Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments

 ,
,  ,
,  , , ,
, , ,  ,
,  ,
,  , ,
, ,
Abstract
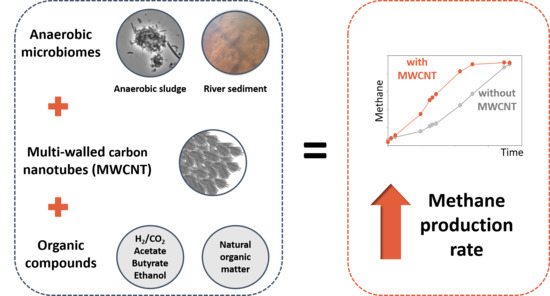
:
1. Introduction
2. Materials and Methods
2.1. Nanomaterials
2.2. Effect of MWCNT on Methane Production by Anaerobic Sludge
2.3. Effect of MWCNT on Methane Production by the Indigenous Microorganisms in River Sediment
2.4. Analytical Methods
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effect of MWCNT on Methane Production by Anaerobic Sludge
3.2. Effect of MWCNT on Methane Production by the Indigenous Microorganisms in River Sediment
4. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fagerström, A.; Al Seadi, T.; Rasi, S.; Briseid, T. The Role of Anaerobic Digestion and Biogas in the Circular Economy; Murphy, J.D., Ed.; IEA Bioenergy Task 37; IEA Bioenergy: Paris, France, 2018; pp. 1–24. [Google Scholar]
- Muhammad Nasir, I.; Mohd Ghazi, T.I.; Omar, R. Production of biogas from solid organic wastes through anaerobic digestion: A review. Appl. Microbiol. Biotechnol. 2012, 95, 321–329. [Google Scholar] [CrossRef]
- Castellano-Hinojosa, A.; Armato, C.; Pozo, C.; González-Martínez, A.; González-López, J. New concepts in anaerobic digestion processes: Recent advances and biological aspects. Appl. Microbiol. Biotechnol. 2018, 102, 5065–5076. [Google Scholar] [CrossRef] [PubMed]
- Náthia-Neves, G.; Berni, M.; Dragone, G.; Mussatto, S.I.; Forster-Carneiro, T. Anaerobic digestion process: Technological aspects and recent developments. Int. J. Environ. Sci. Technol. 2018, 15, 2033–2046. [Google Scholar] [CrossRef]
- Scarlat, N.; Dallemand, J.-F.; Fahl, F. Biogas: Developments and perspectives in Europe. Renew. Energy 2018, 129, 457–472. [Google Scholar] [CrossRef]
- Child, M.; Kemfert, C.; Bogdanov, D.; Breyer, C. Flexible electricity generation, grid exchange and storage for the transition to a 100% renewable energy system in Europe. Renew. Energy 2019, 139, 80–101. [Google Scholar] [CrossRef]
- Van Lier, J.B.; Mahmoud, N.; Zeeman, G. Anaerobic wastewater treatment. In Biological Wastewater Treatment, Principles, Modelling and Design; Henze, M., van Loosdrecht, M.C.M., Ekama, G.A., Brdjanovic, D., Eds.; IWA Publishing: London, UK, 2008; Chapter 16; pp. 401–442. [Google Scholar]
- Angelidaki, I.; Karakashev, D.; Batstone, D.J.; Plugge, C.M.; Stams, A.J. Biomethanation and its potential. Methods Enzymol. 2011, 494, 327–351. [Google Scholar] [CrossRef] [PubMed]
- McInerney, M.J.; Struchtemeyer, C.G.; Sieber, J.; Mouttaki, H.; Stams, A.J.M.; Schink, B.; Rohlin, L.; Gunsalus, R.P. Physiology, ecology, phylogeny, and genomics of microorganisms capable of syntrophic metabolism. Ann. N. Y. Acad. Sci. 2008, 1125, 58–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stams, A.J.M.; Plugge, C.M. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 2009, 7, 568–577. [Google Scholar] [CrossRef]
- Sieber, J.R.; McInerney, M.J.; Gunsalus, R.P. Genomic insights into syntrophy: The paradigm for anaerobic metabolic cooperation. Annu. Rev. Microbiol. 2012, 66, 429–452. [Google Scholar] [CrossRef]
- Nguyen, D.; Wu, Z.; Shrestha, S.; Lee, P.H.; Raskin, L.; Khanal, S.K. Intermittent micro-aeration: New strategy to control volatile fatty acid accumulation in high organic loading anaerobic digestion. Water Res. 2019, 166, 115080. [Google Scholar] [CrossRef]
- González, J.; Sánchez, M.E.; Gómez, X. Enhancing anaerobic digestion: The effect of carbon conductive materials. C—J. Carb. Res. 2018, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.; Salvador, A.F.; Pereira, L.; Alves, M.M. Methane production and conductive materials: A critical review. Environ. Sci. Technol. 2018, 52, 10241–10253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhao, W.; Zhang, H.; Wang, Z.; Fan, C.; Zang, L. Recent achievements in enhancing anaerobic digestion with carbon-based functional materials. Bioresour. Technol. 2018, 266, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, S.; Liang, D.; Li, N. Conductive materials in anaerobic digestion: From mechanism to application. Bioresour. Technol. 2020, 298, 122403. [Google Scholar] [CrossRef]
- Shrestha, P.M.; Rotaru, A.-E.; Aklujkar, M.; Liu, F.; Shrestha, M.; Summers, Z.M.; Malvankar, N.; Flores, D.C.; Lovley, D.R. Syntrophic growth with direct interspecies electron transfer as the primary mechanism for energy exchange. Environ. Microbiol. Rep. 2013, 5, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Nevin, K.P.; Woodard, T.L.; Mu, B.Z.; Lovley, D.R. Expanding the diet for DIET: Electron donors supporting direct interspecies electron transfer (DIET) in defined co-cultures. Front. Microbiol. 2016, 7, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovley, D.R. Happy together: Microbial communities that hook up to swap electrons. ISME J. 2017, 11, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Baek, G.; Kim, J.; Kim, J.; Lee, C. Role and potential of direct interspecies electron transfer in anaerobic digestion. Energies 2018, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Rotaru, A.E.; Shrestha, P.M.; Malvankar, N.S.; Nevin, K.P.; Lovley, D.R. Promoting direct interspecies electron transfer with activated carbon. Energy Environ. Sci. 2012, 5, 8982–8989. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, J.L. Functionalization of porous carbons for catalytic applications. J. Mater. Chem. A 2013, 1, 9351–9364. [Google Scholar] [CrossRef]
- Xu, S.; He, C.; Luo, L.; Lü, F.; He, P.; Cui, L. Comparing activated carbon of different particle sizes on enhancing methane generation in upflow anaerobic digester. Bioresour. Technol. 2015, 196, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Lee, S.-H.; Park, H.-D. Enrichment of specific electro-active microorganisms and enhancement of methane production by adding granular activated carbon in anaerobic reactors. Bioresour. Technol. 2016, 205, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, Y.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Enhancing syntrophic metabolism in up-flow anaerobic sludge blanket reactors with conductive carbon materials. Bioresour. Technol. 2015, 191, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, Y. Conductive Fe3O4 nanoparticles accelerate syntrophic methane production from butyrate oxidation in two different lake sediments. Front. Microbiol. 2016, 7, 1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, A.F.; Martins, G.; Melle-Franco, M.; Serpa, R.; Stams, A.J.M.; Cavaleiro, A.J.; Pereira, M.A.; Alves, M.M. Carbon nanotubes accelerate methane production in pure cultures of methanogens and in a syntrophic coculture. Environ. Microbiol. 2017, 19, 2727–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vrieze, J.; Gildemyn, S.; Arends, J.B.A.; Vanwonterghem, I.; Verbeken, K.; Boon, N.; Verstraete, W.; Tyson, G.W.; Hennebel, T.; Rabaey, K. Biomass retention on electrodes rather than electrical current enhances stability in anaerobic digestion. Water Res. 2014, 54, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, P.-L.; Angenent, L.T.; Zhang, T. Extracellular electron uptake: Among autotrophs and mediated by surfaces. Trends Biotechnol. 2017, 35, 360–371. [Google Scholar] [CrossRef]
- Kang, S.; Herzberg, M.; Rodrigues, D.F.; Elimelech, M. Antibacterial effects of carbon nanotubes: Size does matter! Langmuir 2008, 24, 6409–6413. [Google Scholar] [CrossRef]
- Yadav, T.; Mungray, A.A.; Mungray, A.K. Effect of multiwalled carbon nanotubes on UASB microbial consortium. Environ. Sci. Pollut. Res. 2016, 23, 4063–4072. [Google Scholar] [CrossRef]
- Fujinawa, K.; Nagoya, M.; Kouzuma, A.; Watanabe, K. Conductive carbon nanoparticles inhibit methanogens and stabilize hydrogen production in microbial electrolysis cells. Appl. Microbiol. Biotechnol. 2019, 103, 6385–6392. [Google Scholar] [CrossRef]
- Volkman, J.K.; Rohjans, D.; Rullkötter, J.; Scholz-Böttcher, B.M.; Liebezeit, G. Sources and diagenesis of organic matter in tidal flat sediments from the German Wadden Sea. Cont. Shelf Res. 2000, 20, 1139–1158. [Google Scholar] [CrossRef]
- Ferry, J.G.; Lessner, D.J. Methanogenesis in marine sediments. Ann. N. Y. Acad. Sci. 2008, 1125, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Dias, P.; Soares, O.S.G.P.; Ramalho, P.S.F.; Pereira, M.F.R.; Alves, M.M. Synthesis, characterization and application of magnetic carbon materials as electron shuttles for the biological and chemical reduction of the azo dye Acid Orange 10. Appl. Catal. B Environ. 2017, 212, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Holliger, C.; Alves, M.; Andrade, D.; Angelidaki, I.; Astals, S.; Baier, U.; Bougrier, C.; Buffière, P.; Carballa, M.; de Wilde, V.; et al. Towards a standardization of biomethane potential tests. Water Sci. Technol. 2016, 74, 2515–2522. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.M.; Mota Vieira, J.A.; Álvares Pereira, R.M.; Pereira, M.A.; Mota, M. Effect of lipids and oleic acid on biomass development in anaerobic fixed-bed reactors. Part I: Biofilm growth and activity. Water Res. 2001, 35, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Colleran, E.; Concannon, F.; Golden, T.; Geoghegan, F.; Coates, J. Use of methanogenic activity tests to characterize anaerobic sludges, screen for anaerobic biodegradability and determine toxicity threshold against individual anaerobic trophic groups and species. Water Sci. Technol. 1992, 25, 31–40. [Google Scholar] [CrossRef]
- Coates, J.D.; Coughlan, M.F.; Colleran, E. Simple method for the measurement of the hydrogenotrophic methanogenic activity of anaerobic sludges. J. Microbiol. Methods 1996, 26, 237–246. [Google Scholar] [CrossRef]
- Stams, A.J.M.; Van Dijk, J.B.; Dijkema, C.; Plugge, C.M. Growth of syntrophic propionate-oxidizing bacteria with fumarate in the absence of methanogenic bacteria. Appl. Environ. Microbiol. 1993, 59, 1114–1119. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.; Henriques, I.; Ribeiro, D.C.; Correia, A.; Bodelier, P.L.E.; Cruz, J.V.; Brito, A.G.; Nogueira, R. Bacterial diversity and geochemical profiles in sediments from eutrophic Azorean lakes. Geomicrobiol. J. 2012, 29, 704–715. [Google Scholar] [CrossRef] [Green Version]
- APHA, AWWA, WPCF. Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA-AWWA-WEF: Washington, DC, USA, 1998. [Google Scholar]
- Romero-González, M.E.; Zambrano, E.; Mesa, J.; Ledo de Medina, H. Fractional phosphate composition in sediments from a tropical river (Catatumbo River, Venezuela). Hydrobiologia 2001, 450, 47–55. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Organic matter mineralization with the reduction of ferric iron in anaerobic sediments. Appl. Environ. Microbiol. 1986, 51, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Qiao, S.; Li, X.; Zhang, M.; Zhou, J. Nano-graphene induced positive effects on methanogenesis in anaerobic digestion. Bioresour. Technol. 2017, 224, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.R.; Martins, G.; Castro, A.R.; Pereira, L.; Alves, M.M.; Cavaleiro, A.J.; Soares, O.S.G.P.; Pereira, M.F.R. Microbial conversion of oily wastes to methane: Effect of ferric nanomaterials. In Wastes: Solutions, Treatments and Opportunities III; Vilarinho, C., Castro, F., Gonçalves, M., Fernando, A.L., Eds.; CRC Press, Taylor & Francis Group: London, UK, 2019; pp. 339–345. [Google Scholar] [CrossRef]
- Li, L.-L.; Tong, Z.-H.; Fang, C.-Y.; Chu, J.; Yu, H.-Q. Response of anaerobic granular sludge to single-wall carbon nanotube exposure. Water Res. 2015, 70, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, Y.; Holmes, D.E.; Dang, Y.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Potential enhancement of direct interspecies electron transfer for syntrophic metabolism of propionate and butyrate with biochar in up-flow anaerobic sludge blanket reactors. Bioresour. Technol. 2016, 209, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Sun, D.; Dang, Y.; Chen, H.; Zhao, Z.; Zhang, Y.; Holmes, D.E. Stimulation of methanogenesis in anaerobic digesters treating leachate from a municipal solid waste incineration plant with carbon cloth. Biores. Technol. 2016, 222, 270–276. [Google Scholar] [CrossRef]
- Pereira, R.A.; Salvador, A.F.; Dias, P.; Pereira, M.F.R.; Alves, M.M.; Pereira, L. Perspectives on carbon materials as powerful catalysts in continuous anaerobic bioreactors. Water Res. 2016, 101, 441–447. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MWCNT | SMPR (mL g−1 d−1) | |||||
|---|---|---|---|---|---|---|
| (g L−1) | Acetate | H2/CO2 | Ethanol | Butyrate (1st add.) | Butyrate (2nd add.) | Butyrate (2nd add.) t > 20 h |
| 0 | 38 ± 3 a | 394 ± 10 a | 192 ± 12 a | n.d. | 18 ± 3 a | 40 ± 0 a |
| 0.5 | 62 ± 4 b | 384 ± 33 a,b | 214 ± 4 a | n.d. | 34 ± 3 a,b | 47 ± 0 b |
| 1.0 | 82 ± 10 b | 439 ± 16 b | 235 ± 9 b | n.d. | 47 ± 3 b | 52 ± 3 c |
| MWCNT (g L−1) | Acetate | H2/CO2 | Ethanol | Butyrate (2nd add.) | Butyrate (2nd add.) t > 20 h |
|---|---|---|---|---|---|
| 0.5 | 1.6 | 1.0 | 1.1 | 1.9 | 1.2 |
| 1 | 2.1 | 1.1 | 1.2 | 2.6 | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavaleiro, A.J.; Salvador, A.F.; Martins, G.; Oliveira, C.C.; Liu, Y.; Martins, V.R.; Castro, A.R.; Soares, O.S.G.P.; Pereira, M.F.R.; Pereira, L.; et al. Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments. Appl. Sci. 2020, 10, 8184. https://doi.org/10.3390/app10228184
Cavaleiro AJ, Salvador AF, Martins G, Oliveira CC, Liu Y, Martins VR, Castro AR, Soares OSGP, Pereira MFR, Pereira L, et al. Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments. Applied Sciences. 2020; 10(22):8184. https://doi.org/10.3390/app10228184
Chicago/Turabian StyleCavaleiro, Ana J., Andreia F. Salvador, Gilberto Martins, Cláudia C. Oliveira, Yuchen Liu, Valdo R. Martins, Ana Rita Castro, Olívia Salomé G. P. Soares, Manuel Fernando R. Pereira, Luciana Pereira, and et al. 2020. "Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments" Applied Sciences 10, no. 22: 8184. https://doi.org/10.3390/app10228184
APA StyleCavaleiro, A. J., Salvador, A. F., Martins, G., Oliveira, C. C., Liu, Y., Martins, V. R., Castro, A. R., Soares, O. S. G. P., Pereira, M. F. R., Pereira, L., Langenhoff, A. A. M., Pereira, M. A., & Alves, M. M. (2020). Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments. Applied Sciences, 10(22), 8184. https://doi.org/10.3390/app10228184