



Spontaneous Plants Improve the Inter-Row Soil Fertility in a Citrus Orchard but Nitrogen Lacks to Boost Organic Carbon


Abstract
:1. Introduction
2. Materials and Methods
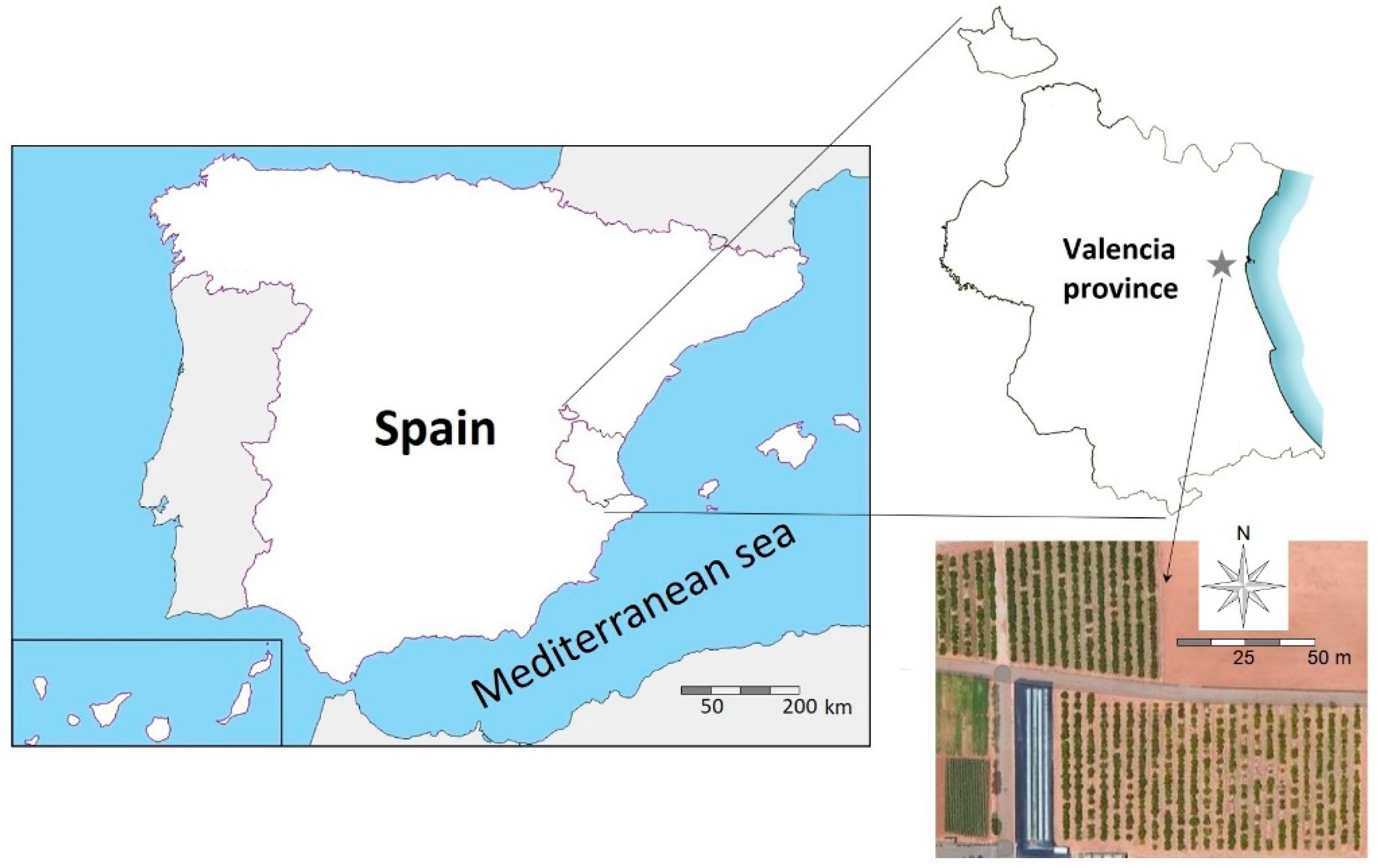
2.1. Study Site
2.2. Crop and Irrigation Characteristics
2.3. Treatments and Trial Plots
2.4. Weather Measurements
2.5. Soil Measurements and Samplings
2.6. Soil Temperature and Water Content Measurements
2.7. Soil Saturated Hydraulic Conductivity
2.8. Soil Carbon Dioxide Emission Rate
2.9. Undisturbed Soil Sampling for Bulk Density
2.10. Undisturbed Soil Sampling for Roots
2.11. Disturbed Soil Sampling
2.12. Statistical Analyses
3. Results
3.1. General Observations
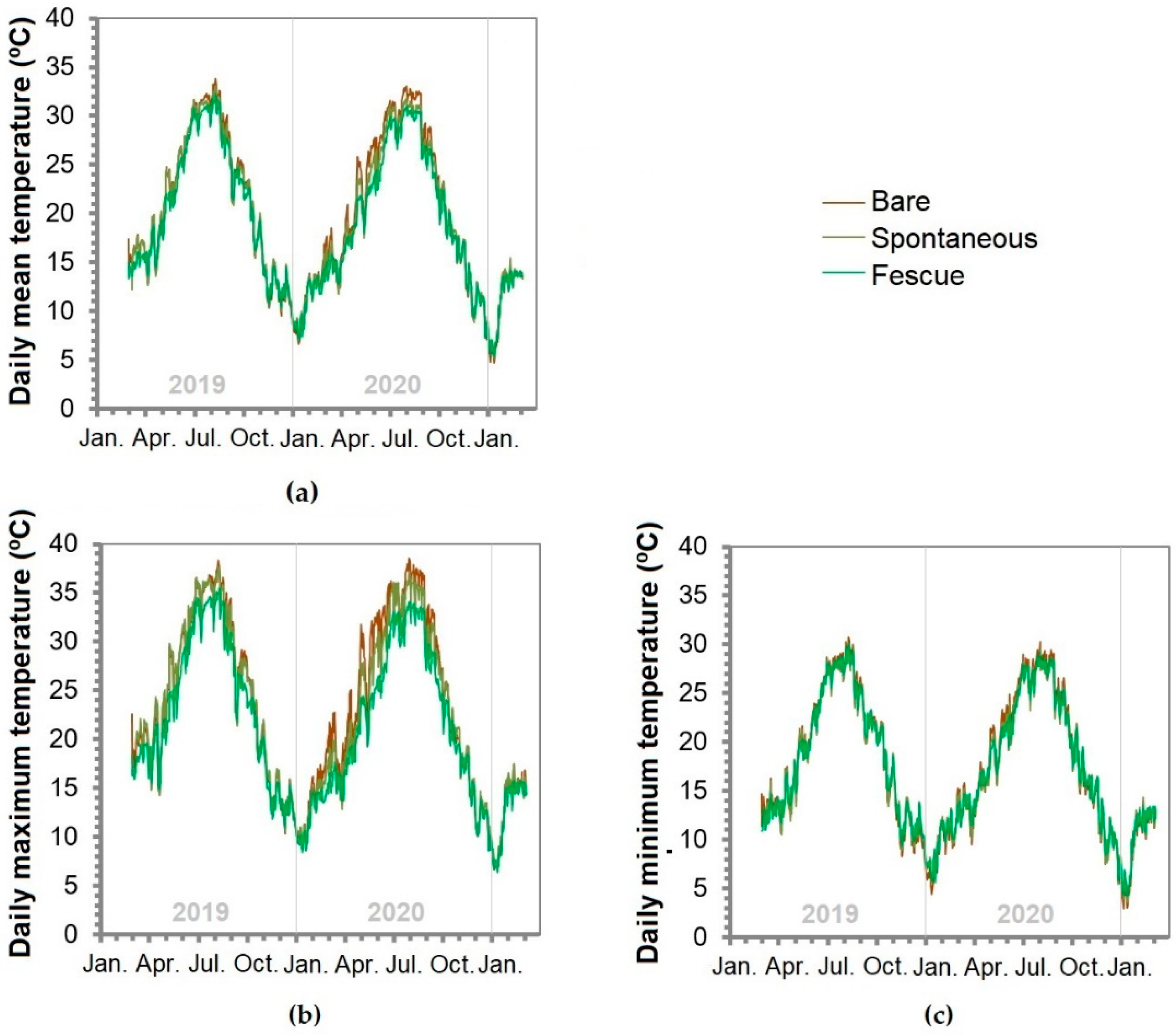
3.2. Soil Temperature
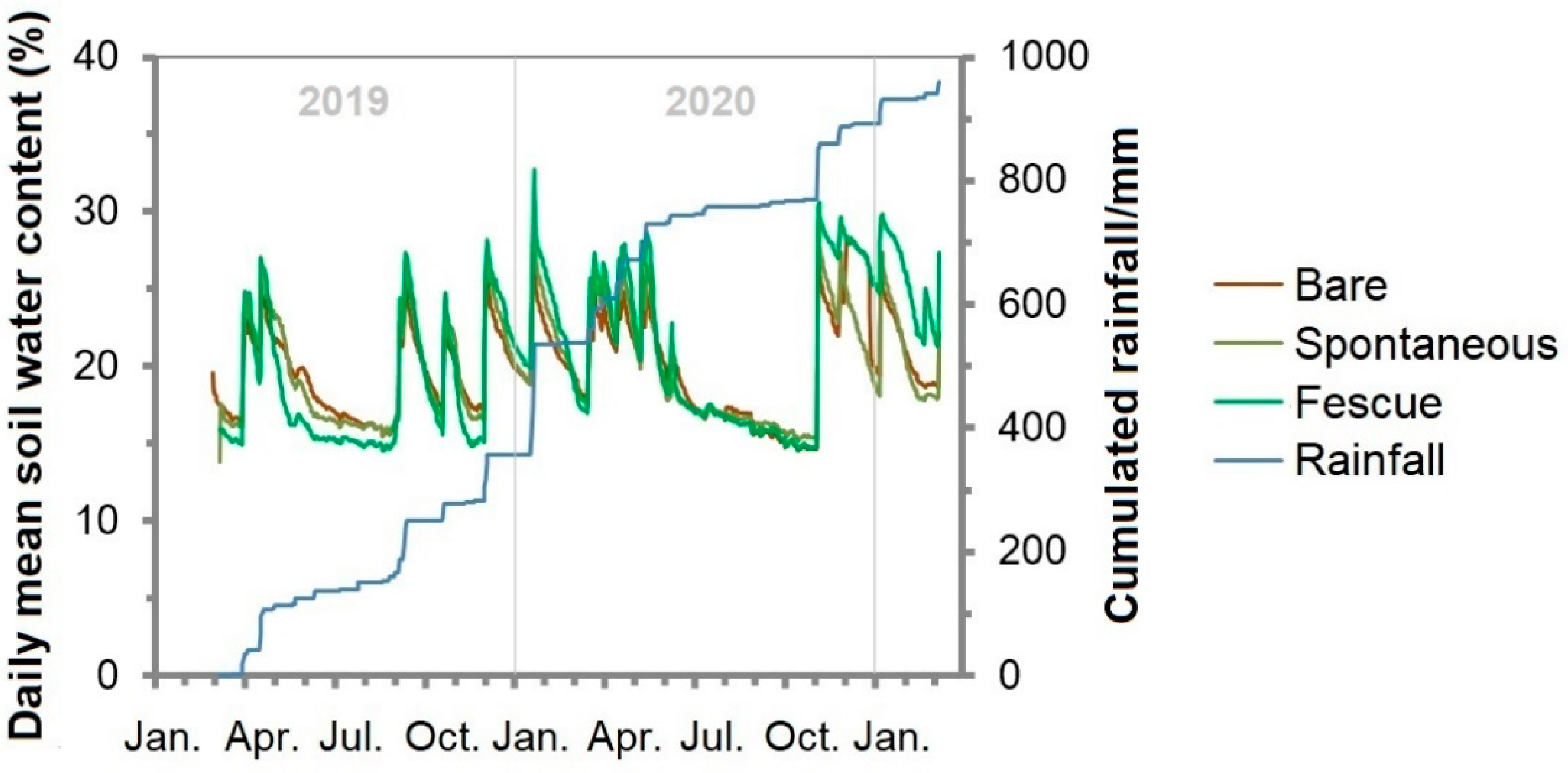
3.3. Soil Water Content
3.4. Statistical Characteristics of the Soil Properties
3.5. Associations among the Soil Properties
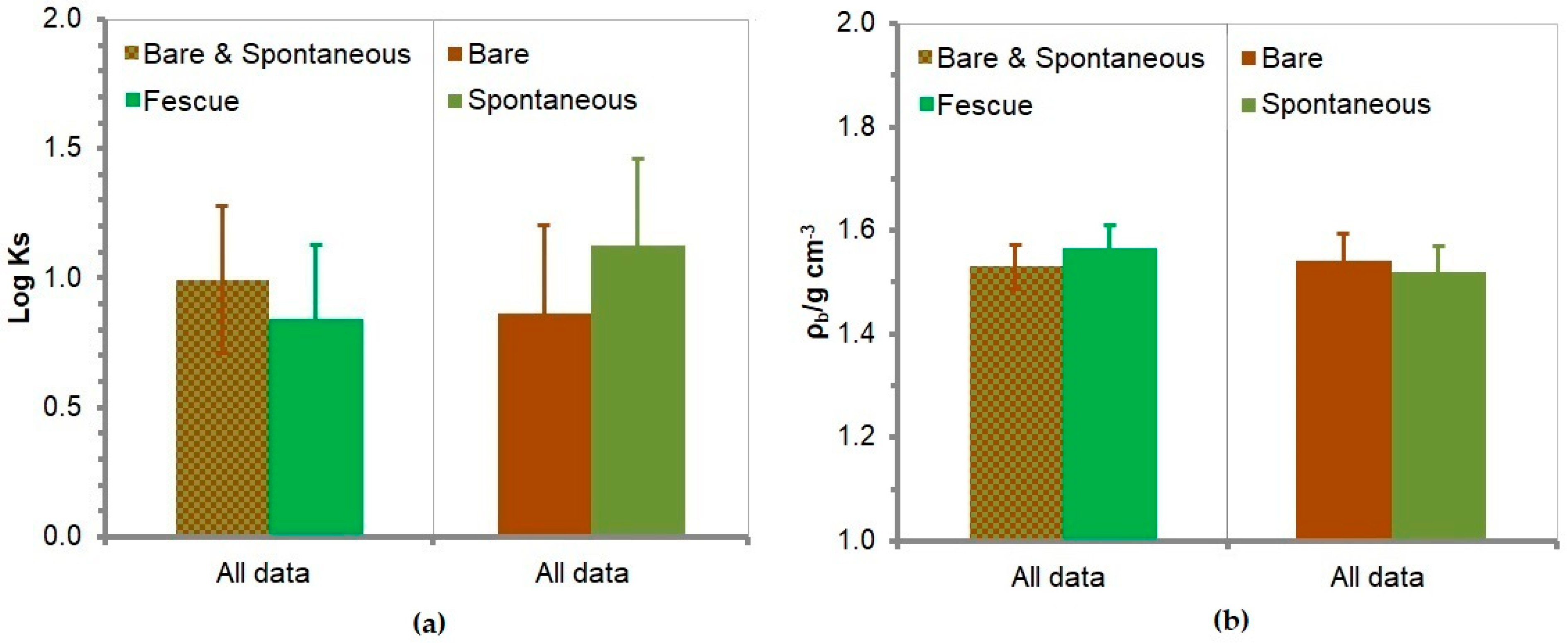
3.6. Soil Saturated Hydraulic Conductivity
3.7. Soil Bulk Density
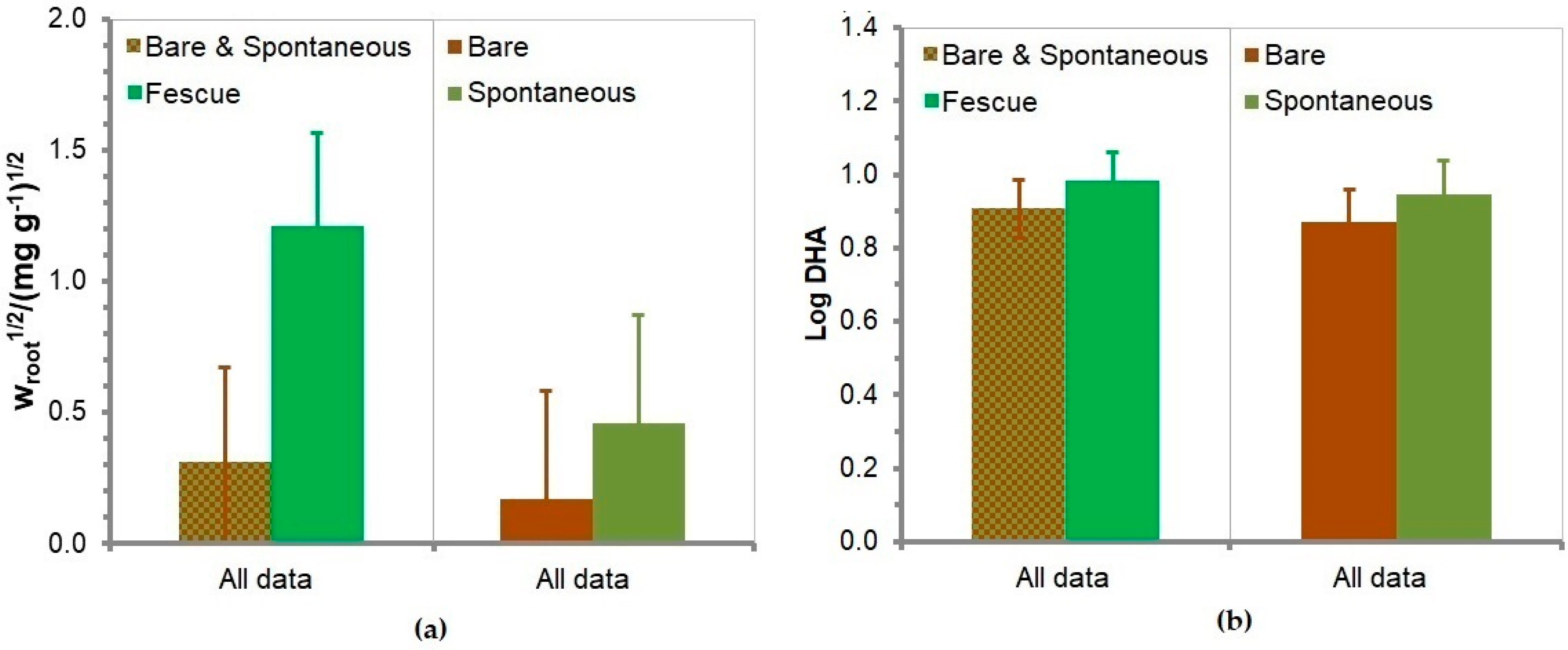
3.8. Soil Root Mass Fraction
3.9. Soil Dehydrogenase Activity
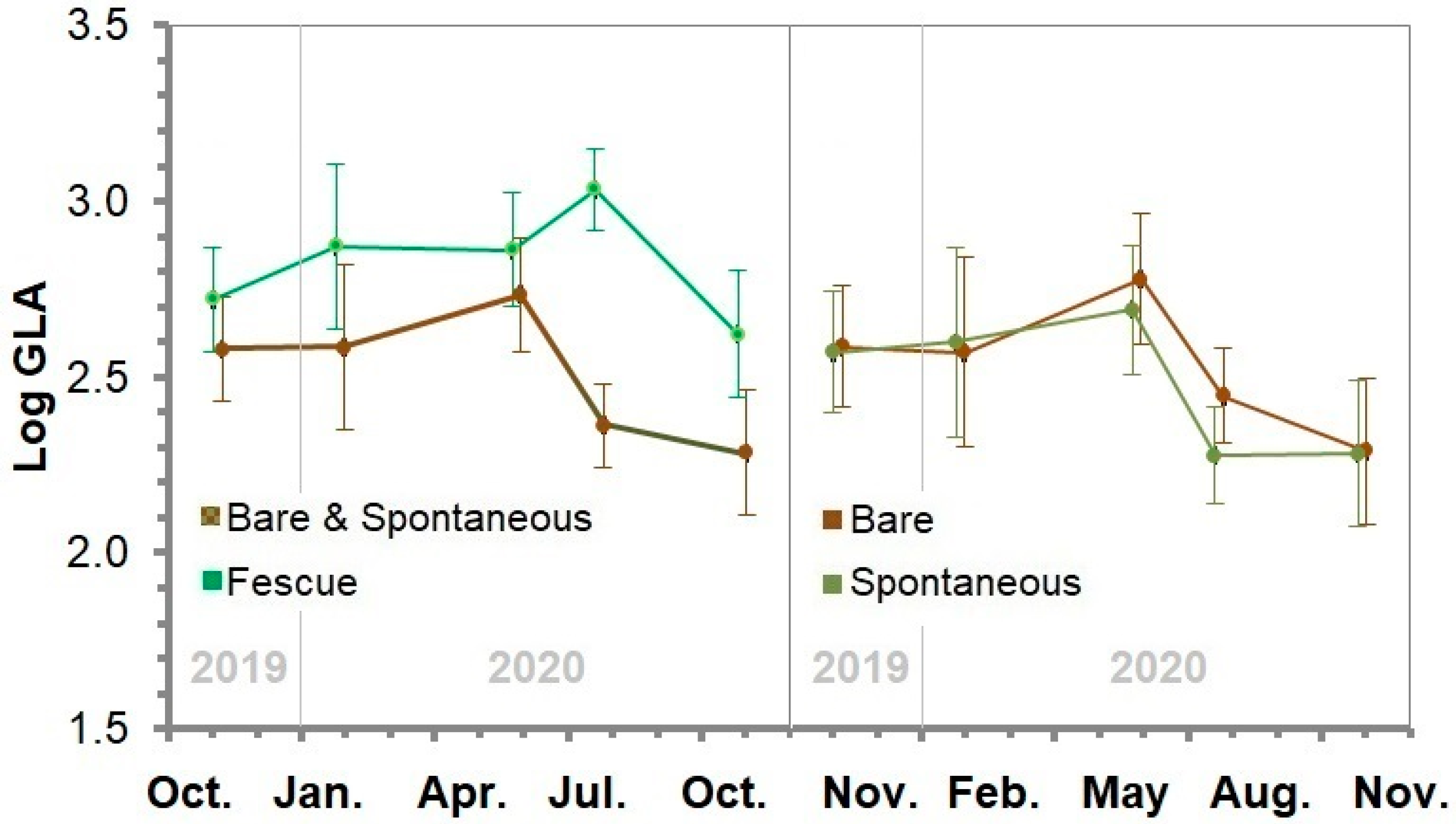
3.10. Soil β-D-Glucosidase Activity
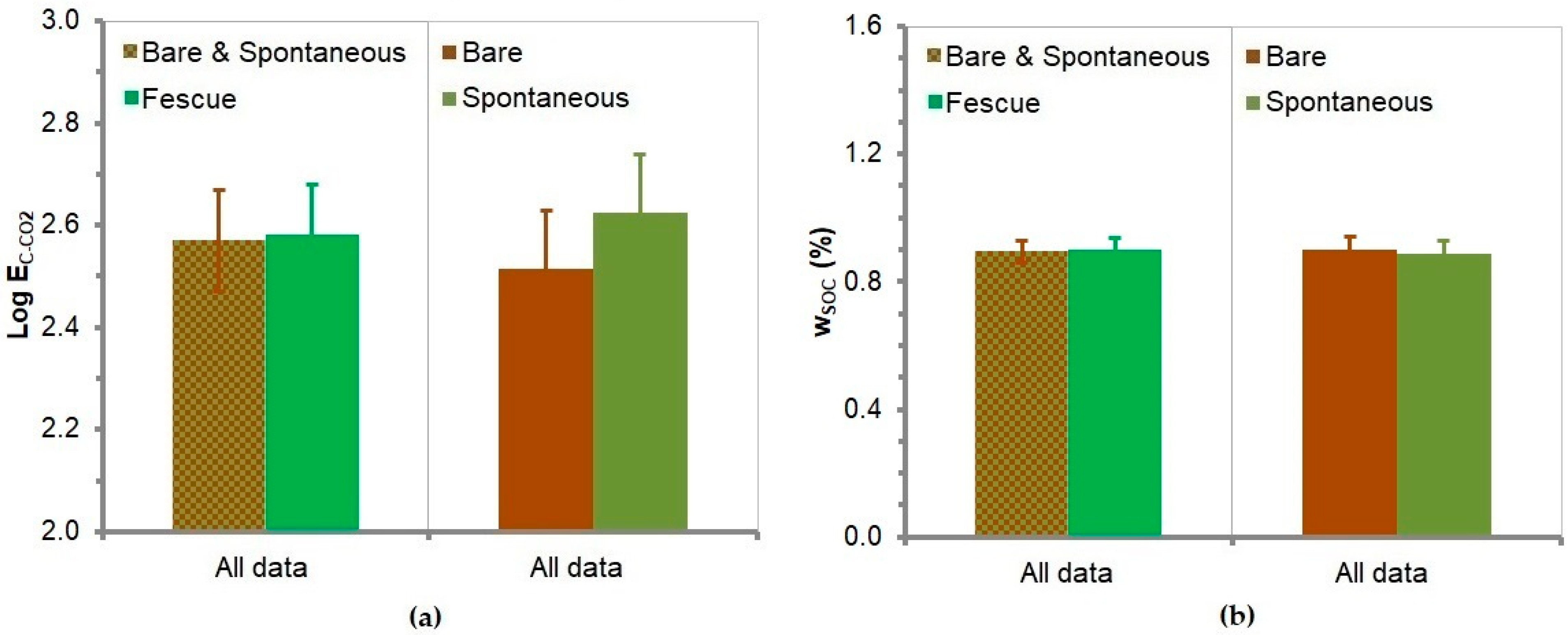
3.11. Soil Carbon Dioxide Emission
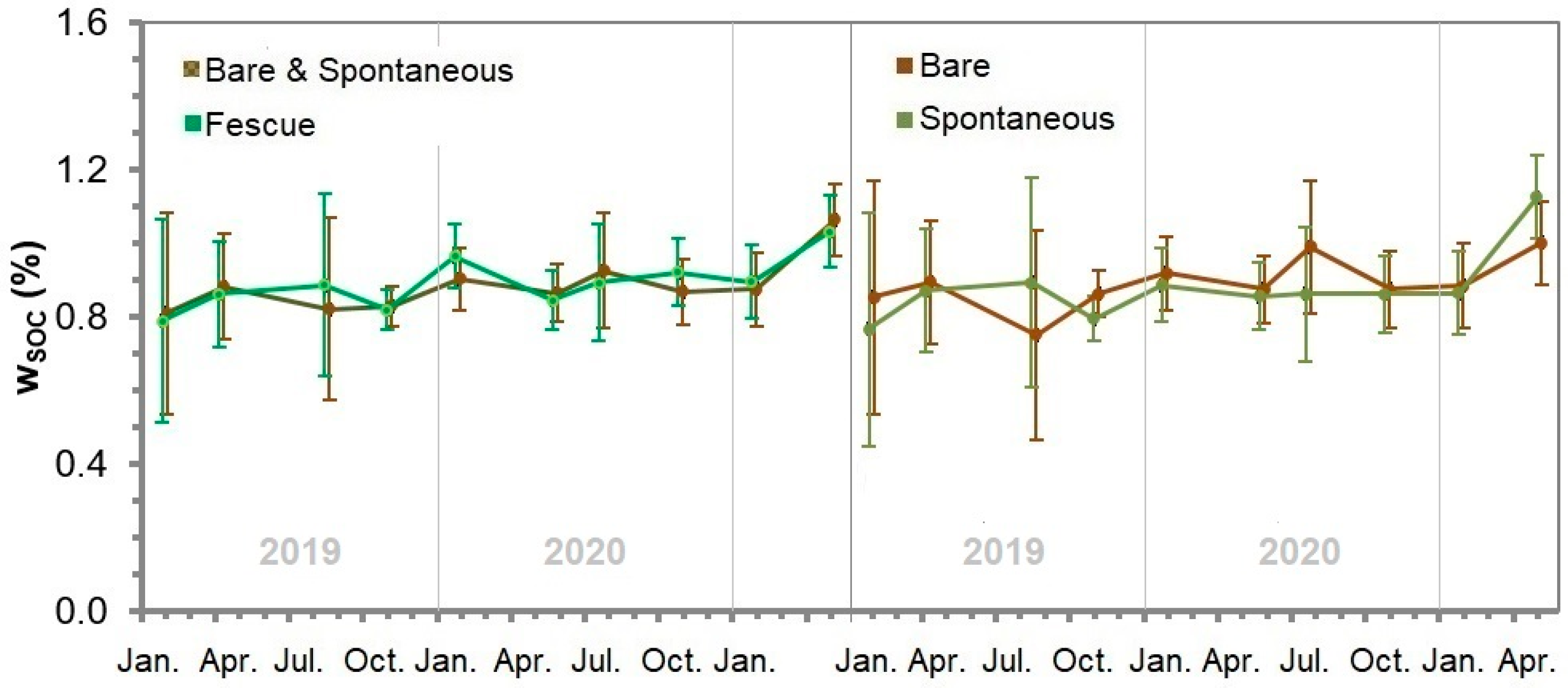
3.12. Soil Organic Carbon
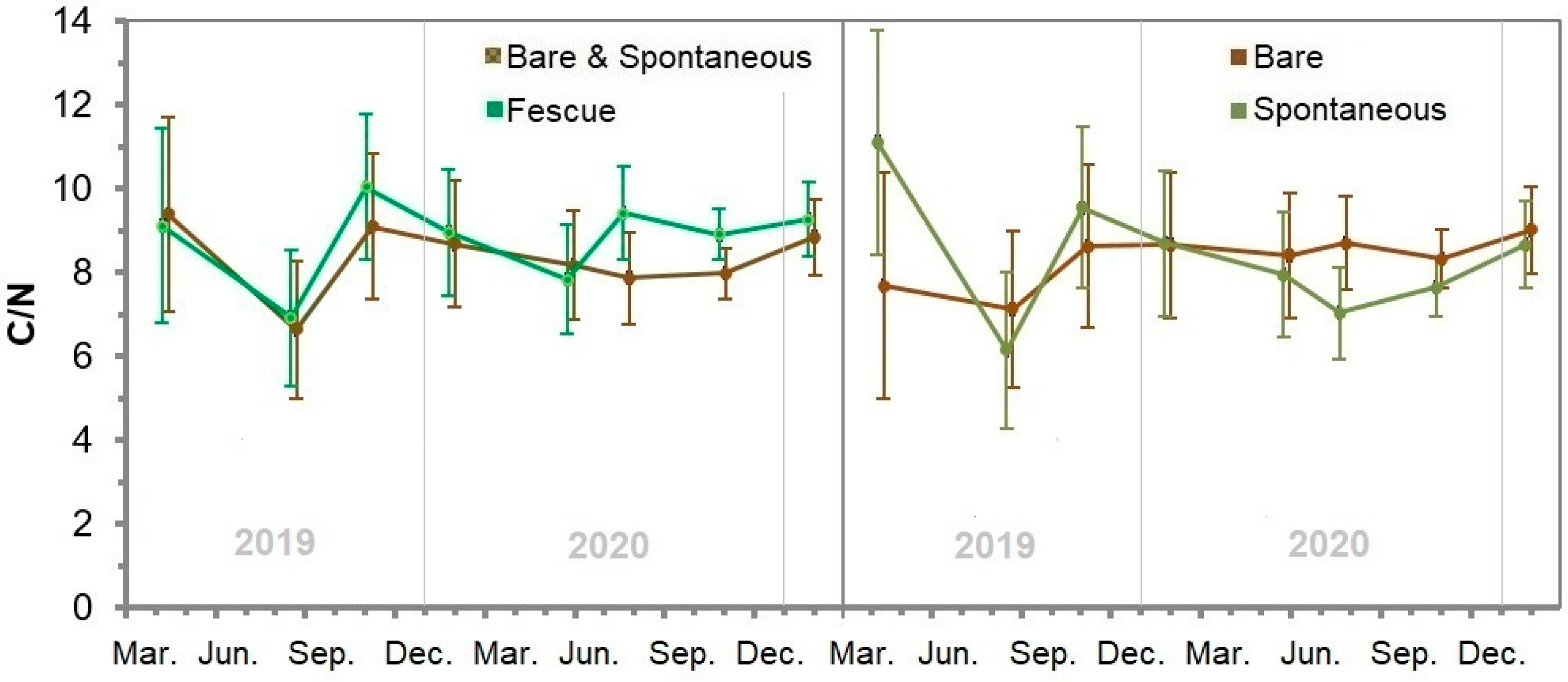
3.13. Soil Nitrogen
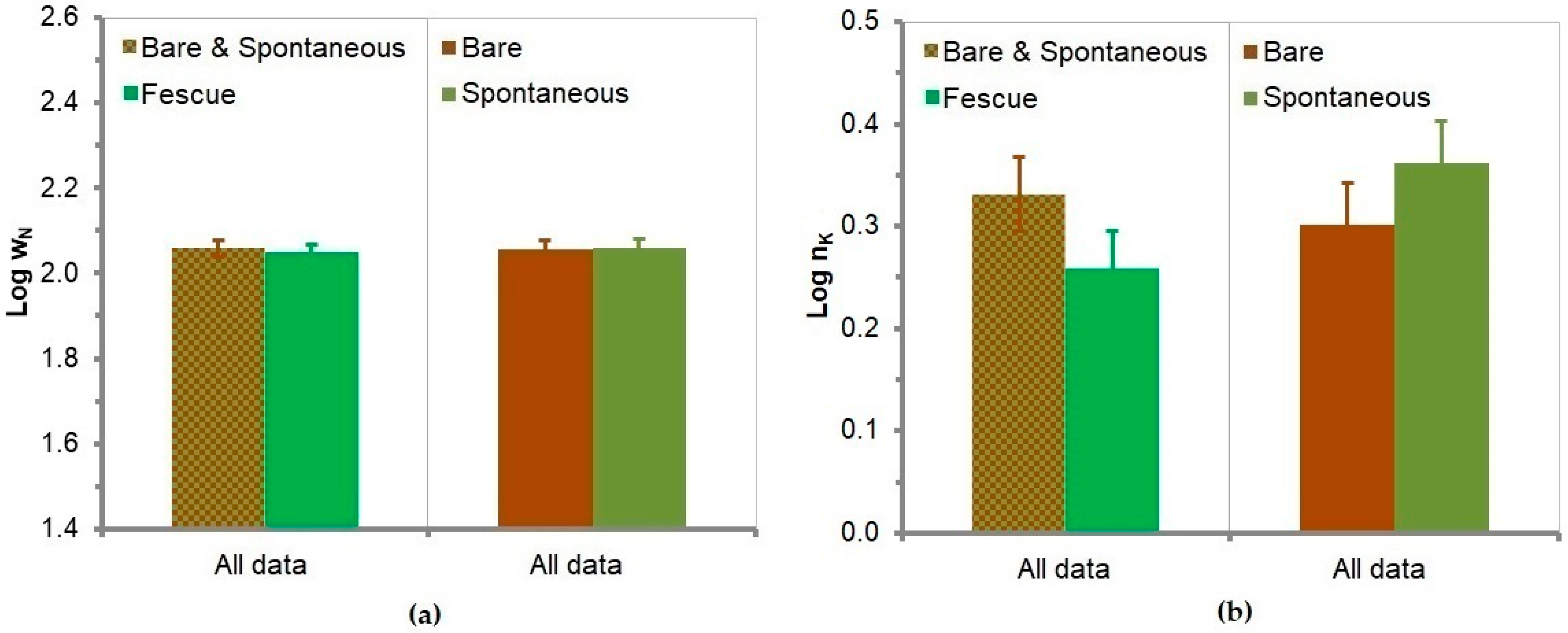
3.14. Soil Potassium
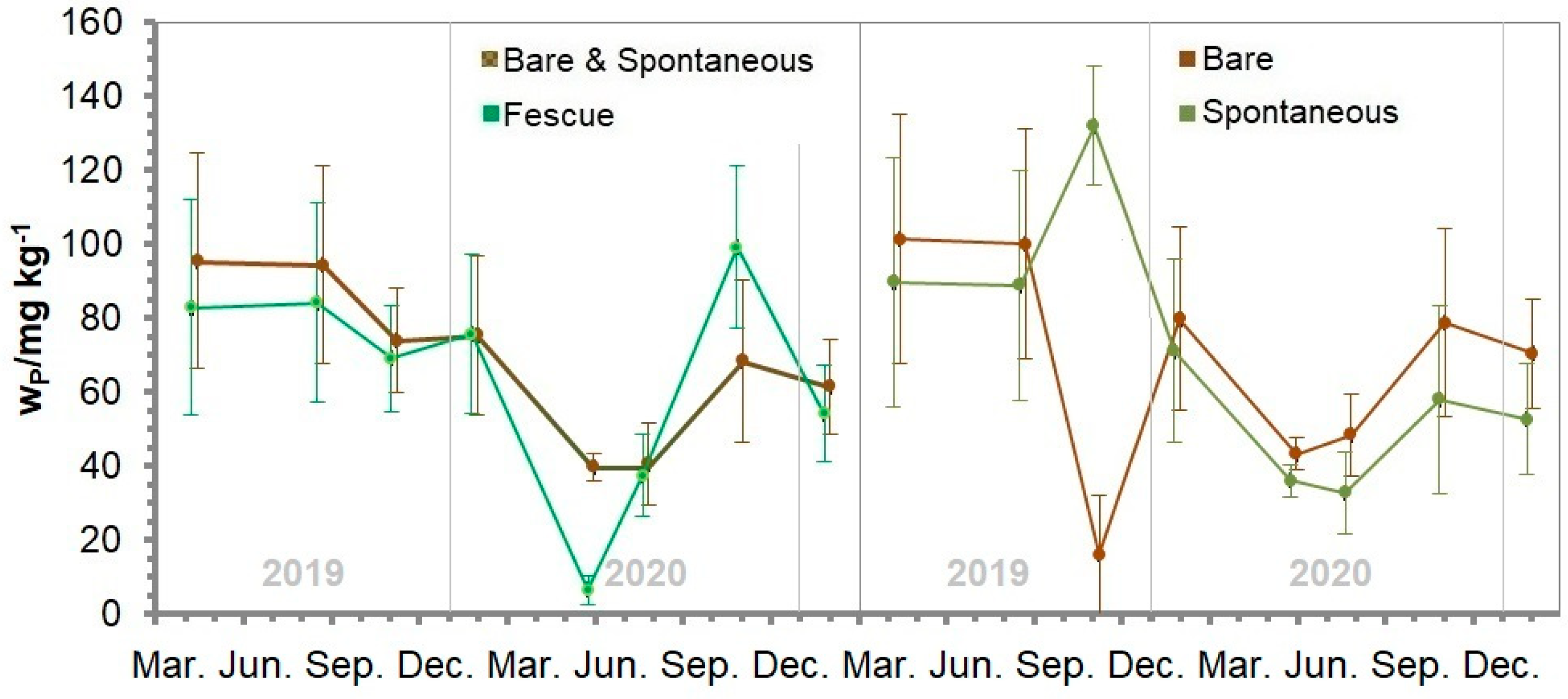
3.15. Soil Phosphorus
4. Discussion
4.1. Soil Characteristics
4.2. Soil Fertility Drivers: Temperature and Water Content
4.3. Soil Organic Carbon
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Abbott, L.K.; Murphy, D.V. Soil Biological Fertility: A Key to Sustainable Land Use in Agriculture; Springer: Dordrecht, The Netherlands, 2007; pp. 1–264. [Google Scholar]
- Pompilica, I.; Romulus, I. Sustainable development of agriculture as means of slowing down the exploitation of nonrenewable natural resources. In Proceedings of the 13th International Multidisciplinary Scientific Geo Conference: Surveying Geology and Mining Ecology Management, SGEM, Albena, Bulgaria, 16–22 June 2013; pp. 685–692. [Google Scholar]
- Melero, S.; Panettieri, M.; Madejón, E.; Macpherson, H.G.; Moreno, F.; Murillo, J.M. Implementation of chiselling and mouldboard ploughing in soil after 8 years of no-till management in SW, Spain: Effect on soil quality. Soil Tillage Res. 2011, 112, 107–113. [Google Scholar] [CrossRef]
- Pagano, M.C.; Dhar, P.P. Arbuscular mycorrhizal fungi under monoculture farming: A review. In Monoculture Farming: Global. Practices, Ecological Impact and Benefits/Drawbacks; Nath, T.K., O’Reilly, P., Eds.; Nova Science Publishers: New York, NY, USA, 2016; pp. 161–170. [Google Scholar]
- Le Gall, M.; Evrard, O.; Dapoigny, A.; Tiecher, T.; Zafar, M.; Minella, J.P.G.; Laceby, J.P.; Ayrault, S. Tracing sediment sources in a subtropical agricultural catchment of southern Brazil cultivated with conventional and conservation farming practices. Land Degrad. Dev. 2017, 28, 1426–1436. [Google Scholar] [CrossRef]
- Xu, Q.; Ling, N.; Chen, H.; Duan, Y.; Wang, S.; Shen, Q.; Vandenkoornhuyse, P. Long-term chemical-only fertilization induces a diversity decline and deep selection on the soil bacteria. mSystems 2020, 5, e00337-20. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, C.V.; Levien, R.; Schwarz, S.F.; Mazurana, M.; Petry, H.B.; Zulpo, L.; Fink, J.A. Physical-hydraulic properties of a sandy loam typic paleudalf soil under organic cultivation of ‘montenegrina’ mandarin (Citrus deliciosa tenore). Rev. Bras. Cienc. Solo. 2014, 38, 1882–1889. [Google Scholar] [CrossRef] [Green Version]
- MAPA. Encuesta Sobre Superficies y Rendimientos de Cultivos. Resultados 2020; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2021; pp. 1–180.
- MAPA. Encuesta de Utilización de Productos Fitosanitarios Campaña 2019. Resultados Agosto 2021; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2021; pp. 1–56.
- Hondebrink, M.A.; Cammeraat, L.H.; Cerdà, A. The impact of agricultural management on selected soil properties in citrus orchards in Eastern Spain: A comparison between conventional and organic citrus orchards with drip and flood irrigation. Sci. Total Environ. 2017, 581, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Morugán-Coronado, A.; García-Orenes, F.; Cerdà, A. Changes in soil microbial activity and physicochemical properties in agricultural soils in eastern Spain. Spanish J. Soil Sci. 2015, 5, 201–213. [Google Scholar] [CrossRef]
- Cerdà, A.; Novara, A.; Moradi, E. Long-term non-sustainable soil erosion rates and soil compaction in drip-irrigated citrus plantation in Eastern Iberian Peninsula. Sci. Total Environ. 2021, 787, 147549. [Google Scholar]
- Di Prima, S.; Rodrigo-Comino, J.; Novara, A.; Iovino, M.; Pirastru, M.; Keesstra, S.; Cerdà, A. Soil Physical Quality of Citrus Orchards Under Tillage, Herbicide, and Organic Managements. Pedosphere 2018, 28, 463–477. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.H.; Wang, L.; Wan, X.G.; Peng, Q.Z.; Huang, Q.; Shi, Z.H. A systematic review of soil erosion in citrus orchards worldwide. Catena 2021, 206, 105558. [Google Scholar]
- Adams, W.A. The effect of organic matter on the bulk and true densities of some uncultivated podzolic soils. J. Soil Sci. 1973, 24, 10–17. [Google Scholar] [CrossRef]
- Gosselink, J.G.; Hatton, R.; Hopkinson, C.S. Relationship of organic carbon and mineral content to bulk density in Louisiana marsh soils. Soil Sci. 1984, 137, 177–180. [Google Scholar] [CrossRef]
- Stavi, I.; Ungar, E.D.; Lavee, H.; Sarah, P. Grazing-induced spatial variability of soil bulk density and content of moisture, organic carbon and calcium carbonate in a semi-arid rangeland. Catena 2008, 75, 288–296. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Russell, J.S. Soil fertility changes in the long-term experimental plots at Klybybolite, South Australia I. Changes in pH, total nitrogen, organic carbon, and bulk density. Australian J. Agr. Res. 1960, 11, 902–926. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Stone, L.R.; Schlegel, A.J.; Lyon, D.J.; Vigil, M.F.; Mikha, M.M.; Stahlman, P.W.; Rice, C.W. No-till induced increase in organic carbon reduces maximum bulk density of soils. Soil Sci. Soc. Am. J. 2009, 73, 1871–1879. [Google Scholar] [CrossRef]
- de Oliveira, F.E.R.; Oliveira, J.M.; Xavier, F.A.S. Changes in soil organic carbon fractions in response to cover crops in an orange orchard. Rev. Bras. Cienc. Solo. 2016, 40, e0150105. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Liu, Y.; Mohamed, I.; Zhang, R.; Wang, X.; Nie, X.; Jiang, M.; Brooks, M.; Chen, F.; Li, Z. Dynamic changes of soil surface organic carbon under different mulching practices in citrus orchards on sloping land. PLoS ONE 2016, 11, e0168384. [Google Scholar] [CrossRef] [Green Version]
- Moreno, B.; García-Rodríguez, S.; Cañizares, R.; Castro, J.; Benítez, E. Rainfed olive farming in south-eastern Spain: Long-term effect of soil management on biological indicators of soil quality. Agric. Ecosyst. Environ. 2009, 131, 333–339. [Google Scholar] [CrossRef]
- Ramos, M.E.; Robles, A.B.; Sánchez-Navarro, A.; González-Rebollar, J.L. Soil responses to different management practices in rainfed orchards in semiarid environments. Soil Tillage Res. 2011, 112, 85–91. [Google Scholar] [CrossRef]
- Herencia, J.F. Enzymatic activities under different cover crop management in a Mediterranean olive orchard. Biol. Agric. Hortic. 2015, 31, 45–52. [Google Scholar] [CrossRef]
- Gavazzi, C.; Schulz, M.; Marocco, A.; Tabaglio, V. Sustainable weed control by allelochemicals from rye cover crops: From the greenhouse to field evidence. Allelopathy J. 2010, 25, 259–273. [Google Scholar]
- Blubaugh, C.K.; Hagler, J.R.; Machtley, S.A.; Kaplan, I. Cover crops increase foraging activity of omnivorous predators in seed patches and facilitate weed biological control. Agric. Ecosyst. Environ. 2016, 231, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Tillman, G.; Schomberg, H.; Phatak, S.; Mullinix, B.; Lachnicht, S.; Timper, P.; Olson, D. Influence of cover crops on insect pests and predators in conservation tillage cotton. J. Econ. Entomol. 2004, 97, 1217–1232. [Google Scholar] [CrossRef]
- Koudahe, K.; Allen, S.C.; Djaman, K. Critical review of the impact of cover crops on soil properties. Int. Soil Water Conserv. Res. 2022, 10, 343–354. [Google Scholar] [CrossRef]
- Rubio-Asensio, J.S.; Hortelano, D.; Ramírez-Cuesta, J.M.; Parra, M.; Buesa, I.; Intrigliolo, D.S. How fertilization regime should be adapted when almond trees are grown under deficit irrigation and with cover crops? Insights and open questions from a field trial. Acta Hortic. 2022, 1333, 351–358. [Google Scholar] [CrossRef]
- Rodríguez-Ballesteros, C. Clasificación Climática de Köppen-Geiger (para España). Periodo de Referencia 1981–2010. Available online: https://climaenmapas.blogspot.com/p/pagina-koppen.html (accessed on 14 April 2021).
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; US Department of Agriculture—Natural Resources Conservation Service: Washington, DC, USA, 2014; pp. 1–372.
- Ingelmo, F.; Molina, M.J.; de Paz, J.M.; Visconti, F. Soil saturated hydraulic conductivity assessment from expert evaluation of field characteristics using an ordered logistic regression model. Soil Tillage Res. 2011, 115, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Pan, L.; Mitchell, J.; Sanden, B. Measuring saturated hydraulic conductivity using a generalized solution for single-ring infiltrometers. Soil Sci. Soc. Am. J. 1999, 63, 788–792. [Google Scholar] [CrossRef]
- Kutzbach, L.; Schneider, J.; Sachs, T.; Giebels, M.; Nykanen, H.; Shurpali, N.J.; Martikainen, P.J.; Alm, J.; Wilmking, M. CO2 flux determination by closed-chamber methods can be seriously biased by inappropriate application of linear regression. Biogeosciences 2007, 4, 1005–1025. [Google Scholar] [CrossRef]
- Livingston, G.P.; Hutchinson, G.L. Enclosure-based measurement of trace gas exchange: Applications and sources of error. In Biogenic Trace Gases: Measuring Emissions from Soil and Water; Matson, P.A., Harris, R.C., Eds.; Blackwell Science Ltd.: Oxford, UK, 1995; pp. 14–51. [Google Scholar]
- Casida, J.L.E.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Bremner, J.M. 1996. Nitrogen total. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA, ASA: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Walkley, A. A critical examination of a rapid method for determining organic carbon in soils: Effects of variation in digestion conditions and of inorganic soil constituents. Soil Sci. 1947, 63, 251–263. [Google Scholar] [CrossRef]
- Visconti, F.; Jiménez, M.G.; de Paz, J.M. How do the chemical characteristics of organic matter explain differences among its determinations in calcareous soils? Geoderma 2022, 406, 115454. [Google Scholar] [CrossRef]
- Kuo, S. Phosphorus. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA, ASA: Madison, WI, USA, 1996; pp. 869–919. [Google Scholar]
- Gee, G.W.; Or, D. Particle-size analysis. In Methods of Soil Analysis. Part 4. Physical Methods; Campbell, G., Horton, R., Jury, W.A., Nielsen, D.R., van Es, H.M., Wierenga, P.J., Dane, J.H., Topp, G.C., Eds.; SSSA, ASA: Madison, WI, USA, 2002; pp. 255–294. [Google Scholar]
- IGME. Mapa Geológico de España E. 1:50.000, Valencia (722); Instituto Geológico y Minero de España. Servicio de Publicaciones Ministerio de Industria: Madrid, España, 1972; p. 1. [Google Scholar]
- GVA. Mapa de Suelos de la Comunidad Valenciana. Valencia (722); Servei d’Estudis Agraris i Comunitaris: Valencia, España, 1996; pp. 1–101. [Google Scholar]
- Rémy, J.C.; Marin-Laflèche, A. L’analyse de la terre: Réalisation d’un programme d’interpretation automatique. Annales Agronomiques 1974, 25, 607–632. [Google Scholar]
- Minasny, B.; Malone, B.P.; McBratney, A.B.; Angers, D.A.; Arrouays, D.; Chambers, A.; Chaplot, V.; Chen, Z.-S.; Cheng, K.; Das, B.S.; et al. Soil carbon 4 per mille. Geoderma 2017, 292, 59–86. [Google Scholar] [CrossRef]
- Alvarez, G.; Shahzad, T.; Andanson, L.; Bahn, M.; Wallenstein, M.D.; Fontaine, S. Catalytic power of enzymes decreases with temperature: New insights for understanding soil C cycling and microbial ecology under warming. Global Chang. Biol. 2018, 24, 4238–4250. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.U.; Bennett, A.E.; Tack, A.J.M. The impact of elevated temperature and drought on the ecology and evolution of plant–soil microbe interactions. J. Ecol. 2020, 108, 337–352. [Google Scholar] [CrossRef]
- Pradel, E.; Pieri, P. Influence of a grass layer on vineyard soil temperature. Aust. J. Grape Wine Res. 2008, 6, 59–67. [Google Scholar] [CrossRef]
- Munsell Color. Munsell Soil Color Charts; Macbeth Division of Kollmorgen Instruments Corporation: New Windsor, NY, USA, 1994; p. 36. [Google Scholar]
- Post, D.F.; Fimbres, A.; Matthias, A.D.; Sano, E.E.; Accioly, L.; Batchily, A.K.; Ferreira, L.G. Predicting Soil Albedo from Soil Color and Spectral Reflectance Data. Soil Sci. Soc. Am. J. 2000, 64, 1027–1034. [Google Scholar] [CrossRef]
- Payero, J.O.; Neale, C.M.U.; Wright, J.L. Near-noon albedo values of alfalfa and tall fescue grass derived from multispectral data. Int. J. Remote Sens. 2006, 27, 569–586. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements; FAO: Rome, Italy, 1998; pp. 1–300. [Google Scholar]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef]
- Sarno, J.L.; Afandi, T.A.; Oki, Y.; Senge, M.; Watanabe, A. Effect of weed management in coffee plantation on soil chemical properties. Nutr. Cycling Agroecosyst. 2004, 69, 1–4. [Google Scholar]
- Johnston, M.B.; Olivares, A.E.; Calderón, C.E. Effect of quantity and distribution of rainfalls on Hordeum murinum L. Growth and development. Chil. J. Agric. Res. 2009, 69, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Matos, E.; Cardoso, I.M.; Souto, R.L.; De Lima, P.C.; De Sá Mendonça, E. Characteristics, residue decomposition, and carbon mineralization of leguminous and spontaneous plants in coffee systems. Commun. Soil Sci. Plant Anal. 2011, 42, 489–502. [Google Scholar] [CrossRef]
- Olego, M.Á.; Cuesta-Lasso, M.D.; Visconti Reluy, F.; López, R.; López-Losada, A.; Garzón-Jimeno, E. Laboratory extractions of soil phosphorus do not reflect the fact that liming increases rye phosphorus content and yield in an acidic soil. Plants 2022, 11, 2871. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Pares, A.; O’Hara, C.; Khanna, P.K.; Bauhus, J. Soil Organic Carbon is Increased in Mixed-Species Plantations of Eucalyptus and Nitrogen-Fixing Acacia. Ecosyst 2013, 16, 123–132. [Google Scholar] [CrossRef]
- Meng, F.; Dungait, J.A.J.; Xu, X.; Bol, R.; Zhang, X.; Wu, W. Coupled incorporation of maize (Zea mays L.) straw with nitrogen fertilizer increased soil organic carbon in Fluvic Cambisol. Geoderma 2017, 304, 19–27. [Google Scholar] [CrossRef]
- Sarma, B.; Gogoi, N. Nitrogen Management for Sustainable Soil Organic Carbon Increase in Inceptisols Under Wheat Cultivation. Commun. Soil Sci. Plant Anal. 2017, 48, 1428–1437. [Google Scholar] [CrossRef]
- Ball, K.R.; Baldock, J.A.; Penfold, C.; Power, S.A.; Woodin, S.J.; Smith, P.; Pendall, E. Soil organic carbon and nitrogen pools are increased by mixed grass and legume cover crops in vineyard agroecosystems: Detecting short-term management effects using infrared spectroscopy. Geoderma 2020, 379, 114619. [Google Scholar] [CrossRef]
- Raphael, J.P.A.; Calonego, J.C.; Milori, D.M.B.P.; Rosolem, C.A. Soil organic matter in crop rotations under no-till. Soil Tillage Res. 2016, 155, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Rocha, K.F.; de Souza, M.; Almeida, D.S.; Chadwick, D.R.; Jones, D.L.; Mooney, S.J.; Rosolem, C.A. Cover crops affect the partial nitrogen balance in a maize-forage cropping system. Geoderma 2020, 360, 114000. [Google Scholar] [CrossRef]
- Kay, B.D.; VandenBygaart, A.J. Conservation tillage and depth stratification of porosity and soil organic matter. Soil Tillage Res. 2002, 66, 107–118. [Google Scholar] [CrossRef]
- Novara, A.; Pulido, M.; Rodrigo-Comino, J.; Prima, S.D.I.; Smith, P.; Gristina, L.; Giménez-Morera, A.; Terol, E.; Salesa, D.; Keesstra, S. Long-term organic farming on a citrus plantation results in soil organic carbon recovery. Geograph. Res. Lett. 2019, 45, 271–286. [Google Scholar] [CrossRef]
- Jury, W.A.; Horton, R. Soil Physics, 6th ed.; John Wiley and Sons: Hoboken, NJ, USA, 2004; pp. 1–370. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth/cm | Texture USDA | wSOM(%) | wCCE(%) | ρb/g cm−3 | |||
|---|---|---|---|---|---|---|---|
| Clay (%) | Silt (%) | Sand (%) | Class | ||||
| 0–20 | 30 | 42 | 29 | Clay loam | 1.4 | 27 | 1.63 |
| 20–40 | 26 | 42 | 32 | Loam | 1.0 | 29 | – |
| 40–60 | 29 | 41 | 31 | Clay loam | 0.6 | 30 | – |
| T | θw | Log Ks | ρb | wroot1/2 | Log DHA | Log GLA | Log EC-CO2 | wSOC | Log wN | Log nK | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| θw | −0.65 *** | ||||||||||
| Log Ks | 0.59 | −0.28 | |||||||||
| ρb | 0.01 | 0.25 | −0.77 ** | ||||||||
| wroot1/2 | −0.43 | 0.70 | −0.33 | 0.70 | |||||||
| Log DHA | 0.50 *** | −0.42 ** | 0.02 | 0.09 | 0.38 | ||||||
| Log GLA | −0.04 | 0.23 | −0.25 | −0.31 | 0.82 | 0.04 | |||||
| Log_ EC-CO2 | −0.11 | 0.36 * | 0.60 | −0.18 | 0.00 | −0.09 | 0.44 * | ||||
| wSOC | −0.03 | 0.12 | 0.19 | 0.02 | 0.20 | 0.27 | 0.18 | −0.15 | |||
| Log wN | 0.67 *** | −0.59 *** | −0.25 | 0.36 | −0.84 * | 0.41 ** | 0.18 | −0.26 | 0.21 | ||
| Log nK | −0.28 | 0.24 | 0.16 | −0.15 | −0.21 | 0.21 | −0.25 | −0.05 | 0.14 | −0.18 | |
| wP | −0.06 | −0.11 | −0.00 | −0.09 | 0.65 | −0.07 | −0.18 | −0.16 | −0.07 | −0.17 | −0.08 |
| Property | Source of Variance | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|---|
| Log Ks | Treatment | 2 | 0.3553 | 0.1777 | 1.869 | 0.1846 |
| Measurement time | 1 | 0.5538 | 0.5538 | 5.827 | 0.0273 | |
| Treat. × Meas. | 2 | 0.0731 | 0.0365 | 0.384 | 0.6866 | |
| Residual | 17 | 1.6157 | 0.095 | |||
| ρb | Treatment | 2 | 0.00889 | 0.004443 | 1.461 | 0.2537 |
| Sampling time | 2 | 0.02897 | 0.014487 | 4.763 | 0.0191 | |
| Treat. × Samp. | 4 | 0.01479 | 0.003698 | 1.216 | 0.3325 | |
| Residual | 22 | 0.06691 | 0.003042 | |||
| wroot1/2 | Treatment | 2 | 2.30754 | 1.15377 | 17.303 | 0.000824 |
| Sampling time | − | − | − | − | − | |
| Treat. × Samp. | − | − | − | − | − | |
| Residual | 9 | 0.6001 | 0.0667 | |||
| Log DHA | Treatment | 2 | 0.294 | 0.1472 | 3.184 | 0.0453 |
| Sampling time | 7 | 4.812 | 0.6874 | 14.865 | 1.85 × 10−13 | |
| Treat. × Samp. | 14 | 0.621 | 0.0443 | 0.959 | 0.4998 | |
| Residual | 108 | 4.995 | 0.0462 | |||
| Log GLA | Treatment | 2 | 1.9993 | 0.9997 | 38.027 | 3.96 × 10−12 |
| Sampling time | 4 | 1.4132 | 0.3533 | 13.44 | 2.63 × 10−8 | |
| Treat. × Samp. | 8 | 0.8462 | 0.1058 | 4.023 | 0.000512 | |
| Residual | 75 | 1.9716 | 0.0263 | |||
| EC-CO2 | Treatment | 2 | 0.356 | 0.1779 | 1.822 | 0.166 |
| Measurement time | 15 | 11.206 | 0.7471 | 7.652 | 6.29 × 10−12 | |
| Treat. × Meas. | 30 | 3.694 | 0.1231 | 1.261 | 0.189 | |
| Residual | 127 | 12.399 | 0.0976 | |||
| wSOC | Treatment | 2 | 0.0056 | 0.00279 | 0.266 | 0.767 |
| Sampling time | 9 | 0.6758 | 0.07509 | 7.15 | 2.49 × 10−8 | |
| Treat. × Samp. | 18 | 0.2094 | 0.01163 | 1.108 | 0.353 | |
| Residual | 126 | 1.3232 | 0.0105 | |||
| Log wN | Treatment | 2 | 0.0024 | 0.0012 | 0.512 | 0.601 |
| Sampling time | 7 | 0.18809 | 0.02687 | 11.471 | 1.02 × 10−10 | |
| Treat. × Samp. | 14 | 0.05042 | 0.003602 | 1.538 | 0.111 | |
| Residual | 104 | 0.24361 | 0.002342 | |||
| Log nK | Treatment | 2 | 0.2278 | 0.11391 | 12.178 | 0.0000177 |
| Sampling time | 7 | 0.4001 | 0.05716 | 6.11 | 0.00000544 | |
| Treat. × Samp. | 14 | 0.1858 | 0.01327 | 1.419 | 0.157 | |
| Residual | 104 | 0.9728 | 0.00935 | |||
| wP | Treatment | 2 | 370 | 185 | 0.763 | 0.469 |
| Sampling time | 7 | 56819 | 8117 | 33.497 | <2 × 10−16 | |
| Treat. × Samp. | 14 | 49095 | 3507 | 14.472 | <2 × 10−16 | |
| Residual | 104 | 25201 | 242 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visconti, F.; Peiró, E.; Baixauli, C.; de Paz, J.M. Spontaneous Plants Improve the Inter-Row Soil Fertility in a Citrus Orchard but Nitrogen Lacks to Boost Organic Carbon. Environments 2022, 9, 151. https://doi.org/10.3390/environments9120151
Visconti F, Peiró E, Baixauli C, de Paz JM. Spontaneous Plants Improve the Inter-Row Soil Fertility in a Citrus Orchard but Nitrogen Lacks to Boost Organic Carbon. Environments. 2022; 9(12):151. https://doi.org/10.3390/environments9120151
Chicago/Turabian StyleVisconti, Fernando, Enrique Peiró, Carlos Baixauli, and José Miguel de Paz. 2022. "Spontaneous Plants Improve the Inter-Row Soil Fertility in a Citrus Orchard but Nitrogen Lacks to Boost Organic Carbon" Environments 9, no. 12: 151. https://doi.org/10.3390/environments9120151
APA StyleVisconti, F., Peiró, E., Baixauli, C., & de Paz, J. M. (2022). Spontaneous Plants Improve the Inter-Row Soil Fertility in a Citrus Orchard but Nitrogen Lacks to Boost Organic Carbon. Environments, 9(12), 151. https://doi.org/10.3390/environments9120151