
An In Silico and In Vitro Study for Investigating Estrogenic Endocrine Effects of Emerging Persistent Pollutants Using Primary Hepatocytes from Grey Mullet (Mugil cephalus)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Endocrine Disruptome Screening Tool
2.2. Hepatocyte Isolation and Primary Cell Culture
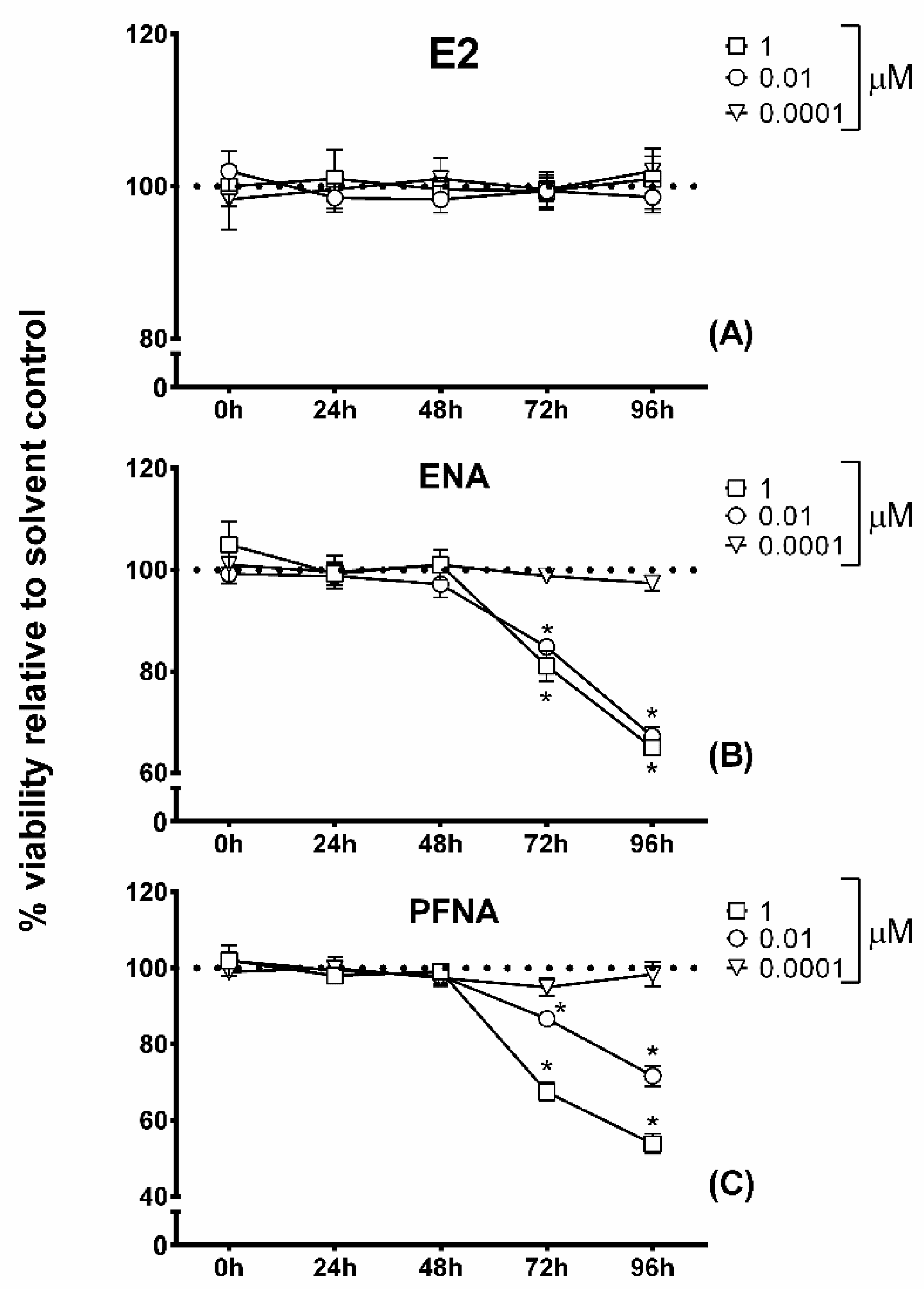
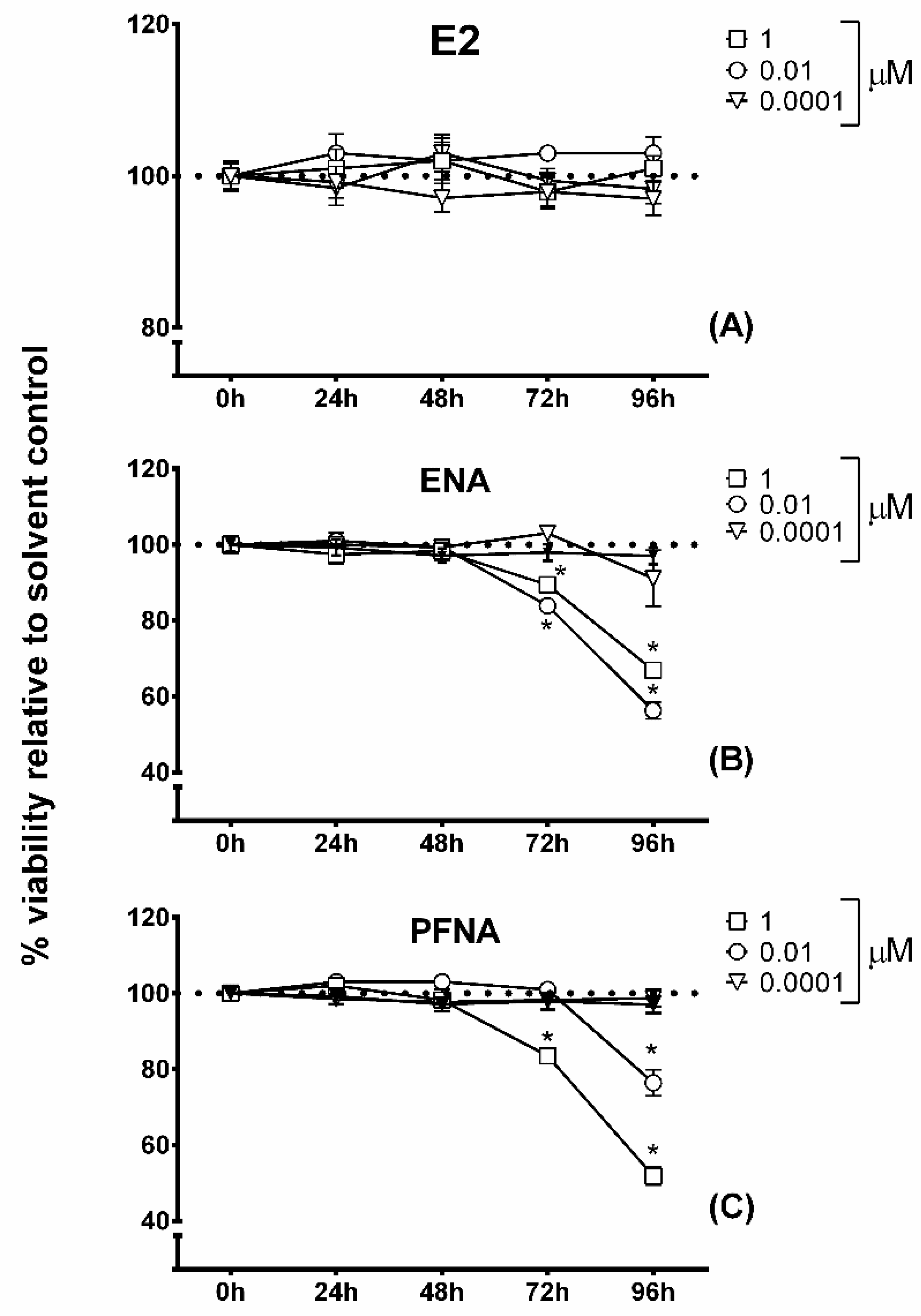
2.3. MTT Cytotoxicity Assay
2.4. Alamar Blue Assay
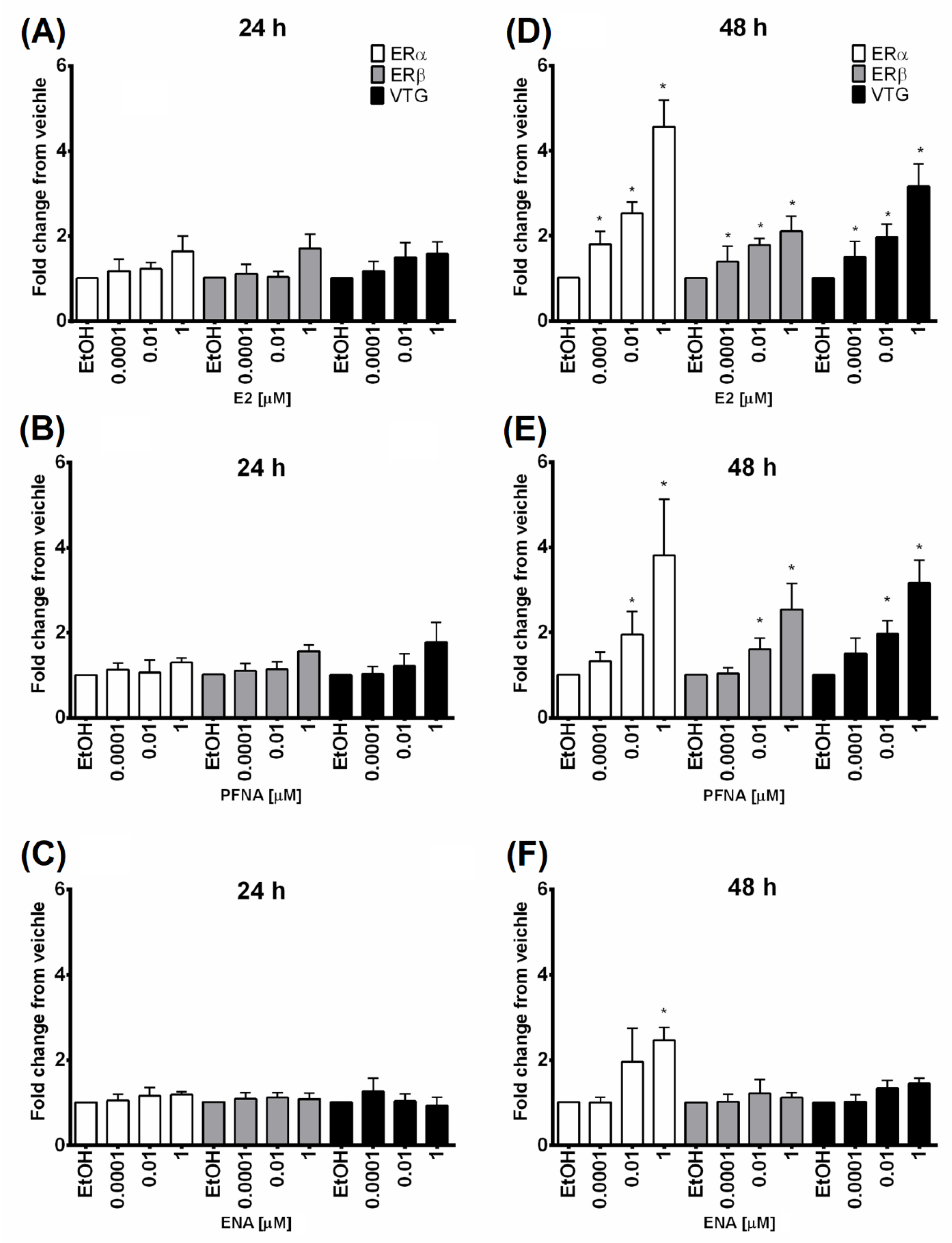
2.5. Quantitative Realtime PCR (q-PCR)
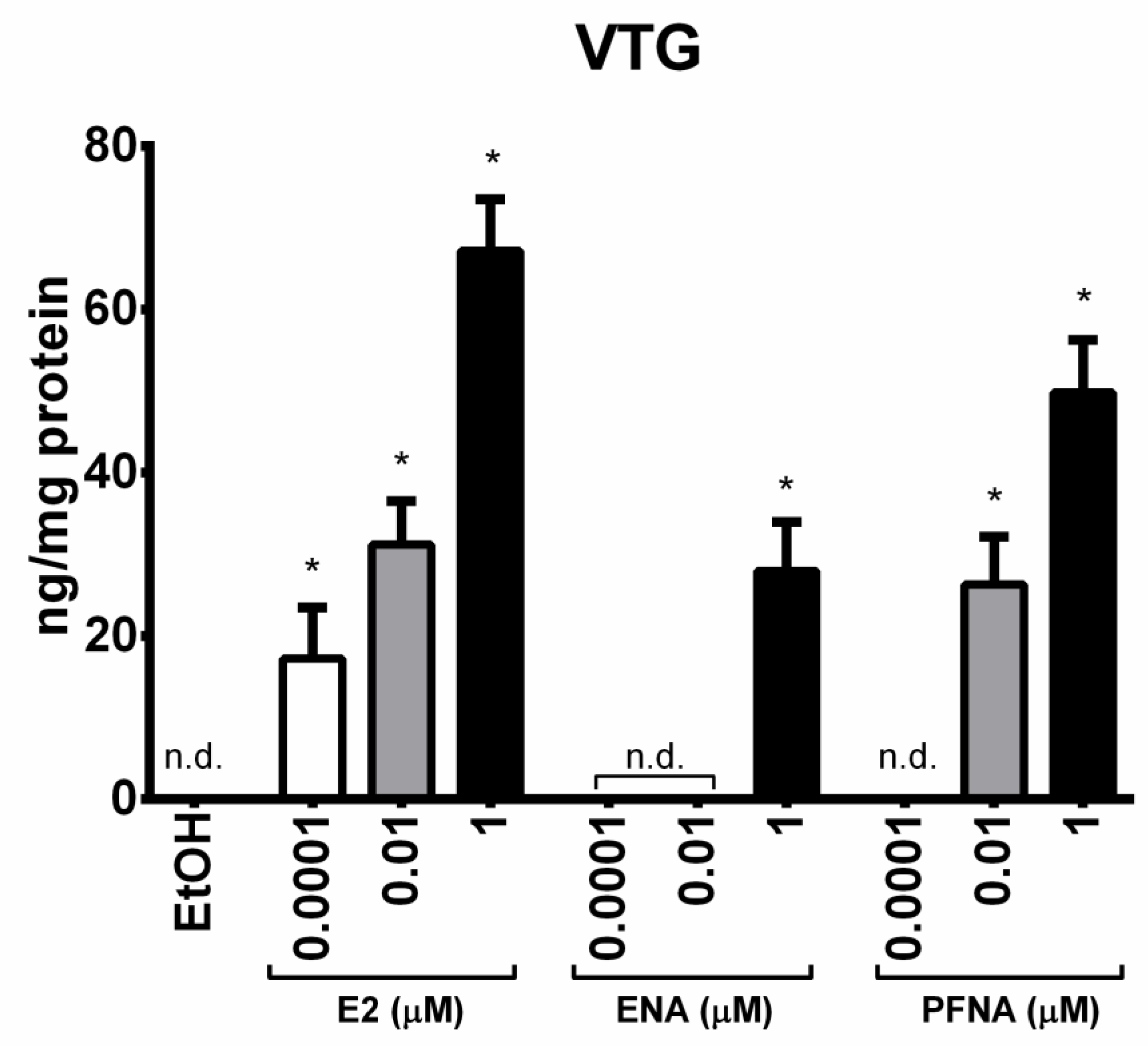
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, A.; Weerakoon, D.; Lim, E.; Padhye, L.P. Fate of environmental pollutants. Water Environ. Res. 2019, 91, 1294–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Cai, Z.-H.; Zhu, X.-S. Are endocrine disruptors among the causes of the deterioration of aquatic biodiversity? Integr. Environ. Assess. Manag. 2010, 6, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Celino-Brady, F.T.; Lerner, D.T.; Seale, A.P. Experimental Approaches for Characterizing the Endocrine-Disrupting Effects of Environmental Chemicals in Fish. Front. Endocrinol. 2021, 11, 619361. [Google Scholar] [CrossRef] [PubMed]
- Corsini, E.; Luebke, R.W.; Germolec, D.R.; DeWitt, J.C. Perfluorinated compounds: Emerging POPs with potential immunotoxicity. Toxicol. Lett. 2014, 230, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, J.L.; Hooda, P.S.; Barker, J.; Barton, S.; Swinden, J. Ecotoxic pharmaceuticals, personal care products, and other emerging contaminants: A review of environmental, receptor-mediated, developmental, and epigenetic toxicity with discussion of proposed toxicity to humans. Crit. Rev. Environ. Sci. Technol. 2015, 46, 336–381. [Google Scholar] [CrossRef] [Green Version]
- OECD. Summary Report on the New Comprehensive Global Database of Per- and Polyfluoroalkyl Substances (PFASs); Publications Series on Risk Management: Paris, France, 2018; No. 39. [Google Scholar]
- Houde, M.; Martin, J.W.; Letcher, R.J.; Solomon, K.R.; Muir, D.C.G. Biological Monitoring of Polyfluoroalkyl Substances: A Review. Environ. Sci. Technol. 2006, 40, 3463–3473. [Google Scholar] [CrossRef]
- Valsecchi, S.; Rusconi, M.; Polesello, S. Determination of perfluorinated compounds in aquatic organisms: A review. Anal. Bioanal. Chem. 2012, 405, 143–157. [Google Scholar] [CrossRef]
- White, S.S.; Fenton, S.E.; Hines, E.P. Endocrine disrupting properties of perfluorooctanoic acid. J. Steroid Biochem. Mol. Biol. 2011, 127, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Song, B.; Xiao, R.; Zeng, G.; Gong, J.; Chen, M.; Xu, P.; Zhang, P.; Shen, M.; Yi, H. Assessing the human health risks of perfluorooctane sulfonate by in vivo and in vitro studies. Environ. Int. 2019, 126, 598–610. [Google Scholar] [CrossRef]
- Fent, K.; Weston, A.A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [Google Scholar] [CrossRef]
- Liu, N.; Jin, X.; Feng, C.; Wang, Z.; Wu, F.; Johnson, A.C.; Xiao, H.; Hollert, H.; Giesy, J.P. Ecological risk assessment of fifty pharmaceuticals and personal care products (PPCPs) in Chinese surface waters: A proposed multiple-level system. Environ. Int. 2020, 136, 105454. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Silva, L.; Laranjeiro, C.; Lino, C.; Pena, A. Selected Pharmaceuticals in Different Aquatic Compartments: Part I—Source, Fate and Occurrence. Molecules 2020, 25, 1026. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman, C.U., Jr.; Mohan, D. Pharmaceuticals of Emerging Concern in Aquatic Systems: Chemistry, Occurrence, Effects, and Removal Methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [Green Version]
- Zuccato, E.; Castiglioni, S.; Fanelli, R.; Reitano, G.; Bagnati, R.; Chiabrando, C.; Pomati, F.; Rossetti, C.; Calamari, D. Pharmaceuticals in the Environment in Italy: Causes, Occurrence, Effects and Control. Environ. Sci. Pollut. Res. 2006, 13, 15–21. [Google Scholar] [CrossRef]
- Calamari, D.; Zuccato, E.; Castiglioni, S.; Bagnati, R.; Fanelli, R. Strategic Survey of Therapeutic Drugs in the Rivers Po and Lambro in Northern Italy. Environ. Sci. Technol. 2003, 37, 1241–1248. [Google Scholar] [CrossRef]
- Isidori, M.; Bellotta, M.; Cangiano, M.; Parrella, A. Estrogenic activity of pharmaceuticals in the aquatic environment. Environ. Int. 2009, 35, 826–829. [Google Scholar] [CrossRef] [PubMed]
- Laurenson, J.P.; Bloom, R.A.; Page, S.; Sadrieh, N. Ethinyl Estradiol and Other Human Pharmaceutical Estrogens in the Aquatic Environment: A Review of Recent Risk Assessment Data. AAPS J. 2014, 16, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Cocci, P.; Mozzicafreddo, M.; Angeletti, M.; Mosconi, G.; Palermo, F.A. In silico prediction and in vivo analysis of antiestrogenic potential of 2-isopropylthioxanthone (2-ITX) in juvenile goldfish (Carassius auratus). Ecotoxicol. Environ. Saf. 2016, 133, 202–210. [Google Scholar] [CrossRef]
- Cocci, P.; Palermo, F.A.; Quassinti, L.; Bramucci, M.; Miano, A.; Mosconi, G. Determination of estrogenic activity in the river Chienti (Marche Region, Italy) by using in vivo and in vitro bioassays. J. Environ. Sci. 2016, 43, 48–53. [Google Scholar] [CrossRef]
- Palermo, F.A.; Cocci, P.; Angeletti, M.; Polzonetti-Magni, A.; Mosconi, G. PCR–ELISA detection of estrogen receptor β mRNA expression and plasma vitellogenin induction in juvenile sole (Solea solea) exposed to waterborne 4-nonylphenol. Chemosphere 2012, 86, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Palermo, F.A.; Mosconi, G.; Angeletti, M.; Polzonetti-Magni, A.M. Assessment of Water Pollution in the Tronto River (Italy) by Applying Useful Biomarkers in the Fish Model Carassius auratus. Arch. Environ. Contam. Toxicol. 2008, 55, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Capriotti, M.; Mosconi, G.; Palermo, F.A. Effects of endocrine disrupting chemicals on estrogen receptor alpha and heat shock protein 60 gene expression in primary cultures of loggerhead sea turtle (Caretta caretta) erythrocytes. Environ. Res. 2017, 158, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Blazer, V.S.; Walsh, H.L.; Shaw, C.H.; Iwanowicz, L.R.; Braham, R.P.; Mazik, P.M. Indicators of exposure to estrogenic compounds at Great Lakes Areas of Concern: Species and site comparisons. Environ. Monit. Assess. 2018, 190, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Cocci, P.; Palermo, F.A.; Pucciarelli, S.; Miano, A.; Cuccioloni, M.; Angeletti, M.; Roncarati, A.; Mosconi, G. Identification, partial characterization, and use of grey mullet (Mugil cephalus) vitellogenins for the development of ELISA and biosensor immunoassays. Int. Aquat. Res. 2019, 11, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Tollefsen, K.-E.; Bratsberg, E.; Bøyum, O.; Finne, E.F.; Gregersen, I.K.; Hegseth, M.; Sandberg, C.; Hylland, K. Use of fish in vitro hepatocyte assays to detect multi-endpoint toxicity in Slovenian river sediments. Mar. Environ. Res. 2006, 62, S356–S359. [Google Scholar] [CrossRef]
- Tollefsen, K.-E.; Mathisen, R.; Stenersen, J. Induction of vitellogenin synthesis in an Atlantic salmon (Salmo salar) hepatocyte culture: A sensitivein vitrobioassay for the oestrogenic and anti-oestrogenic activity of chemicals. Biomarkers 2003, 8, 394–407. [Google Scholar] [CrossRef]
- Cocci, P.; Capriotti, M.; Mosconi, G.; Campanelli, A.; Frapiccini, E.; Marini, M.; Caprioli, G.; Sagratini, G.; Aretusi, G.; Palermo, F.A. Alterations of gene expression indicating effects on estrogen signaling and lipid homeostasis in seabream hepatocytes exposed to extracts of seawater sampled from a coastal area of the central Adriatic Sea (Italy). Mar. Environ. Res. 2017, 123, 25–37. [Google Scholar] [CrossRef]
- Pesonen, M.; Andersson, T.B. Fish primary hepatocyte culture; an important model for xenobiotic metabolism and toxicity studies. Aquat. Toxicol. 1997, 37, 253–267. [Google Scholar] [CrossRef]
- Ellesat, K.S.; Yazdani, M.; Holth, T.F.; Hylland, K. Species-dependent sensitivity to contaminants: An approach using primary hepatocyte cultures with three marine fish species. Mar. Environ. Res. 2011, 72, 216–224. [Google Scholar] [CrossRef]
- Zuccato, E.; Castiglioni, S.; Fanelli, R. Identification of the pharmaceuticals for human use contaminating the Italian aquatic environment. J. Hazard. Mater. 2005, 122, 205–209. [Google Scholar] [CrossRef]
- Burcea, A.; Boeraş, I.; Mihuţ, C.-M.; Bănăduc, D.; Matei, C.; Curtean-Bănăduc, A. Adding the Mureş River Basin (Transylvania, Romania) to the List of Hotspots with High Contamination with Pharmaceuticals. Sustainability 2020, 12, 10197. [Google Scholar] [CrossRef]
- Fick, J.; Söderström, H.; Lindberg, R.H.; Phan, C.; Tysklind, M.; Larsson, D.J. Contamination of Surface, Ground and Drinking Water from Pharmaceutical Production. Environ. Toxicol. Chem. 2009, 28, 2522–2527. [Google Scholar] [CrossRef]
- Fliedner, A.; Rüdel, H.; Dreyer, A.; Pirntke, U.; Koschorreck, J. Chemicals of emerging concern in marine specimens of the German Environmental Specimen Bank. Environ. Sci. Eur. 2020, 32, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Lohmann, R.; Sunderland, E.M. Poly- and Perfluoroalkyl Substances in Seawater and Plankton from the Northwestern Atlantic Margin. Environ. Sci. Technol. 2019, 53, 12348–12356. [Google Scholar] [CrossRef] [PubMed]
- Kolšek, K.; Mavri, J.; Dolenc, M.S.; Gobec, S.; Turk, S. Endocrine Disruptome—An Open Source Prediction Tool for Assessing Endocrine Disruption Potential through Nuclear Receptor Binding. J. Chem. Inf. Model. 2014, 54, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Mosconi, G.; Arukwe, A.; Mozzicafreddo, M.; Angeletti, M.; Aretusi, G.; Palermo, F.A. Effects of Diisodecyl Phthalate on PPAR: RXR-Dependent Gene Expression Pathways in Sea Bream Hepatocytes. Chem. Res. Toxicol. 2015, 28, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Palermo, F.A.; Cocci, P.; Mozzicafreddo, M.; Arukwe, A.; Angeletti, M.; Aretusi, G.; Mosconi, G. Tri-m-cresyl phosphate and PPAR/LXR interactions in seabream hepatocytes: Revealed by computational modeling (docking) and transcriptional regulation of signaling pathways. Toxicol. Res. 2015, 5, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Celiz, M.D.; Pérez, S.; Barceló, D.; Aga, D.S. Trace Analysis of Polar Pharmaceuticals in Wastewater by LC-MS-MS: Comparison of Membrane Bioreactor and Activated Sludge Systems. J. Chromatogr. Sci. 2009, 47, 19–25. [Google Scholar] [CrossRef]
- Kim, J.-W.; Tue, N.M.; Isobe, T.; Misaki, K.; Takahashi, S.; Viet, P.H.; Tanabe, S. Contamination by perfluorinated compounds in water near waste recycling and disposal sites in Vietnam. Environ. Monit. Assess. 2012, 185, 2909–2919. [Google Scholar] [CrossRef]
- Loos, R.; Wollgast, J.; Huber, T.; Hanke, G. Polar herbicides, pharmaceutical products, perfluorooctanesulfonate (PFOS), perfluorooctanoate (PFOA), and nonylphenol and its carboxylates and ethoxylates in surface and tap waters around Lake Maggiore in Northern Italy. Anal. Bioanal. Chem. 2007, 387, 1469–1478. [Google Scholar] [CrossRef]
- Smeets, J.M.; Rankouhi, T.R.; Nichols, K.M.; Komen, H.; Kaminski, N.E.; Giesy, J.P.; van den Berg, M. In VitroVitellogenin Production by Carp (Cyprinus carpio) Hepatocytes as a Screening Method for Determining (Anti)Estrogenic Activity of Xenobiotics. Toxicol. Appl. Pharmacol. 1999, 157, 68–76. [Google Scholar] [CrossRef]
- Cocci, P.; Mosconi, G.; Palermo, F.A. Changes in expression of microRNA potentially targeting key regulators of lipid metabolism in primary gilthead sea bream hepatocytes exposed to phthalates or flame retardants. Aquat. Toxicol. 2019, 209, 81–90. [Google Scholar] [CrossRef]
- Cocci, P.; Mosconi, G.; Palermo, F.A. Pregnane X receptor (PXR) signaling in seabream primary hepatocytes exposed to extracts of seawater samples collected from polycyclic aromatic hydrocarbons (PAHs)-contaminated coastal areas. Mar. Environ. Res. 2017, 130, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Ribecco, C.; Baker, M.E.; Šášik, R.; Zuo, Y.; Hardiman, G.; Carnevali, O. Biological effects of marine contaminated sediments on Sparus aurata juveniles. Aquat. Toxicol. 2011, 104, 308–316. [Google Scholar] [CrossRef]
- Vieira, F.A.; Pinto, P.I.; Guerreiro, P.M.; Power, D.M. Divergent responsiveness of the dentary and vertebral bone to a selective estrogen-receptor modulator (SERM) in the teleost Sparus auratus. Gen. Comp. Endocrinol. 2012, 179, 421–427. [Google Scholar] [CrossRef]
- Cabas, I.; Liarte, S.; García-Alcázar, A.; Meseguer, J.; Mulero, V.; García-Ayala, A. 17α-Ethynylestradiol alters the immune response of the teleost gilthead seabream (Sparus aurata L.) both in vivo and in vitro. Dev. Comp. Immunol. 2012, 36, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, J.; Borrel, M.; Bermejo-Nogales, A.; Benedito-Palos, L.; Saera-Vila, A.; Calduch-Giner, J.A.; Kaushik, S. Dietary oils mediate cortisol kinetics and the hepatic mRNA expression profile of stress-responsive genes in gilthead sea bream (Sparus aurata) exposed to crowding stress. Implications on energy homeostasis and stress susceptibility. Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navas, J.M.; Segner, H. Vitellogenin synthesis in primary cultures of fish liver cells as endpoint for in vitro screening of the (anti)estrogenic activity of chemical substances. Aquat. Toxicol. 2006, 80, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, P.E.; Li, P.; Lenhart, J.R.; Chappell, M.C.; Brosnihan, K.B. Estrogen Regulation of Angiotensin-Converting Enzyme mRNA. Hypertension 1999, 33, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Zilberman, J.M.; Licy, Y.L.; Sartori-Vallinoti, J.C.; Iliescu, R.; Reckelhoff, J.F. Angiotensin converting enzyme inhibitor upreg-ulates the expression of estrogen receptors in the kidney in old female rats. Faseb J. 2008, 22, 941–948. [Google Scholar] [CrossRef]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl Acids: A Review of Monitoring and Toxicological Findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [Green Version]
- DRBC. Contaminants of Emerging Concern in the Delaware River Basin. 2016. Available online: http://www.nj.gov/drbc/quality/reports/emerging/ (accessed on 16 April 2021).
- Lindstrom, A.B.; Strynar, M.J.; Libelo, E.L. Polyfluorinated Compounds: Past, Present, and Future. Environ. Sci. Technol. 2011, 45, 7954–7961. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, S.K. Effect of gestational exposure to perfluorononanoic acid on neonatal mice testes. J. Appl. Toxicol. 2019, 39, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Benninghoff, A.D.; Bisson, W.H.; Koch, D.C.; Ehresman, D.J.; Kolluri, S.K.; Williams, D.E. Estrogen-Like Activity of Perfluoroalkyl Acids In Vivo and Interaction with Human and Rainbow Trout Estrogen Receptors In Vitro. Toxicol. Sci. 2010, 120, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Jantzen, C.E.; Annunziato, K.A.; Bugel, S.M.; Cooper, K.R. PFOS, PFNA, and PFOA sub-lethal exposure to embryonic zebrafish have different toxicity profiles in terms of morphometrics, behavior and gene expression. Aquat. Toxicol. 2016, 175, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurima-Romet, M.; Huang, H.S.; Paul, C.J.; Thomas, B.H. Enalapril cytotoxicity in primary cultures of rat hepatocytes. II. Role of glutathione. Toxicol. Lett. 1991, 58, 269–277. [Google Scholar] [CrossRef]
- Purclutepe, O.; Iskender, G.; Kiper, H.D.; Tezcanli, B.; Selvi, N.; Avci, C.B.; Kosova, B.; Gokbulut, A.A.; Sahin, F.; Baran, Y.; et al. Enalapril-induced apoptosis of acute promyelocytic leukaemia cells involves STAT5A. Anticancer. Res. 2012, 32, 2885–2893. [Google Scholar] [PubMed]
- Louisse, J.; Rijkers, D.; Stoopen, G.; Janssen, A.; Staats, M.; Hoogenboom, R.; Kersten, S.; Peijnenburg, A. Perfluorooctanoic acid (PFOA), perfluorooctane sulfonic acid (PFOS), and perfluorononanoic acid (PFNA) increase triglyceride levels and decrease cholesterogenic gene expression in human HepaRG liver cells. Arch. Toxicol. 2020, 94, 3137–3155. [Google Scholar] [CrossRef]
- Aerni, H.-R.; Kobler, B.; Rutishauser, B.V.; Wettstein, F.E.; Fischer, R.; Giger, W.; Hungerbühler, A.; Marazuela, M.D.; Peter, A.; Schönenberger, R.; et al. Combined biological and chemical assessment of estrogenic activities in wastewater treatment plant effluents. Anal. Bioanal. Chem. 2003, 378, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Lozano, N.; Rice, C.P.; Pagano, J.; Zintek, L.; Barber, L.B.; Murphy, E.W.; Nettesheim, T.; Minarik, T.; Schoenfuss, H.L. Concentration of organic contaminants in fish and their biological effects in a wastewater-dominated urban stream. Sci. Total. Environ. 2012, 420, 191–201. [Google Scholar] [CrossRef]
- Madsen, L.L.; Korsgaard, B.; Pedersen, K.L.; Bjerregaard, L.B.; Aagaard, T.; Bjerregaard, P. Vitellogenin as biomarker for estrogenicity in flounder Platichthys flesus in the field and exposed to 17α-ethinylestradiol via food and water in the laboratory. Mar. Environ. Res. 2013, 92, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; Jensen, K.M.; Kahl, M.D.; Korte, J.J.; Makynen, E.A. Description and evaluation of a short-term reproduction test with the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2001, 20, 1276–1290. [Google Scholar] [CrossRef] [PubMed]
- Hinck, J.E.; Blazer, V.S.; Denslow, N.D.; Echols, K.R.; Gross, T.S.; May, T.W.; Anderson, P.J.; Coyle, J.J.; Tillitt, D.E. Chemical contaminants, health indicators, and reproductive biomarker responses in fish from the Colorado River and its tributaries. Sci. Total. Environ. 2007, 378, 376–402. [Google Scholar] [CrossRef]
- Pomatto, V.; Palermo, F.; Mosconi, G.; Cottone, E.; Cocci, P.; Nabissi, M.; Borgio, L.; Polzonetti-Magni, A.M.; Franzoni, M.F. Xenoestrogens elicit a modulation of endocannabinoid system and estrogen receptors in 4NP treated goldfish, Carassius auratus. Gen. Comp. Endocrinol. 2011, 174, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.K.; Bjørnstad, A.; Sundt, R.C.; Taban, I.C.; Pampanin, D.M.; Andersen, O.K. Comparison of protein expression in plasma from nonylphenol and bisphenol A-exposed Atlantic cod (Gadus morhua) and turbot (Scophthalmus maximus) by use of SELDI-TOF. Aquat. Toxicol. 2006, 78, S25–S33. [Google Scholar] [CrossRef]
- Petersen, K.; Tollefsen, K.E. Assessing combined toxicity of estrogen receptor agonists in a primary culture of rainbow trout (Oncorhynchus mykiss) hepatocytes. Aquat. Toxicol. 2011, 101, 186–195. [Google Scholar] [CrossRef]
- Griffin, L.B.; January, K.E.; Ho, K.W.; Cotter, K.A.; Callard, G.V. Morpholino-Mediated Knockdown of ERα, ERβa, and ERβb mRNAs in Zebrafish (Danio rerio) Embryos Reveals Differential Regulation of Estrogen-Inducible Genes. Endocrinology 2013, 154, 4158–4169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, E.R.; Habibi, H.R. Functional Significance of Nuclear Estrogen Receptor Subtypes in the Liver of Goldfish. Endocrinology 2010, 151, 1668–1676. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Imamura, E.; Shinohara, H.; Kiri, K.; Muramatsu, Y.; Harada, A.; Takada, H. Occurrence and Sources of Perfluorinated Surfactants in Rivers in Japan. Environ. Sci. Technol. 2008, 42, 6566–6572. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Genbank | Product Size | Efficiency (%) |
|---|---|---|---|---|
| ERα | CTGGTGCCTTCTCTTTTTGC TGTCTGATGTGGGAGAGCAG | AF136979 | 181 | 96.85 |
| ERβ | TGTCATCGGGCGGGAAGG GCTCTTACGGCGGTTCTTGTCT | AF136980 | 188 | 91.74 |
| VTG | CTGCTGAAGAGGGACCAGAC TTGCCTGCAGGATGATGATA | AF210428 | 158 | 96.31 |
| 18s rRNA | GCATTTATCAGACCCAAAACC AGTTGATAGGGCAGACATTCG | AY993930 | 135 | 98.65 |
| CAS | Name | Structure | Receptor/Predictions Free Binding Energies (kcal mol−1) |
|---|---|---|---|
| 50-28-2 | E2 |  | ERα+ (−10.2) |
| ERα− (−10.0) | |||
| ERβ+ (−9.6) | |||
| ERβ− (−8.8) | |||
| 75847-73-3 | ENA |  | ERα+ (−8.3) |
| ERα− (−7.7) | |||
| ERβ+ (−6.8) | |||
| ERβ− (−7.8) | |||
| 375-95-1 | PFNA |  | ERα+ (−8.9) |
| ERα− (−9.1) | |||
| ERβ+ (−9.7) | |||
| ERβ− (−8.9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocci, P.; Mosconi, G.; Palermo, F.A. An In Silico and In Vitro Study for Investigating Estrogenic Endocrine Effects of Emerging Persistent Pollutants Using Primary Hepatocytes from Grey Mullet (Mugil cephalus). Environments 2021, 8, 58. https://doi.org/10.3390/environments8060058
Cocci P, Mosconi G, Palermo FA. An In Silico and In Vitro Study for Investigating Estrogenic Endocrine Effects of Emerging Persistent Pollutants Using Primary Hepatocytes from Grey Mullet (Mugil cephalus). Environments. 2021; 8(6):58. https://doi.org/10.3390/environments8060058
Chicago/Turabian StyleCocci, Paolo, Gilberto Mosconi, and Francesco A. Palermo. 2021. "An In Silico and In Vitro Study for Investigating Estrogenic Endocrine Effects of Emerging Persistent Pollutants Using Primary Hepatocytes from Grey Mullet (Mugil cephalus)" Environments 8, no. 6: 58. https://doi.org/10.3390/environments8060058
APA StyleCocci, P., Mosconi, G., & Palermo, F. A. (2021). An In Silico and In Vitro Study for Investigating Estrogenic Endocrine Effects of Emerging Persistent Pollutants Using Primary Hepatocytes from Grey Mullet (Mugil cephalus). Environments, 8(6), 58. https://doi.org/10.3390/environments8060058