
Case Study Comparing Effects of Microplastic Derived from Bottle Caps Collected in Two Cities on Triticum aestivum (Wheat)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microplastic
2.2. Wheat Plant Cultivation
2.3. Exposure Scenarios
2.4. Measurement of Root and Shoot Length
2.5. Oxidative Stress Status
2.5.1. Measurement of Lipid Peroxidation
2.5.2. Measurement of Catalase Activity
2.6. Statistical Tests
3. Results and Discussion
3.1. Length of Root and Shoot
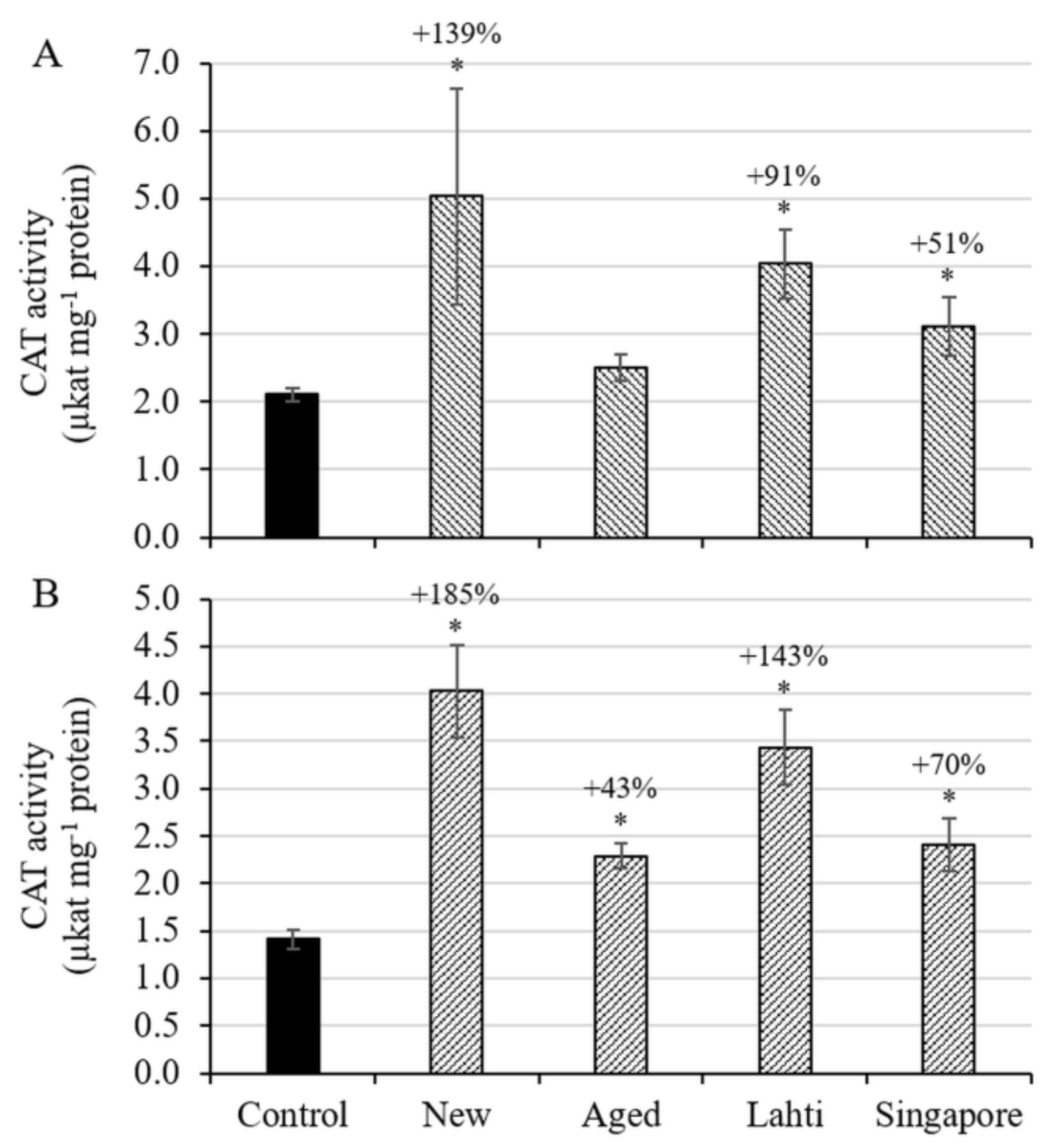
3.2. Oxidative Stress Status
3.3. Potential Relationship between MP Toxicity and Environmental Factors
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vethaak, A.D.; Legler, J. Microplastics and human health. Science 2021, 12, 672–674. [Google Scholar] [CrossRef]
- Arthur, C.; Baker, J.; Bamford, H. Proceedings of the International Research Workshop on the Occurrence, Effects, and Fate of Microplastic Marine Debris, Tacoma, WA, USA, 9–11 September 2008; NOAA Marine Debris Division: Silver Spring, MD, USA, 2008; p. 530. [Google Scholar]
- Dris, R.; Gasperi, J.; Rocher, V.; Saad, M.; Renault, N.; Tassin, B. Microplastic contamination in an urban area: A case study in Greater Paris. Environ. Chem. 2015, 12, 592–599. [Google Scholar] [CrossRef]
- Allen, S.; Allen, D.; Phoenix, V.R.; Le Roux, G.; Jiménez, P.D.; Simonneau, A.; Binet, S.; Galop, D. Atmospheric transport and deposition of microplastics in a remote mountain catchment. Nat. Geosci. 2019, 12, 339–344. [Google Scholar] [CrossRef]
- Scopetani, C.; Chelazzi, D.; Cincinelli, A.; Esterhuizen-Londt, M. Assessment of microplastic pollution: Occurrence and characterisation in Vesijärvi lake and Pikku Vesijärvi pond, Finland. Environ. Monit. Assess. 2019, 191, 652. [Google Scholar] [CrossRef] [Green Version]
- Kulshreshtha, A.K. Chemical degradation. In Handbook of Polymer Degradation; Hamid, S.H., Amin, M.B., Maadhah, A.G., Eds.; Taylor & Francis: Oxford, UK, 1992; pp. 55–94. [Google Scholar]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: A comprehensive review. Biotech. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.R. Cause and effect in the weathering of plastics. Polym. Eng. Sci. 1970, 10, 108–121. [Google Scholar] [CrossRef]
- Davis, A.; Sims, D. Weathering of Polymers; Springer Science & Business Media: London, UK, 1983. [Google Scholar]
- Rabinovitch, E.; Queensbury, G.J.; Summers, J.W. Predicting heat build-up due to the sun’s energy. J. Vinyl. Addit. Technol. 1983, 5, 110–115. [Google Scholar] [CrossRef]
- Andrady, A.L. Proceedings of Symposium on Degradable Plastics, Washington, DC, USA, 10 June 1987; Society of Plastics Industry: Washington, DC, USA, 1987; pp. 22–25. [Google Scholar]
- Fischer, C.E.; Castelli, V.J.; Rodgers, S.D.; Bleile, H.R. Technology for control of marine biofouling. In Marine Biodeterioration: An Interdisciplinary Study; Costlow, J.D., Tipper, R.C., Eds.; Naval Institute Press: Annapolis, MD, USA, 1984; pp. 261–300. [Google Scholar]
- Cooper, D.A.; Corcoran, P.L. Effects of mechanical and chemical processes on the degradation of plastic beach debris on the island of Kauai, Hawaii. Mar. Pollut. Bull. 2010, 60, 650–654. [Google Scholar] [CrossRef]
- Köppen, W. Die Wärmezonen der Erde, nach der Dauer der heissen, gemässigten und kalten Zeit und nach der Wirkung der Wärme auf die organische Welt betrachtet (The thermal zones of the earth according to the duration of hot, moderate and cold periods and to the impact of heat on the organic world). Meteorol. Z 1884, 1, 215–226, Translated and edited by Volken, E., Ronnimänn, S.B. Meteorol. Z. 2011, 20, 351–360. [Google Scholar]
- Geiger, R. Klassifikation der Klimate Nach W. Köppen (Classification of Climates after W. Köppen). Landolt-Börnstein—Zahlenwerte und Funktionen aus Physik, Chemie, Astronomie, Geophysik und Technik, Alte Serie 3; Springer: Berlin/Heidelberg, Germany, 1954; pp. 603–607. [Google Scholar]
- Arutchelvi, J.; Sudhakar, M.; Arkatkar, A.; Doble, M.; Bhaduri, S.; Uppara, P.V. Biodegradation of polyethylene and polypropylene. Indian J. Biotech. 2008, 7, 9–22. [Google Scholar]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Luo, Y.; Lu, S.; Liu, M.; Song, Y.; Lei, L. Microplastics in soils: Analytical methods, pollution characteristics and ecological risks. TrAC Trends Anal. Chem. 2018, 109, 163–172. [Google Scholar] [CrossRef]
- Fejdyś, M.; Landwijt, M.; Strusczyk, M.H. Effects of accelerated ageing conditions on the degradation process of dyneema ® polyethylene composites. Fibres Text East Eur. 2011, 19, 60–65. [Google Scholar]
- PN-EN 12280-1:2002. Rubber- or Plastics-Coated Fabrics—Accelerated Ageing Tests—Part 1: Heat Ageing; EN 12280-1:1997; International Organization for Standards: Geneva, Switzerland, 1997. [Google Scholar]
- Walters, D.R.; Kingham, G. Biosynthesis inhibitors by barley seedlings: Effects of mildew infections. New Phytol. 1990, 114, 659–665. [Google Scholar] [CrossRef]
- Fuller, S.; Gautam, A.A. Procedure for measuring microplastics using pressurised fluid extraction. Environ. Sci. Technol. 2016, 50, 5774–5780. [Google Scholar] [CrossRef] [Green Version]
- Scopetani, C.; Esterhuizen-Londt, M.; Chelazzi, D.; Cincinelli, A.; Setälä, H.; Pflugmacher, S. Self-contamination from clothing in microplastics research. Ecotoxicol. Environ. Saf. 2020, 189, 110036. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, sensitive, and specific thiobarbituric acid method for measuring lipid peroxidation in animal tissue, food, and feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Stüven, J.; Pflugmacher, S. Antioxidative stress response of Lepidium sativum due to exposure to cyanobacterial secondary metabolites. Toxicon 2007, 50, 85–93. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Sokal, R.R.; Rohlf, F.J. Biometry. In The Principles and Practice of Statistic in Biological Research; WH Freeman and Company: New York, NY, USA, 1997. [Google Scholar]
- Atta, B.M.; Mahmood, T.; Trethowan, R.M. Relationship between root morphology and grain yield of wheat in North-Western NSW, Australia. Aust. J. Crop. Sci. 2013, 7, 2108–2115. [Google Scholar] [CrossRef]
- Walter, A.; Schurr, U. Dynamics of leaf and root growth: Endogenous control versus environmental impact. Ann. Bot. 2005, 95, 900–981. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimise water and N acquisition by maise root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, M.; Palta, J.A.; Fillery, I.R.P. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust. J. Agr. Res. 2006, 57, 1097–1107. [Google Scholar] [CrossRef]
- Palta, J.A.; Fillery, I.R.P.; Rebetzke, G.J. Restricted-tillering wheat does not lead to greater investment in roots and early nitrogen uptake. Field Crop Res. 2007, 104, 52–59. [Google Scholar] [CrossRef]
- Pflugmacher, S. Promotion of oxidative stress in the aquatic macrophyte Ceratophyllum demersum during biotransformation of the cyanobacterial toxin microcystin-LR. Aquat. Toxicol. 2004, 70, 169–178. [Google Scholar] [CrossRef]
- Manikowski, H.; Jurga, J. Free radicals in plastic food containers. Mater. Sci. 2003, 9, 206–208. [Google Scholar]
- Fotopoulou, K.; Karapanagioti, H. Degradation of various plastics in the environment. In Hazardous Chemicals Associated with Plastics in the Marine Environment; Takada, H., Karapanagioti, H.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T.G. The effect of β-N-methylamino-L-alanine (BMAA) on oxidative stress response enzymes of the macrophyte Ceratophyllum demersum. Toxicon 2011, 57, 803–810. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Tallinen, S.; Kim, Y.J.; Kim, S.; Esterhuizen, M. Ageing affects microplastic toxicity over time: Effects of aged polycarbonate on germination, growth, and oxidative stress of Lepidium sativum. Sci. Total Environ. 2021, 790, 148166. [Google Scholar] [CrossRef]
- Yu, P.; Liuz, Z.; Wu, D.; Chen, M.; Lv, W.; Zhao, Y. Accumulation of polystyrene microplastics in juvenile Eriocheir sinensis and oxidative stress effects in the liver. Aquat. Toxicol. 2018, 200, 28–36. [Google Scholar] [CrossRef]
- Oliveira, P.; Barboza, L.G.A.; Branco, V.; Figueiredo, N.; Carvalho, C.; Guilhermino, L. Effects of microplastics and mercury in the freshwater bivalve Corbicula fluminea (Müller, 1774): Filtration rate, biochemical biomarkers and mercury bioconcentration. Ecotox. Environ. Saf. 2018, 164, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Carvalho, C.; Guilhermino, L. Microplastics increase mercury bioconcentration in gills and bioaccumulation in the liver, and cause oxidative stress and damage in Dicentrarchus labrax juveniles. Sci. Rep. 2018, 8, 15655. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCFTIR1-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.A.D.S.; Lau, D.W.; Kloas, W.; Bergmann, J.; Bachelier, J.B.; Faltin, E.; Becker, R.; Görlich, A.S.; Rillig, M.C. Microplastics can change soil properties and affect plant performance. Environ. Sci. Technol. 2019, 53, 6044–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflugmacher, S.; Huttunen, J.; van Wolf, M.; Kim, Y.; Kim, S.; Mitrovic, S.; Esterhuizen-Londt, M. Enchytraeus crypticus avoid soil spiked with microplastic. Toxics 2020, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, A.; Leifheit, E.F.; Feng, L.; Bergmann, J.; Wulf, A.; Rillig, M.C. Microplastic fiber and drought effects on plants and soil are only slightly modified by arbuscular mycorrhizal fungi. Soil Ecol. Lett. 2020, 1–13. [Google Scholar] [CrossRef]
- Rushton, B.M.; Salomons, N.S. An investigation of factors relating to the exudation of polyester plasticisers from poly(vinyl) chloride. J. Appl. Polym. Sci. 1969, 13, 2341–2358. [Google Scholar] [CrossRef]
- Madorsky, S.L. Thermal Degradation of Polymers; R.E. Kreiger: New York, NY, USA, 1975. [Google Scholar]
- Anonymous. Thermal Degradation of Plastics; Zeus Industrial Products, Inc: Orangeburg, SC, USA, 2005; pp. 1–8. [Google Scholar]
- Pospíšil, J.; Nešpůrek, S. Highlights in chemistry and physics of polymer stabilisation. Macromol Symp. 1997, 115, 143–163. [Google Scholar] [CrossRef]
- Scalenghe, R. Resource or waste? A perspective of plastics degradation in soil with a focus on end-of-life options. Heliyon 2018, 4, e00941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflugmacher, S.; Sulek, A.; Mader, H.; Heo, J.; Noh, J.H.; Penttinen, O.-P.; Kim, Y.; Kim, S.; Esterhuizen, M. The influence of new and artificial aged microplastic and leachates on the germination of Lepidium sativum L. Plants 2020, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Singapore | Lahti | |
|---|---|---|
| Ave temperature [°C] | 26.7 min 25.8 (January) max 27.4 (May) | 5.2 min −6.3 (January) max 18 (July) |
| Ave temperature variation [°C] | 1.3 | 24.3 |
| Sunshine [Ave h yr−1] | 2064 | 1860 |
| UV-Index | 7–9 | 1–5 |
| Precipitation [mm yr−1] | 2366 | 691 |
| Acidity [pH] | 4.2 | 5.5 |
| Rain [Days yr−1] | 102 | 206 |
| Humidity [%] | 84 | 80 |
| Köppen-Geiger classification | Af tropical rainforest climate | Dfb continental climate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pflugmacher, S.; Tallinen, S.; Mitrovic, S.M.; Penttinen, O.-P.; Kim, Y.-J.; Kim, S.; Esterhuizen, M. Case Study Comparing Effects of Microplastic Derived from Bottle Caps Collected in Two Cities on Triticum aestivum (Wheat). Environments 2021, 8, 64. https://doi.org/10.3390/environments8070064
Pflugmacher S, Tallinen S, Mitrovic SM, Penttinen O-P, Kim Y-J, Kim S, Esterhuizen M. Case Study Comparing Effects of Microplastic Derived from Bottle Caps Collected in Two Cities on Triticum aestivum (Wheat). Environments. 2021; 8(7):64. https://doi.org/10.3390/environments8070064
Chicago/Turabian StylePflugmacher, Stephan, Saila Tallinen, Simon M. Mitrovic, Olli-Pekka Penttinen, Young-Jun Kim, Sanghun Kim, and Maranda Esterhuizen. 2021. "Case Study Comparing Effects of Microplastic Derived from Bottle Caps Collected in Two Cities on Triticum aestivum (Wheat)" Environments 8, no. 7: 64. https://doi.org/10.3390/environments8070064
APA StylePflugmacher, S., Tallinen, S., Mitrovic, S. M., Penttinen, O.-P., Kim, Y.-J., Kim, S., & Esterhuizen, M. (2021). Case Study Comparing Effects of Microplastic Derived from Bottle Caps Collected in Two Cities on Triticum aestivum (Wheat). Environments, 8(7), 64. https://doi.org/10.3390/environments8070064