
Seasonal Spectral Separation of Western Snowberry and Wolfwillow in Grasslands with Field Spectroradiometer and Simulated Multispectral Bands


Abstract
:1. Introduction
2. Materials and Methods
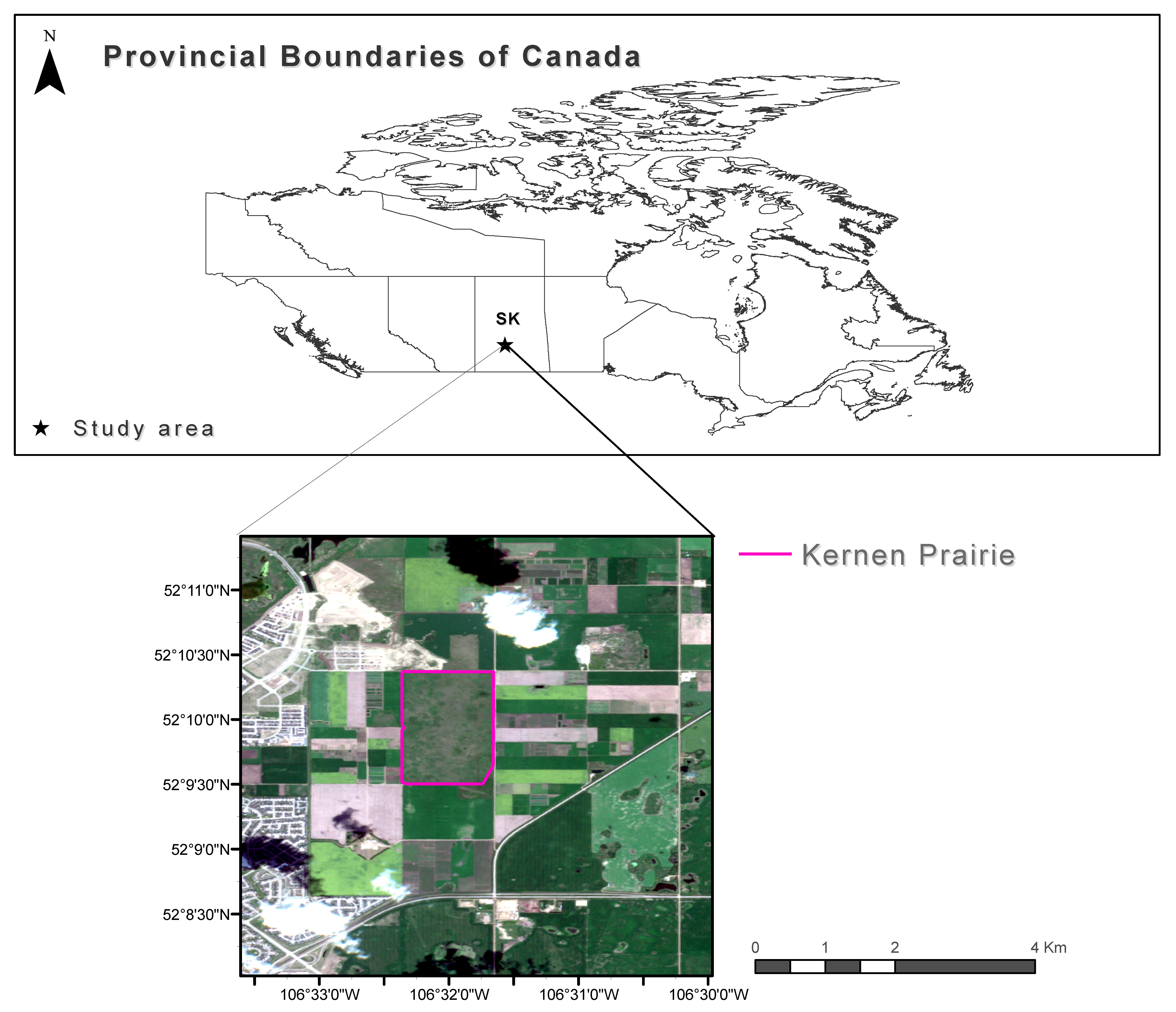
2.1. Study Area and Case Study Species
2.2. Data Collection
2.3. Data Processing
2.3.1. Calculation of Separability Metrics
2.3.2. Thresholding and Selection of Important Wavelength Regions
2.3.3. Broadband Spectral Difference between Shrub Species
3. Results
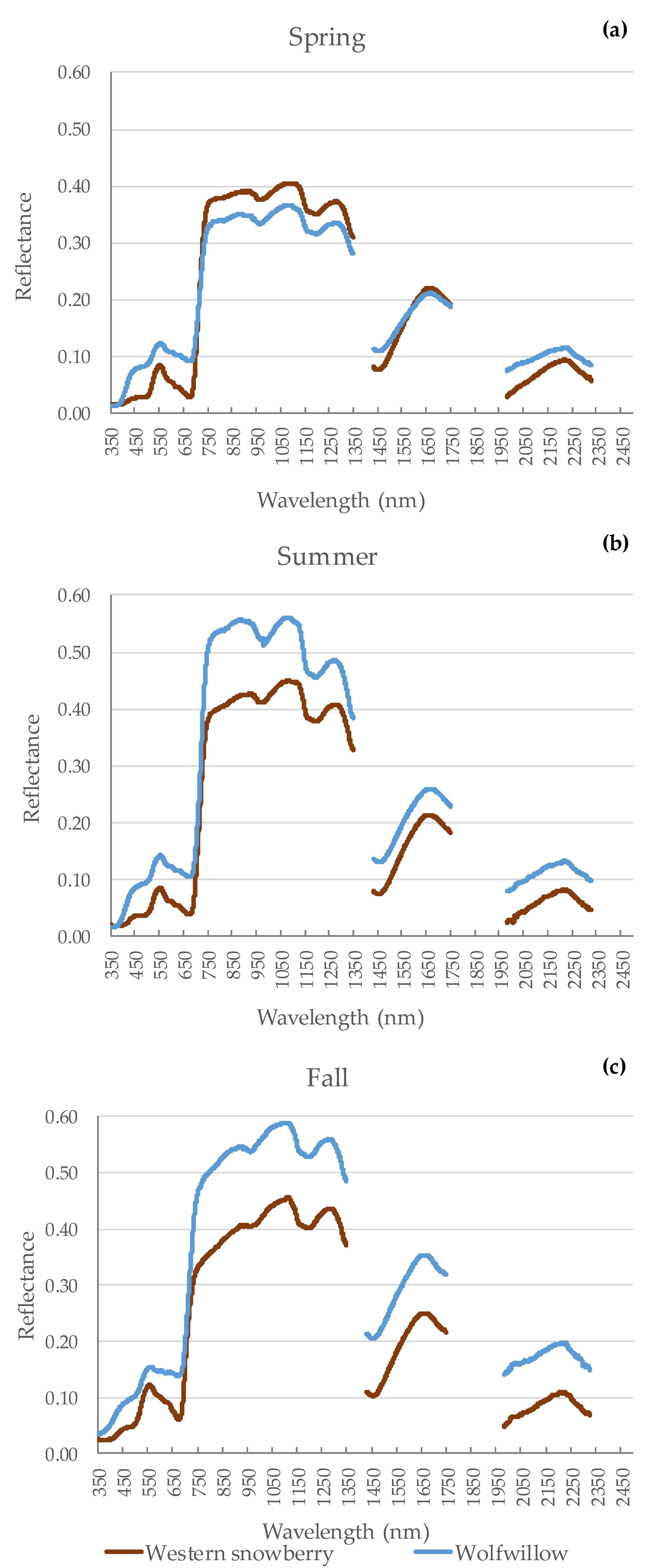
3.1. Seasonal Spectra of Shrub Species
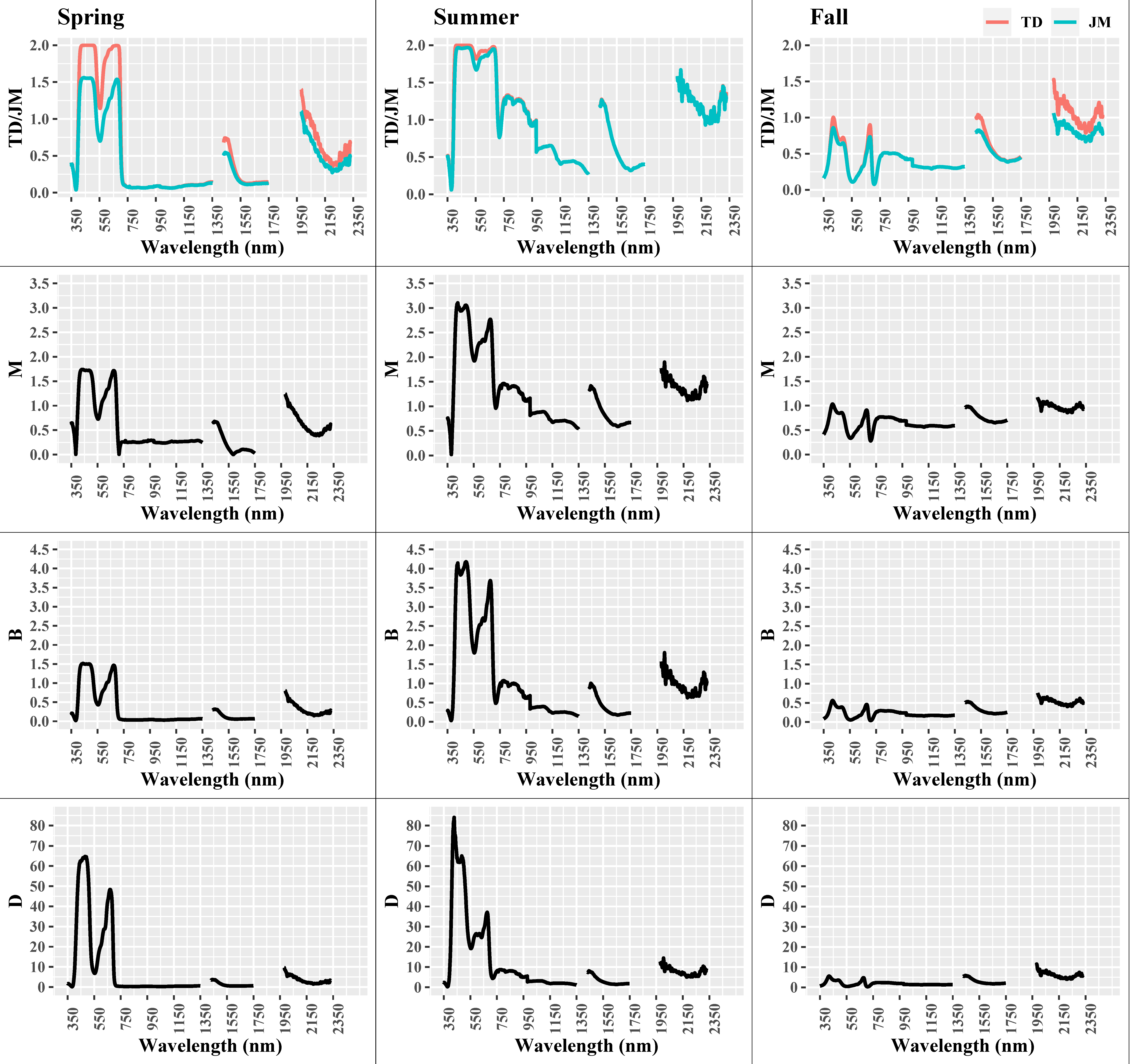
3.2. Hyperspectral Separability of Shrub Species
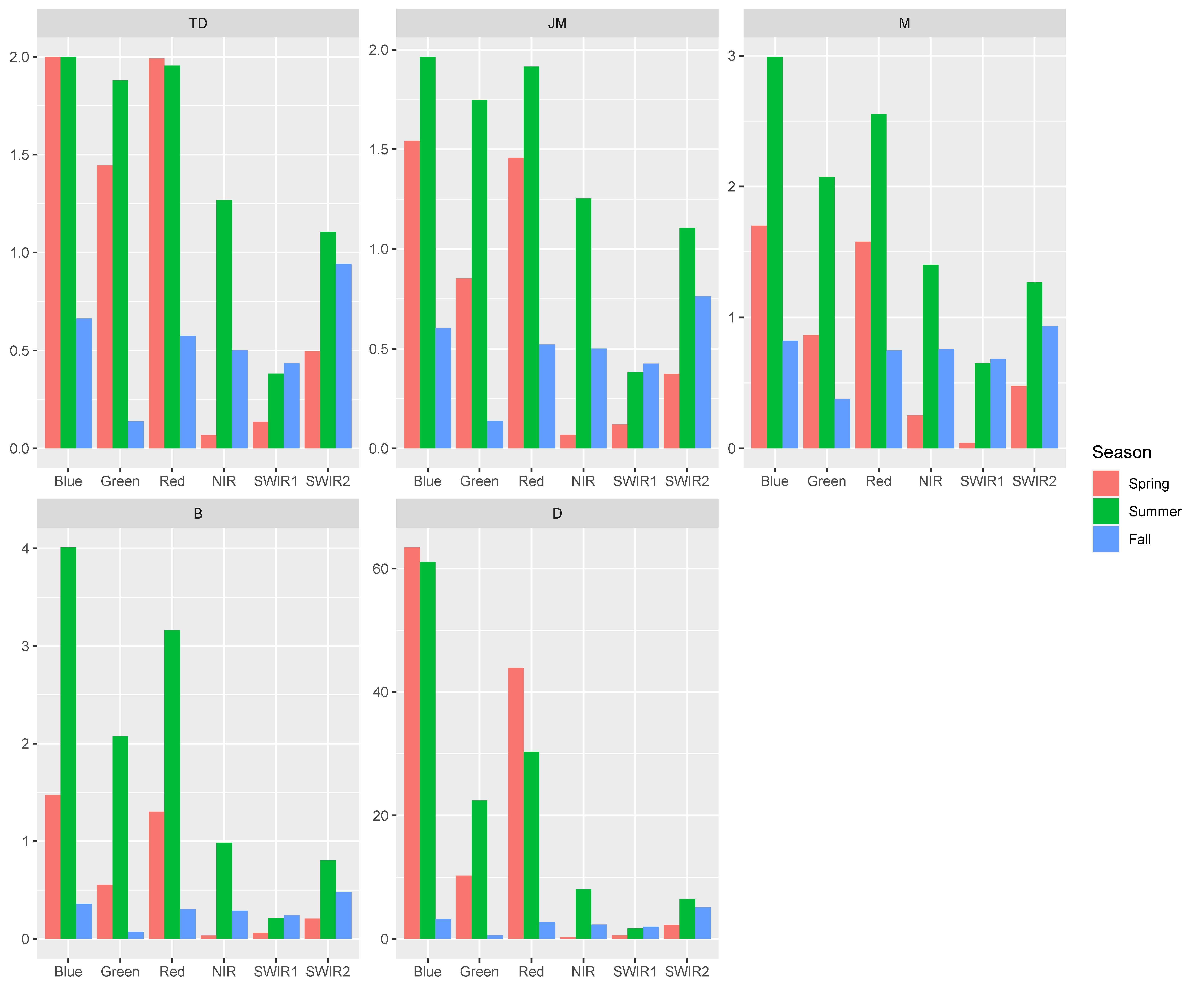
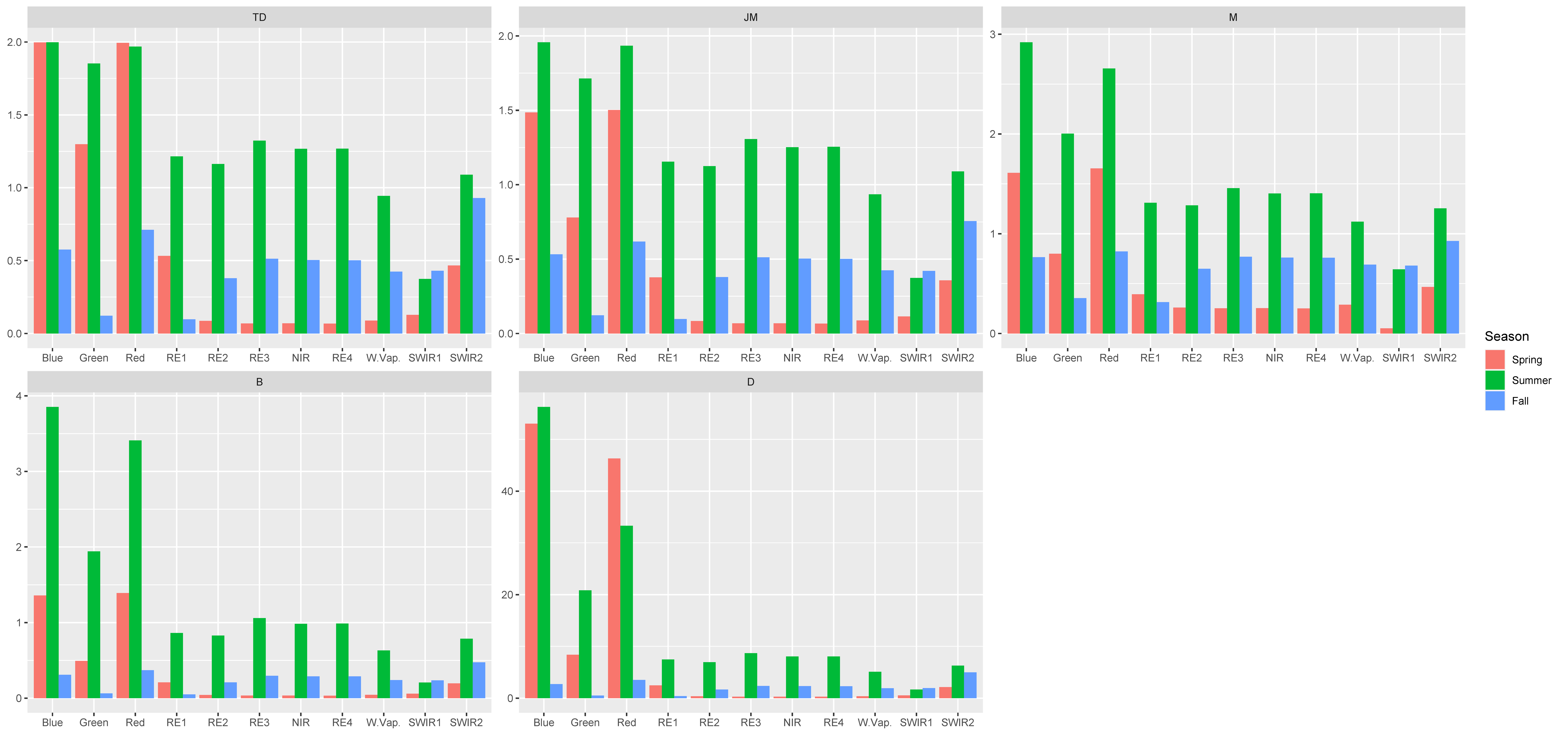
3.3. Broadband Simulation and Shrub Species Spectral Band Difference
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| B | Bhattacharyya distance |
| D | Divergence |
| JM | Jeffries–Matusita distance |
| M | M-Statistic |
| NIR | Near Infrared |
| RE | Red Edge |
| SWIR | Shortwave Infrared |
| TD | Transformed Divergence |
| WPE | Woody Plant Encroachment |
References
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More important for ecosystem services than you might think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Gang, C.; Zhou, W.; Chen, Y.; Wang, Z.; Sun, Z.; Li, J.; Qi, J.; Odeh, I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody Plant Encroachment-Causes and Concequences. In Rangeland Systems-Processes, Management and Challenges; Briske, D.D., Ed.; Springer Series on Environmental Management; Springer: Cham, Switzerland, 2017; pp. 25–84. [Google Scholar]
- Bond, W.J.; Midgley, G.F. A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Glob. Chang. Biol. 2000, 6, 865–869. [Google Scholar] [CrossRef]
- Leite, P.A.; Wilcox, B.P.; McInnes, K.J. Woody plant encroachment enhances soil infiltrability of a Semiarid Savanna. Environ. Res. Commun. 2020, 2, 115005. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, G.F. Carbon dioxide and the uneasy interactions of trees and savannah grasses. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, J.N.; Morgan, J.W. Woody plant encroachment reduces species richness of herb-rich woodlands in southern Australia. Austral Ecol. 2008, 33, 278–289. [Google Scholar] [CrossRef]
- Archer, S.; Boutton, T.W.; Hibbard, K.A. Trees in Grasslands: Biogeochemical Consequences Woody Plant Expansion. In Global Biogeochemical Cycles in the Climate System; Academic Press: Cambridge, MA, USA, 2001; pp. 115–137. [Google Scholar]
- Liao, J.D.; Boutton, T.W.; Jastrow, J.D. Organic matter turnover in soil physical fractions following woody plant invasion of grassland: Evidence from natural 13C and 15N. Soil Biol. Biochem. 2006, 38, 3197–3210. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub Invasions of North American Semiarid Grasslands. Annu. Rev. Ecol. Syst. 2000, 31, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Leis, S.A.; Blocksome, C.E.; Twidwell, D.; Fuhlendorf, S.D.; Briggs, J.M.; Sanders, L.D. Juniper Invasions in Grasslands: Research Needs and Intervention Strategies. Rangelands 2017, 39, 64–72. [Google Scholar] [CrossRef]
- Chaneton, E.J.; Mazia, N.; Batista, W.B.; Rolhauser, A.G.; Ghersa, C.M. Woody Plant Invasions in Pampa Grasslands: A Biogeographical and Community Assembly Perspective. In Ecotones Between Forest and Grassland; Springer: New York, NY, USA, 2012; pp. 115–144. ISBN 9781461437970. [Google Scholar]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, D.J.; Soliveres, S.; Bowker, M.A.; Val, J. Grazing dampens the positive effects of shrub encroachment on ecosystem functions in a semi-arid woodland. J. Appl. Ecol. 2013, 50, 1028–1038. [Google Scholar] [CrossRef]
- Zhou, D.; Xia, Z.; Dong, L.; Wenjiang, H.; Dailiang, P.; Linsheng, H. Remote sensing identification of shrub encroachment in grassland in Inner Mongolia. Transations Chin. Soc. Agric. Eng. 2014, 30, 152–158. [Google Scholar]
- Sanjuán, Y.; Arnáez, J.; Beguería, S.; Lana-Renault, N.; Lasanta, T.; Gómez-Villar, A.; Álvarez-Martínez, J.; Coba-Pérez, P.; García-Ruiz, J.M. Woody plant encroachment following grazing abandonment in the subalpine belt: A case study in northern Spain. Reg. Environ. Chang. 2018, 18, 1103–1115. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Hik, D.S.; Kennedy, C.; Cooley, D.; Johnstone, J.F.; Kenney, A.J.; Krebs, C.J. Expansion of canopy-forming willows over the twentieth century on Herschel Island, Yukon Territory, Canada. Ambio 2011, 40, 610–623. [Google Scholar] [CrossRef]
- Bailey, A.A.W.; Irving, B.D.; Fitzgerald, R.D.; Journal, S.; May, N. Regeneration of Woody Species following Burning and Grazing in Aspen Parkland. J. Range Manag. 1990, 43, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Hilton, J.E.; Bailey, A.W. Cattle Use of a Sprayed Aspen Parkland Range. J. Range Manag. 1972, 25, 257–260. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Wilson, S.D. Hailstorm damage promotes aspen invasion into grassland. Can. J. Bot. 2006, 84, 1142–1147. [Google Scholar] [CrossRef]
- Guedo, D.D.; Lamb, E.G. Prescribed burning has limited long-term effectiveness in controlling trembling aspen (populus tremuloides) encroachment into fescue grassland in prince albert national park. Can. Field-Nat. 2013, 127, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, A.R.D.; Bailey, A.W. Control of Aspen Regrowth by Grazing with Cattle. J. Range Manag. 1984, 37, 156–158. [Google Scholar] [CrossRef]
- Campbell, C.; Campbell, I.D.; Blyth, C.B.; Mcandrews, J.H. Bison Extirpatio May Have Caused Aspen Expansion in Western Canada. Ecography 1994, 17, 360–362. [Google Scholar] [CrossRef]
- Moss, R.; Gardiner, B.; Bailey, A.; Oliver, G. A Guide to Integrated Brush Management on the Western Canadian Plains; Canada Manitoba Forage Council: Brandon, MB, Canada, 2008. [Google Scholar]
- Dahl, R. Characterizing Thorny Buffaloberry (Shepherdia argentea) Encroachment into the Mixedgrass Prairie in Alberta, Western Canada. Available online: https://rri.ualberta.ca/2018/09/26/characterizing-thorny-buffaloberry-shepherdia-argentea-encroachment-into-the-mixedgrass-prairie-in-alberta-western-canada-regina-dahl-m-sc-2014/ (accessed on 11 December 2020).
- Bai, Y.; Colberg, T.; Romo, J.T.; McConkey, B.; Pennock, D.; Farrell, R. Does expansion of western snowberry enhance ecosystem carbon sequestration and storage in Canadian Prairies? Agric. Ecosyst. Environ. 2009, 134, 269–276. [Google Scholar] [CrossRef]
- Schellberg, J.; Verbruggen, E. Frontiers and perspectives on research strategies in grassland technology. Crop Pasture Sci. 2014, 65, 508–523. [Google Scholar] [CrossRef]
- Becker, R.H.; Zmijewski, K.A.; Crail, T. Seeing the forest for the invasives: Mapping buckthorn in the Oak Openings. Biol. Invasions 2013, 15, 315–326. [Google Scholar] [CrossRef]
- Somers, B.; Asner, G.P. Invasive species mapping in hawaiian rainforests using multi-temporal hyperion spaceborne imaging spectroscopy. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Oldeland, J.; Dorigo, W.; Wesuls, D.; Jürgens, N. Mapping bush encroaching species by seasonal differences in hyperspectral imagery. Remote Sens. 2010, 2, 1416–1438. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.T.; Rima, P.; Einzmann, K.; Immitzer, M.; Atzberger, C.; Eckert, S. Assessing the potential of sentinel-2 and pléiades data for the detection of Prosopis and Vachellia spp. in Kenya. Remote Sens. 2017, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Mirik, M.; Chaudhuri, S.; Surber, B.; Ale, S.; James Ansley, R. Detection of two intermixed invasive woody species using color infrared aerial imagery and the support vector machine classifier. J. Appl. Remote Sens. 2013, 7, 073588. [Google Scholar] [CrossRef] [Green Version]
- Kattenborn, T.; Lopatin, J.; Förster, M.; Braun, A.C.; Fassnacht, F.E. UAV data as alternative to field sampling to map woody invasive species based on combined Sentinel-1 and Sentinel-2 data. Remote Sens. Environ. 2019, 227, 61–73. [Google Scholar] [CrossRef]
- Hantson, W.; Kooistra, L.; Slim, P.A. Mapping invasive woody species in coastal dunes in the Netherlands: A remote sensing approach using LIDAR and high-resolution aerial photographs. Appl. Veg. Sci. 2012, 15, 536–547. [Google Scholar] [CrossRef]
- Mori, N. Composition and Structure of Fescue Prairie Respond to Burning and Environmental Conditions More Than to Grazing or Burning Plus Grazing in the Short Term. Master’s Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2009. [Google Scholar]
- Archibold, O.W.; Ripley, E.A.; Bretell, D.L. Comparison of the Microclimates of a Small Aspen Grove and Adjacent Prairie in Saskatchewan. Am. Midl. Nat. 1996, 136, 248–261. [Google Scholar] [CrossRef]
- Soubry, I.; Guo, X. Identification of the Optimal Season and Spectral Regions for Shrub Cover Estimation in Grasslands. Sensors 2021, 21, 3098. [Google Scholar] [CrossRef]
- Romo, J.T.; Grilz, P.L.; Redmann, R.E.; Driver, E.A. Standing Crop, Biomass Allocation Patterns and Soil-Plant Water Relations in Symphoricarpos occidentalis Hook. Following Autumn or Spring Burning. Am. Midl. Nat. 1993, 130, 106–115. [Google Scholar] [CrossRef]
- Slopek, J.I.; Lamb, E.G. Long-Term Efficacy of Glyphosate for Smooth Brome Control in Native Prairie. Invasive Plant Sci. Manag. 2017, 10, 350–355. [Google Scholar] [CrossRef]
- Baines, G.B.K. Plant Distribution on a Saskatchewan Prairie. Vegetatio 1973, 28, 99–123. [Google Scholar] [CrossRef]
- Statistics Canada 2016 Census-Boundary Files. Available online: https://www12.statcan.gc.ca/census-recensement/2011/geo/bound-limit/bound-limit-2016-eng.cfm (accessed on 20 January 2021).
- ESA Copernicus Open Access Hub. Available online: https://scihub.copernicus.eu/dhus/#/home (accessed on 8 March 2021).
- Scott, H.A. Symphoricarpos occidentalis. Available online: https://www.fs.fed.us/database/feis/plants/shrub/symocc/all.html (accessed on 16 April 2020).
- Manske, L.L. Western Snowberry Biology, 2006 Annual Report, Grassland Section; Dickinson Research Extension Center: Dickinson, ND, USA, 2006. [Google Scholar]
- Schneider, R.E.; Faber-Langendoen, D.; Don Crawford, R.C.; Weakley, A.S. The Status of Biodiversity in the Great Plains: Great Plains Vegetation Classification; Supplemental Document 1; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Missoula, MT, USA, 1997. [Google Scholar]
- Lawrence, D.L.; Romo, J. Tree and shrub communities of woodes draws near the matador research station in Southern Saskatchewan. Can. Field-Nat. 1994, 108, 397–412. [Google Scholar]
- Köchy, M.; Wilson, S.D. Semiarid grassland responses to short-term variation in water availability. Plant Ecol. 2004, 174, 197–203. [Google Scholar] [CrossRef]
- Clarke, S.E.; Tisdale, E.W.; Skoglund, N.A. The Effects of Climate and Grazing Practices on Short-Grass Prairie Vegetation in Southern Alberta and Southwestern Saskatchewan; Technical Bulletin; Dominion of Canada, Department of Agriculture: Ottawa, ON, Canada, 1947. [Google Scholar]
- Hardy BBT Limited. Manual of Plant Species Suitability for Reclamation in Alberta, 2nd ed.; Alberta Land Concervation and Reclamation Council: Edmonton, AB, Canada, 1989. [Google Scholar] [CrossRef]
- Hall, J.B.; Hansen, P.L. A Preliminary Riparian Habitat Type Classification System for the Bureau of Land Management Districts in Southern and Eastern Idaho. Available online: https://archive.org/details/preliminaryripar32hall/mode/2up (accessed on 16 April 2020).
- Hansen, P.L.; Thompson, W.H.; Smith, R.; Yeager, T. Classification and Management of Upland, Riparian, and Wetland Sites of USDI Bureau of Land Management’s Miles City Field Office, Eastern Montana USA. Nat. Resour. Environ. Issues 2011, 16, 32. [Google Scholar]
- Bowes, G.G.; Spurr, D.T. Improved forage production following western snowberry (Symphoricarpos occidentalis Hook control with metsulfuron methyl. Can. J. Plant Sci. 1995, 75, 935–940. [Google Scholar] [CrossRef]
- Pelton, J. Studies on the Life-History of Symphoricarpos occidentalis Hook, in Minnesota. Ecol. Monogr. 1953, 23, 17–39. [Google Scholar] [CrossRef]
- Government of Saskatchewan. Managing Saskatchewan Rangeland, 1st ed.; Bruynooghe, J., Macdonald, R., Eds.; Agriculture and Agri-Food Canada: Regina, SK, Canada, 2008. [Google Scholar]
- Lackschewitz, K. Vascular Plants of West-Central Montana-Identification Guidebook; US Department of Agriculture, Forest Service: Ogden, UT, USA, 1991. [Google Scholar]
- Esser, L.L. Elaeagnus commutata. Available online: https://www.fs.fed.us/database/feis/plants/shrub/elacom/all.html (accessed on 16 April 2020).
- Nesom, G. American Silverberry—Elaeagnus commutata Bernh. ex Rydb.; US Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 1998. [Google Scholar]
- Arnold, T.W.; Higgins, K.F. Effects of shrub coverages on birds of North Dakota mixed-grass prairies. Can. Field-Nat. 1986, 100, 10–14. [Google Scholar]
- Pylypec, B. Impacts of fire on bird populations in a fescue prairie. Can. Field-Nat. 1991, 105, 346–349. [Google Scholar]
- Bailey, A.W. Barrier Effect of the Shrub Elaeagnus commutata on Grazing Cattle and Forage Production in Central Alberta. J. Range Manag. 1970, 23, 248–251. [Google Scholar] [CrossRef]
- Rowe, J.S. Uses of Undergrowth Plant Species in Forestry Author. Ecology 1956, 37, 461–473. [Google Scholar] [CrossRef]
- Corns, W.G.; Schraa, R.J. Mechanical and Chemical Control of Silverberry (Elaeagnus commutata Bernh.) on Native Grassland. J. Range Manag. 1965, 18, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S.; Murphy, M.A.; Ram, K. Package ‘spatialEco’—Spatial Analysis and Modelling Utilities; Version 1.3-5; CRAN. Available online: https://github.com/jeffreyevans/spatialEco (accessed on 20 February 2021).
- Kaufman, Y.J.; Remer, L.A. Detection of Forests Using Mid-IR Reflectance: An Application for Aerosol Studies. IEEE Trans. Geosci. Remote Sens. 1994, 32, 672–683. [Google Scholar] [CrossRef]
- Bhattacharyya, A. On a Measure of Divergence between Two Multinomial Populations. Indian J. Stat. 1946, 7, 401–406. [Google Scholar]
- Bruzzone, L.; Roli, F.; Serpico, S.B. An Extension of the Jeffreys-Matusita Distance to Multiclass Cases for Feature Selection. IEEE Trans. Geosci. Remote Sens. 1995, 33, 1318–1321. [Google Scholar] [CrossRef] [Green Version]
- Jeffreys, H. Theory of Probability, 2nd ed.; Clarendon Press: Oxford, UK, 1948. [Google Scholar]
- Davis, S.M.; Landgrebe, D.A.; Phillips, T.L.; Swain, P.H.; Hoffer, R.M.; Lindenlaub, J.C.; Silva, L.F. Remote Sensing: The Quantitative Approach; Swain, P.H., Davis, S.M., Eds.; McGraw-Hill International Book Co.: New York, NY, USA, 1978; ISBN 007062576X. [Google Scholar]
- Gunal, S.; Edizkan, R. Subspace based feature selection for pattern recognition. Inf. Sci. 2008, 178, 3716–3726. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Mohd Razali, N.; Bee Wah, Y. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]
- Huang, H.; Roy, D.P.; Boschetti, L.; Zhang, H.K.; Yan, L.; Kumar, S.S.; Gomez-Dans, J.; Li, J. Separability analysis of Sentinel-2A Multi-Spectral Instrument (MSI) data for burned area discrimination. Remote Sens. 2016, 8, 873. [Google Scholar] [CrossRef] [Green Version]
- Kailath, T. The Divergence and Bhattacharyya Distance Measures in Signal Selection. IEEE Trans. Commun. Technol. 1967, 15, 52–60. [Google Scholar] [CrossRef]
- Campbell, J.E.; Harris, J.R.; Huntley, D.H.; McMartin, I.; Wityk, U.; Dredge, L.A.; Eagles, S. Remote Predictive Mapping of Surficial Earth Materials: Wager Bay North Area, OPEN FILE 7118 Remote Predictive Mapping of Surficial Earth Materials: Wager Bay North Area, Nunavut—NTS 46-E (N), 46-K (SW), 46-L, 46-M (SW), 56-H (N), 56-I and 56-J (S); Geological Survey of Canada: Ottawa, ON, Canada, 2013. [Google Scholar] [CrossRef]
- Bindel, M.; Hese, S.; Berger, C.; Schmullius, C. Feature selection from high resolution remote sensing data for biotope mapping. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2012, 38, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, L.W.; Meyer, H.; Obermeier, W.A.; Silva, B.; Regeling, B.; Thies, B.; Bendix, J. Hyperspectral data analysis in R: The hsdar package. J. Stat. Softw. 2019, 89. [Google Scholar] [CrossRef] [Green Version]
- Tesfamichael, S.G.; Newete, S.W.; Adam, E.; Dubula, B. Field spectroradiometer and simulated multispectral bands for discriminating invasive species from morphologically similar cohabitant plants. GIScience Remote Sens. 2018, 55, 417–436. [Google Scholar] [CrossRef]
- Goslee, S.C.; Havstad, K.M.; Peters, D.P.C.; Rango, A.; Schlesinger, W.H. High-resolution images reveal rate and pattern of shrub encroachment over six decades in New Mexico, USA. J. Arid Environ. 2003, 54, 755–767. [Google Scholar] [CrossRef]
- Cao, X.; Liu, Y.; Cui, X.; Chen, J.; Chen, X. Mechanisms, monitoring and modeling of shrub encroachment into grassland: A review. Int. J. Digit. Earth 2019, 12, 625–641. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Separability Statistic | Threshold Value | Separability Class |
|---|---|---|
| M-Statistic | >1 | Good |
| ≤1 | Poor | |
| Transformed Divergence & Jeffries–Matusita Distance | ≥1.8 | Good |
| 1.51–1.79 | Moderate | |
| ≤1.5 | Poor |
| Separability between Western Snowberry and Wolfwillow | ||||
|---|---|---|---|---|
| Season | Wavelength Areas | |||
| Moderate | Good | |||
| (nm) | Category | (nm) | Category | |
| Spring | / | / | 409–525 | B |
| / | / | 590–693 | R/RE | |
| Summer | 532–577 | G | 406–531 | B |
| 1981–1991 | SWIR | 578–692 | R/RE | |
| Fall | / | / | / | / |
| Two-Sample t-Test p-Values | Yel. | <0.05 | Red | <0.01 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Landsat-8 | Sentinel-2A | |||||||||||||||
| B | G | R | NIR | SWIR 1 | SWIR 2 | B | G | R | RE 1 | RE 2 | RE 3 | NIR | RE 4 | W. Vap. | SWIR 1 | SWIR 2 | |
| Spring | 0.000 | 0.000 | 0.000 | 0.049 | 0.748 | 0.001 | 0.000 | 0.000 | 0.000 | 0.006 | 0.046 | 0.051 | 0.049 | 0.052 | 0.027 | 0.702 | 0.001 |
| Summer | 0.000 | 0.000 | 0.000 | 0.000 | 0.005 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.006 | 0.000 |
| Fall | 0.000 | 0.035 | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.047 | 0.000 | 0.077 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 | 0.000 |
| Separability between Western Snowberry and Wolfwillow | ||||
|---|---|---|---|---|
| Season | Wavelength Bands | |||
| Moderate | Good | |||
| Landsat 8 | Sentinel-2A | Landsat 8 | Sentinel-2A | |
| Spring | B | / | / | / |
| R | ||||
| Summer | G | G | B | B |
| R | R | |||
| Fall | / | / | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soubry, I.; Guo, X. Seasonal Spectral Separation of Western Snowberry and Wolfwillow in Grasslands with Field Spectroradiometer and Simulated Multispectral Bands. Environments 2021, 8, 60. https://doi.org/10.3390/environments8070060
Soubry I, Guo X. Seasonal Spectral Separation of Western Snowberry and Wolfwillow in Grasslands with Field Spectroradiometer and Simulated Multispectral Bands. Environments. 2021; 8(7):60. https://doi.org/10.3390/environments8070060
Chicago/Turabian StyleSoubry, Irini, and Xulin Guo. 2021. "Seasonal Spectral Separation of Western Snowberry and Wolfwillow in Grasslands with Field Spectroradiometer and Simulated Multispectral Bands" Environments 8, no. 7: 60. https://doi.org/10.3390/environments8070060
APA StyleSoubry, I., & Guo, X. (2021). Seasonal Spectral Separation of Western Snowberry and Wolfwillow in Grasslands with Field Spectroradiometer and Simulated Multispectral Bands. Environments, 8(7), 60. https://doi.org/10.3390/environments8070060