Salinomycin Treatment Specifically Inhibits Cell Proliferation of Cancer Stem Cells Revealed by Longitudinal Single Cell Tracking in Combination with Fluorescence Microscopy



{kind=link}
{kind=link}
{kind=link}
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culturing
2.2. Cell Seeding
2.3. Treatment
2.4. Digital Holographic Time-Lapse Imaging in Combination with Fluorescence and Tracking
2.5. Fluorescence Labeling
2.6. Fluorescence Quantification
2.7. Longitudinal Tracking of Cells Using Time-Lapses and Matching with Fluorescence Images
2.8. Statistics
3. Results
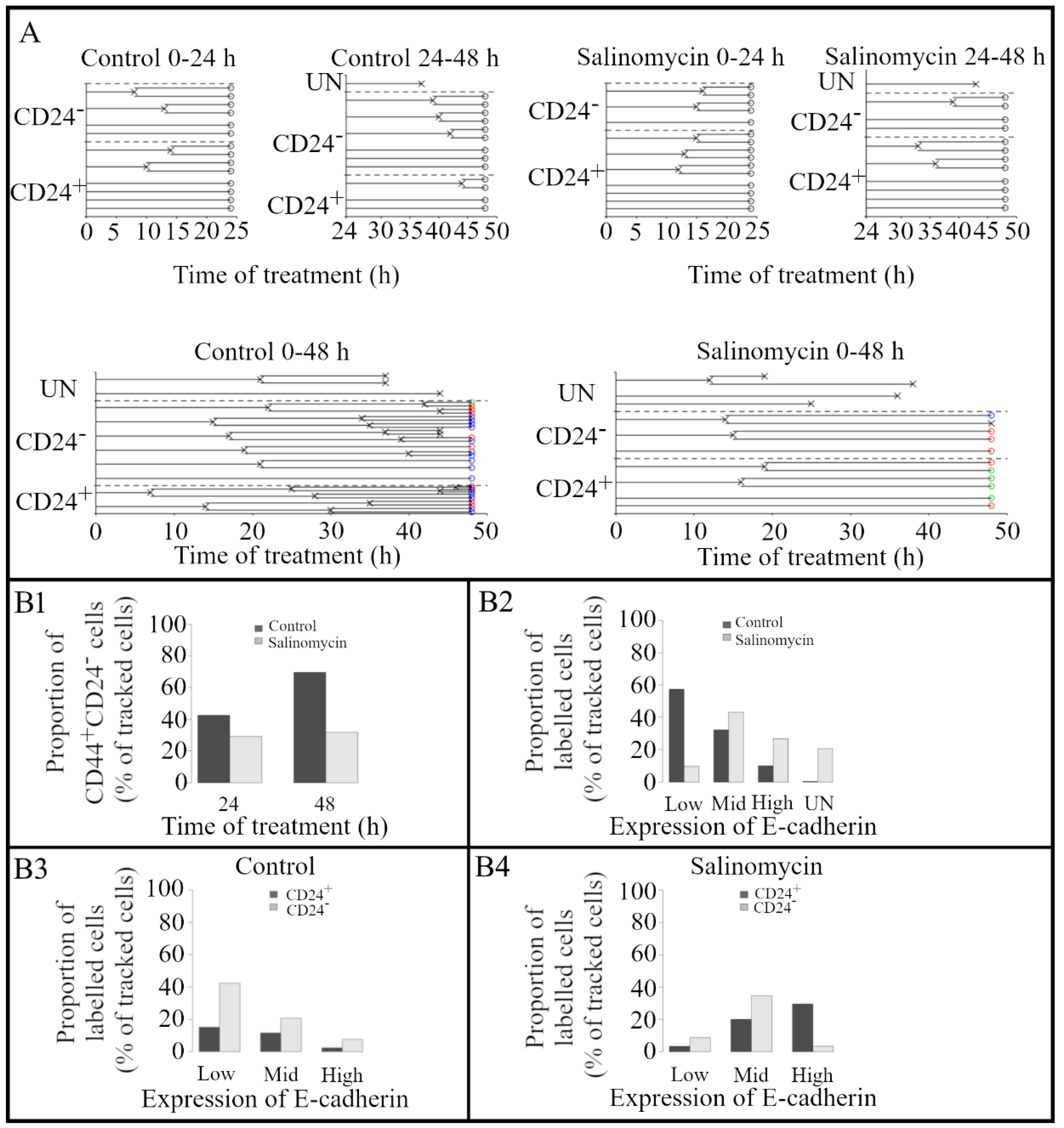
3.1. Evaluating the Effect of Salinomycin on Proliferation Using a Combination of Longitudinal Tracking of Time-Lapses Obtained with DHM and Fluorescence Microscopy
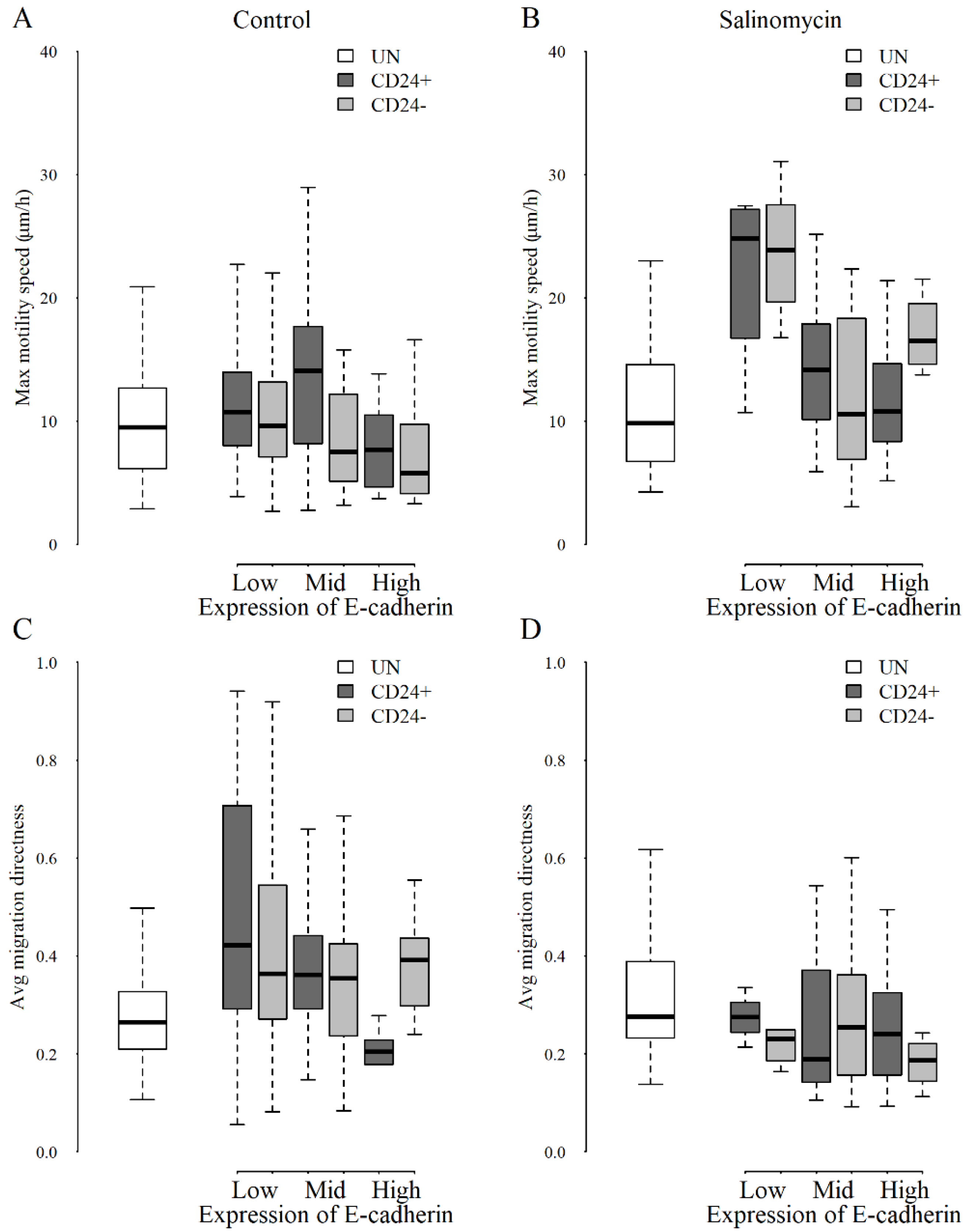
3.2. Longitudinal Tracking of Effects of Salinomycin on Motility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sneddon, J.B. Cancer stem cells. Adv. Struct. Saf. Stud. 2009, 568, 217–232. [Google Scholar] [CrossRef]
- Ailles, L.E.; Weissman, I.L. Cancer stem cells in solid tumors. Curr. Opin. Biotechnol. 2007, 18, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Becker, M.; Wicha, M.; Weissman, I.; Clarke, M.F. Therapeutic implications of cancer stem cells. Curr. Opin. Genet. Dev. 2004, 14, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Bedard, P.L.; Cardoso, F.; Piccart-Gebhart, M.J. Stemming Resistance to HER-2 Targeted Therapy. J. Mammary Gland. Boil. Neoplasia 2009, 14, 55–66. [Google Scholar] [CrossRef]
- Dick, J. Breast cancer stem cells revealed. Proc. Natl. Acad. Sci. USA 2003, 100, 3547–3549. [Google Scholar] [CrossRef] [Green Version]
- Al-hajj, M.; Wicha, M.S.; Benito-hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.B.; Önder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.; Lander, E.S. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, D.; Daniel, V.; Sadeghi, M.; Opelz, G.; Naujokat, C. Salinomycin overcomes ABC transporter-mediated multidrug and apoptosis resistance in human leukemia stem cell-like KG-1a cells. Biochem. Biophys. Res. Commun. 2010, 394, 1098–1104. [Google Scholar] [CrossRef]
- Zhi, Q.M.; Chen, X.H.; Ji, J.; Zhang, J.N.; Li, J.F.; Cai, Q.; Liu, B.Y.; Gu, Q.L.; Zhu, Z.G.; Yu, Y.Y. Salinomycin can effectively kill ALDHhigh stem-like cells on gastric cancer. Biomed. Pharmacother. 2011, 65, 509–515. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.; Xue, X.; He, S.; Kuang, Y.; Zhao, H.; Chen, S.; Zhi, Q.; Guo, X. Salinomycin induces apoptosis in cisplatin-resistant colorectal cancer cells by accumulation of reactive oxygen species. Toxicol. Lett. 2013, 222, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.-L.; Zhao, Z.-Q.; Li, J.-C.; Liang, Y.; Yin, J.-Q.; Zou, C.-Y.; Xie, X.-B.; Zeng, Y.-X.; Shen, J.-N.; Kang, T.; et al. Salinomycin inhibits osteosarcoma by targeting its tumor stem cells. Cancer Lett. 2011, 311, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-N.; Liang, Y.; Zhou, L.-J.; Chen, S.-P.; Chen, G.; Zhang, T.; Kang, T.; Zhao, Y. Combination of salinomycin and gemcitabine eliminates pancreatic cancer cells. Cancer Lett. 2011, 313, 137–144. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, F.; Dai, W.-Q.; Wu, D.; Lin, C.-L.; Wu, S.-M.; Cheng, P.; Zhang, Y.; Shen, M.; Wang, C.-F.; et al. Mechanism of action of salinomycin on growth and migration in pancreatic cancer cell lines. Pancreatology 2013, 13, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ketola, K.; Hilvo, M.; Hyötyläinen, T.; Vuoristo, A.; Ruskeepää, A.-L.; Orešič, M.; Kallioniemi, O.; Iljin, K. Salinomycin inhibits prostate cancer growth and migration via induction of oxidative stress. Br. J. Cancer 2012, 106, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Shang, Z.; Cai, Q.; Zhang, M.; Zhu, S.; Ma, Y.; Sun, L.; Jiang, N.; Tian, J.; Niu, X.; Chen, J.; et al. A switch from CD44+ cell to EMT cell drives the metastasis of prostate cancer. Oncotarget 2014, 6, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.Z.; Blair, K.J.; Rahimy, E.; Kiang, A.; Abhold, E.; Fan, J.-B.; Wang-Rodriguez, J.; Altuna, X.; Ongkeko, W. Salinomycin induces cell death and differentiation in head and neck squamous cell carcinoma stem cells despite activation of epithelial-mesenchymal transition and Akt. BMC Cancer 2012, 12, 556. [Google Scholar] [CrossRef] [Green Version]
- Larzabal, L.; El-Nikhely, N.; Redrado, M.; Seeger, W.; Savai, R.; Calvo, A. Differential effects of drugs targeting cancer stem cell (CSC) and non-CSC populations on lung primary tumors and msetastasis. PLoS ONE 2013, 8, e79798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenk, M.; Aykut, B.; Teske, C.; Giese, N.A.; Weitz, J.; Welsch, T. Salinomycin inhibits growth of pancreatic cancer and cancer cell migration by disruption of actin stress fiber integrity. Cancer Lett. 2015, 358, 161–169. [Google Scholar] [CrossRef]
- Lee, H.-G.; Shin, S.-J.; Chung, H.-W.; Kwon, S.-H.; Cha, S.-D.; Lee, J.-E.; Cho, C.-H. Salinomycin reduces stemness and induces apoptosis on human ovarian cancer stem cell. J. Gynecol. Oncol. 2017, 28, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jangamreddy, J.R.; Ghavami, S.; Grabarek, J.; Kratz, G.; Wiechec, E.; Fredriksson, B.-A.; Pariti, R.K.R.; Cieślar-Pobuda, A.; Panigrahi, S.; Łos, M.J. Salinomycin induces activation of autophagy, mitophagy and affects mitochondrial polarity: Differences between primary and cancer cells. Biochim. Biophys. Acta Bioenerg. 2013, 1833, 2057–2069. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Choi, Y.-J.; Kim, K.-Y.; Yu, S.-N.; Seo, Y.-K.; Chun, S.-S.; Noh, K.-T.; Suh, J.-T.; Ahn, S.-C. Salinomycin simultaneously induces apoptosis and autophagy through generation of reactive oxygen species in osteosarcoma U2OS cells. Biochem. Biophys. Res. Commun. 2016, 473, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Boehmerle, W.; Endres, M. Salinomycin induces calpain and cytochrome c-mediated neuronal cell death. Cell Death Dis. 2011, 2, e168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdoodt, B.; Vogt, M.; Schmitz, I.; Liffers, S.-T.; Tannapfel, A.; Mirmohammadsadegh, A. Salinomycin induces autophagy in colon and breast cancer cells with concomitant generation of reactive oxygen species. PLoS ONE 2012, 7, e44132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, T.T.; Hamaï, A.; Hienzsch, A.; Cañeque, T.; Müller, S.; Wicinski, J.; Cabaud, O.; Leroy, C.; David, A.; Acevedo, V.; et al. Salinomycin kills cancer stem cells by sequestering iron in lysosomes. Nat. Chem. 2017, 9, 1025–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Dhaheri, Y.; Attoub, S.; Arafat, K.; AbuQamar, S.F.; Eid, A.; Al Faresi, N.; Iratni, R. Salinomycin induces apoptosis and senescence in breast cancer: Upregulation of p21, downregulation of survivin and histone H3 and H4 hyperacetylation. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3121–3135. [Google Scholar] [CrossRef]
- An, H.; Kim, J.Y.; Oh, E.; Lee, N.; Cho, Y.; Seo, J.H. Salinomycin promotes anoikis and decreases the CD44+/CD24− stem-like population via inhibition of STAT3 activation in MDA-MB-231 cells. PLoS ONE 2015, 10, e0141919. [Google Scholar] [CrossRef] [Green Version]
- An, H.; Kim, J.Y.; Lee, N.; Cho, Y.; Oh, E.; Seo, J.H. Salinomycin possesses anti-tumor activity and inhibits breast cancer stem-like cells via an apoptosis-independent pathway. Biochem. Biophys. Res. Commun. 2015, 466, 696–703. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, Y.; Li, Q.; Mao, J.; Hou, Z.; Yu, X.; Fan, S.; Li, J.; Gao, T.; Yan, B.; et al. Salinomycin suppresses TGF-β1-induced epithelial-to-mesenchymal transition in MCF-7 human breast cancer cells. Chem. Int. 2016, 248, 74–81. [Google Scholar] [CrossRef]
- Kopp, F.; Hermawan, A.; Oak, P.S.; Herrmann, A.; Wagner, E.; Roidl, A. Salinomycin treatment reduces metastatic tumor burden by hampering cancer cell migration. Mol. Cancer 2014, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Borgström, B.; Kempengren, S.; Persson, L.; Hegardt, C.; Strand, D.; Oredsson, S. Breast cancer stem cell selectivity of synthetic nanomolar-active salinomycin analogs. BMC Cancer 2016, 16, 145. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Hermawan, A.; Oak, P.S.; Ulaganathan, V.K.; Herrmann, A.; Elnikhely, N.; Thakur, C.; Xiao, Z.; Knyazev, P.; Ataseven, B.; et al. Sequential salinomycin treatment results in resistance formation through clonal selection of epithelial-like tumor cells. Transl. Oncol. 2014, 7, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeichi, M. Cadherins: A molecular family important in selective cell-cell adhesion Annu. Rev. Biochem. 1990, 59, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Takeichi, M. The cadherins: Cell-cell adhesion molecules controlling animal morphogenesis. Development 1988, 102, 639–655. [Google Scholar] [PubMed]
- Chekhun, S.; Bezdenezhnykh, N.; Shvets, J.; Lukianova, N. Expression of biomarkers related to cell adhesion, metastasis and invasion of breast cancer cell lines of different molecular subtype. Exp. Oncol. 2013, 35, 174–179. [Google Scholar] [PubMed]
- Popescu, C.I.; Giuşcă, S.E.; Liliac, L.; Avadanei, R.; Ceauşu, R.; Cimpean, A.M.; Balan, R.; Amalinei, C.; Apostol, D.C.; Caruntu, I.D. E-cadherin expression in molecular types of breast carcinoma. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2013, 54, 267–273. [Google Scholar]
- Fulga, V.; Rudico, L.; Balica, A.R.; Cimpean, A.M.; Saptefrati, L.; Margan, M.-M.; Raica, M. Differential expression of e-cadherin in primary breast cancer and corresponding lymph node metastases. Anticancer Res. 2015, 35, 759–765. [Google Scholar]
- Huang, X.; Borgström, B.; Stegmayr, J.; Abassi, Y.; Kruszyk, M.; Leffler, H.; Persson, L.; Albinsson, S.; Massoumi, R.; Scheblykin, I.G.; et al. The molecular basis for inhibition of stemlike cancer cells by salinomycin. ACS Cent. Sci. 2018, 4, 760–767. [Google Scholar] [CrossRef]
- Lu, D.; Choi, M.Y.; Yu, J.; Castro, J.E.; Kipps, T.J.; Carson, D.A. Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocytic leukemia cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13253–13257. [Google Scholar] [CrossRef] [Green Version]
- Alm, K.; El-Schich, Z.; Falck, M.; Wingren, A.G.; Janicke, B.; Oredsso, S. Cells and holograms—holograms and digital holographic microscopy as a tool to study the morphology of living cells. Hologr. Basic Princ. Contemp. Appl. 2013, 335–351. [Google Scholar]
- Burke, R.T.; Orth, J.D. Through the looking glass: Time-lapse microscopy and longitudinal tracking of single cells to study anti-cancer therapeutics. J. Vis. Exp. 2016, 111, e53994. [Google Scholar] [CrossRef]
- Kamlund, S.; Strand, D.; Janicke, B.; Alm, K.; Oredsson, S. Influence of salinomycin treatment on division and movement of individual cancer cells cultured in normoxia or hypoxia evaluated with time-lapse digital holographic microscopy. Cell Cycle 2017, 16, 2128–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hejna, M.; Jorapur, A.; Song, J.S.; Judson-Torres, R.L. High accuracy label-free classification of kinetic cell states from holographic cytometry. Reports 2017, 7, 127449. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Borgström, B.; Månsson, L.K.; Persson, L.; Oredsson, S.; Hegardt, C.; Strand, D. Semisynthesis of SY-1 for Investigation of breast cancer stem cell selectivity of C-ring-modified salinomycin analogues. ACS Chem. Boil. 2014, 9, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
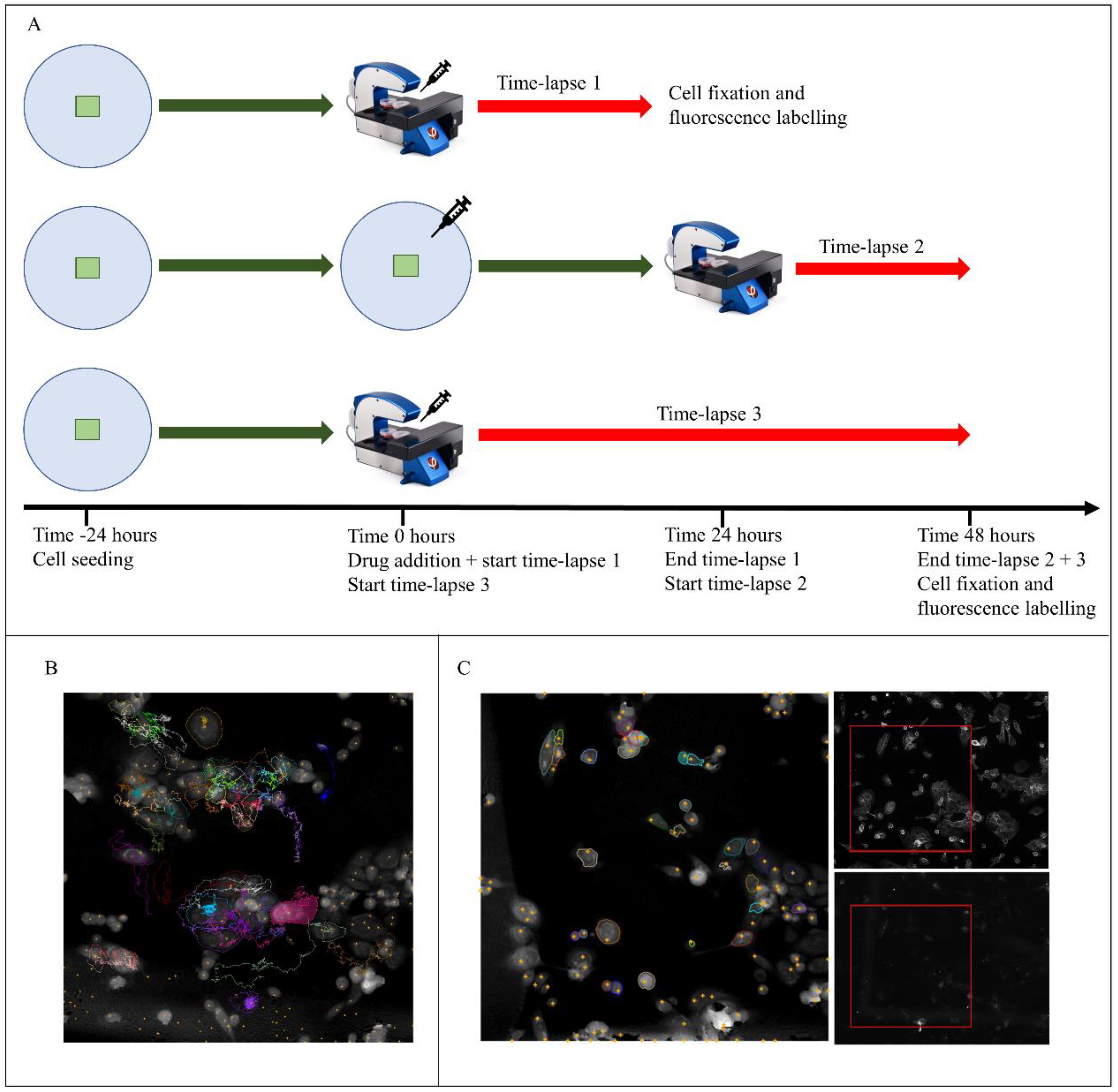
 ). Salinomycin was added to a final concentration of 0.5 µM 24 h after seeding (
). Salinomycin was added to a final concentration of 0.5 µM 24 h after seeding (  ). The cells inside the marked area were imaged by digital holography for 24-h or 48-h periods (
). The cells inside the marked area were imaged by digital holography for 24-h or 48-h periods (  , Time-lapse 3), with the start of the time-lapses immediately after treatment (
, Time-lapse 3), with the start of the time-lapses immediately after treatment (  , Time-lapse 1) or 24 h after treatment (
, Time-lapse 1) or 24 h after treatment (  , Time-lapse 2). After the time-lapse, the cells were fixed and labeled with FITC-conjugated anti-CD44 antibodies and PE-conjugated anti-CD24-antibodies or labeled with Alexa Flour 488-labeled anti-E-cadherin and PE-conjugated anti-CD24-antibodies and then imaged in a fluorescence microscope. (B) Representative image of cells tracked through a time-lapse of digital holographic images. The colored trails show cell movement during the time-lapse. The tracking is the basis for all longitudinal analysis. (C) Representative images used for visual over-lay, which allowed for cell identification in the time-lapse. Left image: digital holography. Upper right image: fluorescence, CD44 staining. Lower right image: fluorescence, CD24 staining. The red box corresponds to the area image by digital holography.
). Salinomycin was added to a final concentration of 0.5 µM 24 h after seeding ( ). The cells inside the marked area were imaged by digital holography for 24-h or 48-h periods ( , Time-lapse 3), with the start of the time-lapses immediately after treatment ( , Time-lapse 1) or 24 h after treatment ( , Time-lapse 2). After the time-lapse, the cells were fixed and labeled with FITC-conjugated anti-CD44 antibodies and PE-conjugated anti-CD24-antibodies or labeled with Alexa Flour 488-labeled anti-E-cadherin and PE-conjugated anti-CD24-antibodies and then imaged in a fluorescence microscope. (B) Representative image of cells tracked through a time-lapse of digital holographic images. The colored trails show cell movement during the time-lapse. The tracking is the basis for all longitudinal analysis. (C) Representative images used for visual over-lay, which allowed for cell identification in the time-lapse. Left image: digital holography. Upper right image: fluorescence, CD44 staining. Lower right image: fluorescence, CD24 staining. The red box corresponds to the area image by digital holography.
, Time-lapse 2). After the time-lapse, the cells were fixed and labeled with FITC-conjugated anti-CD44 antibodies and PE-conjugated anti-CD24-antibodies or labeled with Alexa Flour 488-labeled anti-E-cadherin and PE-conjugated anti-CD24-antibodies and then imaged in a fluorescence microscope. (B) Representative image of cells tracked through a time-lapse of digital holographic images. The colored trails show cell movement during the time-lapse. The tracking is the basis for all longitudinal analysis. (C) Representative images used for visual over-lay, which allowed for cell identification in the time-lapse. Left image: digital holography. Upper right image: fluorescence, CD44 staining. Lower right image: fluorescence, CD24 staining. The red box corresponds to the area image by digital holography.
). Salinomycin was added to a final concentration of 0.5 µM 24 h after seeding ( ). The cells inside the marked area were imaged by digital holography for 24-h or 48-h periods ( , Time-lapse 3), with the start of the time-lapses immediately after treatment ( , Time-lapse 1) or 24 h after treatment ( , Time-lapse 2). After the time-lapse, the cells were fixed and labeled with FITC-conjugated anti-CD44 antibodies and PE-conjugated anti-CD24-antibodies or labeled with Alexa Flour 488-labeled anti-E-cadherin and PE-conjugated anti-CD24-antibodies and then imaged in a fluorescence microscope. (B) Representative image of cells tracked through a time-lapse of digital holographic images. The colored trails show cell movement during the time-lapse. The tracking is the basis for all longitudinal analysis. (C) Representative images used for visual over-lay, which allowed for cell identification in the time-lapse. Left image: digital holography. Upper right image: fluorescence, CD44 staining. Lower right image: fluorescence, CD24 staining. The red box corresponds to the area image by digital holography.


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamlund, S.; Janicke, B.; Alm, K.; Oredsson, S. Salinomycin Treatment Specifically Inhibits Cell Proliferation of Cancer Stem Cells Revealed by Longitudinal Single Cell Tracking in Combination with Fluorescence Microscopy. Appl. Sci. 2020, 10, 4732. https://doi.org/10.3390/app10144732
Kamlund S, Janicke B, Alm K, Oredsson S. Salinomycin Treatment Specifically Inhibits Cell Proliferation of Cancer Stem Cells Revealed by Longitudinal Single Cell Tracking in Combination with Fluorescence Microscopy. Applied Sciences. 2020; 10(14):4732. https://doi.org/10.3390/app10144732
Chicago/Turabian StyleKamlund, Sofia, Birgit Janicke, Kersti Alm, and Stina Oredsson. 2020. "Salinomycin Treatment Specifically Inhibits Cell Proliferation of Cancer Stem Cells Revealed by Longitudinal Single Cell Tracking in Combination with Fluorescence Microscopy" Applied Sciences 10, no. 14: 4732. https://doi.org/10.3390/app10144732
APA StyleKamlund, S., Janicke, B., Alm, K., & Oredsson, S. (2020). Salinomycin Treatment Specifically Inhibits Cell Proliferation of Cancer Stem Cells Revealed by Longitudinal Single Cell Tracking in Combination with Fluorescence Microscopy. Applied Sciences, 10(14), 4732. https://doi.org/10.3390/app10144732