Scholar] [CrossRef]
- Zane, L.; Patarnello, T.; Ludwig, A.; Fontana, F.; Congiu, L. Isolation and characterization of microsatellites in the Adriatic sturgeon (Acipenser Naccarii). Mol. Ecol. Res. 2002, 2, 586–588. [Google Scholar] [CrossRef]
- Rajkov, J.; Shao, Z.; Berrebi, P. Evolution of polyploidy and functional diploidization in sturgeons: Microsatellite analysis in 10 sturgeon species. J. Hered. 2014, 105, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscari, E.; Vidotto, M.; Martini, D.; Papetti, C.; Ogden, R.; Congiu, L. Microsatellites from the genome and the trascriptome of the tetraploid Adriatic sturgeon, Acipenser naccarii (Bonaparte, 1836) and cross-species applicability to the diploid beluga sturgeon, Huso huso (Linneaus, 1758). J. Appl. Ichthyol 2015, 31, 977–983. [Google Scholar] [CrossRef]
- Congiu, L.; Pujolar, J.M.; Forlani, A.; Cenadelli, S.; Dupanloup, I.; Barbisan, F.; Galli, A.; Fontana, F. Managing Polyploidy in Ex Situ Conservation genetics: The Case of the Critically Endagered Adriatic Sturgeon (Acipenser Naccarii). PLoS ONE 2001, 6, e18249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic acids symposium series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.F.; Willis, D.P.M.; Shipley, P. MICRO-CHECKER: A software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 63–72. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel Sous Windows TM Pour la Genetique des Populations; Laboratoire Génome, Populations, Interactions: Montpellier, France, 2004. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Dent, A.E.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Zwickl, D.J. GARLI: Genetic Algorithm for Rapid Likelihood Inference. 2006. Available online: http://www.bio.utexas.edu/faculty/antisense/garli/Garli.html (accessed on 6 July 2020).
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J. MRBAYES 3: Bayesian Phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Doukakis, P.; Birstein, V.J.E.; De Salle, R. Intraspecific structure three caviar-producing sturgeons (Acipenser gueldenstadtii, A, stellatus and Huso huso) based on mitochondrial DNA analysis. J. Appl. Ichthyol 2005, 21, 457–460. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 1999, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.M.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seliger, C.; Zeiringer, B. River Connectivity, habitat fragmentation and related restoration measures. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 8, pp. 171–186. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Cooke, S.J. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Fang, Y.; Jawitz, J.W.; Yan, J.; Cui, B. River network connectivity and fish diversity. Sci. Total Environ. 2019, 689, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.D. Empirical Approaches to the Conservation of Margaritifera margaritifera. Ph.D. Thesis, Queen’s University, Belfast, Northern Ireland, 2010. [Google Scholar]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Jones, J.W.; Hallerman, E.M.; Neves, R.J. Genetic management guidelines for captive propagation of freshwater mussels (Unionoidea). J. Shellfish. Res. 2006, 25, 527–535. [Google Scholar] [CrossRef]
- Doukakis, P.; Babcock, E.A.; Pikitch, E.K.; Sharov, A.R.; Baimukhanov, M.; Erbulekov, S.; Bokova, Y.; Nimatov, A. Management and recovery options for Ural River Beluga sturgeon. Conserv. Biol. 2010, 24, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Kotlik, P.; Markova, S.; Choleva, L.; Bogutskaya, N.G.; Ekmekcis, F.G.; Ivanova, P.P. Divergence with gene flow between Ponto-Caspian refugia in an anadromous cyprinid Rutilus Frisii revealed bymultiple gene phylogeography. Mol. Ecol. 2008, 17, 1076–1088. [Google Scholar] [CrossRef]
- Balandina, L.G. Reproduction of beluga in the Azov basin. In Biological Bases of Sturgeon Farming; Barannikova, I.A., Berdichevsky, L.S., Eds.; Nauka: Moscow, Russian, 1983; pp. 216–223. [Google Scholar]
- Attard, C.R.M.; Möller, L.M.; Sasaki, M.; Hammer, M.P.; Bice, C.M.; Brauer, C.J.; Carvalho, D.C.; Harris, J.O.; Beheregaray, L.B. A novel holistic framework for genetic-based captive-breeding and reintroduction programs. Conserv. Biol. 2016, 30, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, F.; Quadroni, S.; Gentili, G.; Crosa, G. Thermal regime of a highly regulated Italian river (Ticino River) and implications for aquatic communities. J. Limnol. 2017, 76, 23–33. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Variability | FST Matrix | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Stock | N | NA | Ho | He | FIS | Stock | S1 | S2 | S3 | S4 |
| S1 | 47 | 3.5 | 0.45 ± 0.32 | 0.42 ± 0.26 | −0.05 | S1 | - | 0 | 0.2 | 0 |
| S2 | 12 | 2.6 | 0.57 ± 0.41 | 0.45 ± 0.31 | −0.22 | S2 | 0.28 | - | 0 | 0 |
| S3 | 9 | 2.6 | 0.57 ± 0.39 | 0.43 ± 0.28 | −0.26 | S3 | 0.09 | 0.21 | - | 0 |
| S4 | 14 | 2.6 | 0.61 ± 0.32 | 0.46 ± 0.22 | −0.30 | S4 | 0.21 | 0.25 | 0.19 | - |
| Tot | 82 | 4.5 | 0.50 ± 0.26 | 0.50 ± 0.23 | 0.00 | |||||
| Stock | N | Hap1 | Hap2 | Hap3 | Hap4 | Hap5 | Hap6 | Hap7 | Hap8 | Hap9 | Hap10 | Hap11 | Hap12 | Hap13 | H ± s.d. | π ± s.d. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 47 | 21 | 3 | 1 | 1 | 1 | 0.39 ± 0.11 | 0.0005 ± 0.0002 | ||||||||
| S2 | 12 | 12 | 0.00 | 0.00 | ||||||||||||
| S3 | 9 | 5 | 1 | 0.33 ± 0.21 | 0.0003 ± 0.0002 | |||||||||||
| S4 | 14 | 9 | 1 | 1 | 1 | 1 | 1 | 0.60 ± 0.15 | 0.0011 ± 0.0003 | |||||||
| Total | 82 | 21 | 3 | 1 | 1 | 1 | 12 | 14 | 1 | 1 | 1 | 1 | 1 | 1 | 0.78 ± 0.03 | 0.0079 ± 0.0007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antognazza, C.M.; Vanetti, I.; De Santis, V.; Bellani, A.; Di Francesco, M.; Puzzi, C.M.; Casoni, A.G.; Zaccara, S. Genetic Investigation of Four Beluga Sturgeon (Huso huso, L.) Broodstocks for its Reintroduction in the Po River Basin. Environments 2021, 8, 25. https://doi.org/10.3390/environments8040025
Antognazza CM, Vanetti I, De Santis V, Bellani A, Di Francesco M, Puzzi CM, Casoni AG, Zaccara S. Genetic Investigation of Four Beluga Sturgeon (Huso huso, L.) Broodstocks for its Reintroduction in the Po River Basin. Environments. 2021; 8(4):25. https://doi.org/10.3390/environments8040025
Chicago/Turabian StyleAntognazza, Caterina M., Isabella Vanetti, Vanessa De Santis, Adriano Bellani, Monica Di Francesco, Cesare M. Puzzi, Andrea G. Casoni, and Serena Zaccara. 2021. "Genetic Investigation of Four Beluga Sturgeon (Huso huso, L.) Broodstocks for its Reintroduction in the Po River Basin" Environments 8, no. 4: 25. https://doi.org/10.3390/environments8040025
APA StyleAntognazza, C. M., Vanetti, I., De Santis, V., Bellani, A., Di Francesco, M., Puzzi, C. M., Casoni, A. G., & Zaccara, S. (2021). Genetic Investigation of Four Beluga Sturgeon (Huso huso, L.) Broodstocks for its Reintroduction in the Po River Basin. Environments, 8(4), 25. https://doi.org/10.3390/environments8040025