Influence of Macrophyte and Gut Microbiota on Mercury Contamination in Fish: A Microcosms Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling Procedure
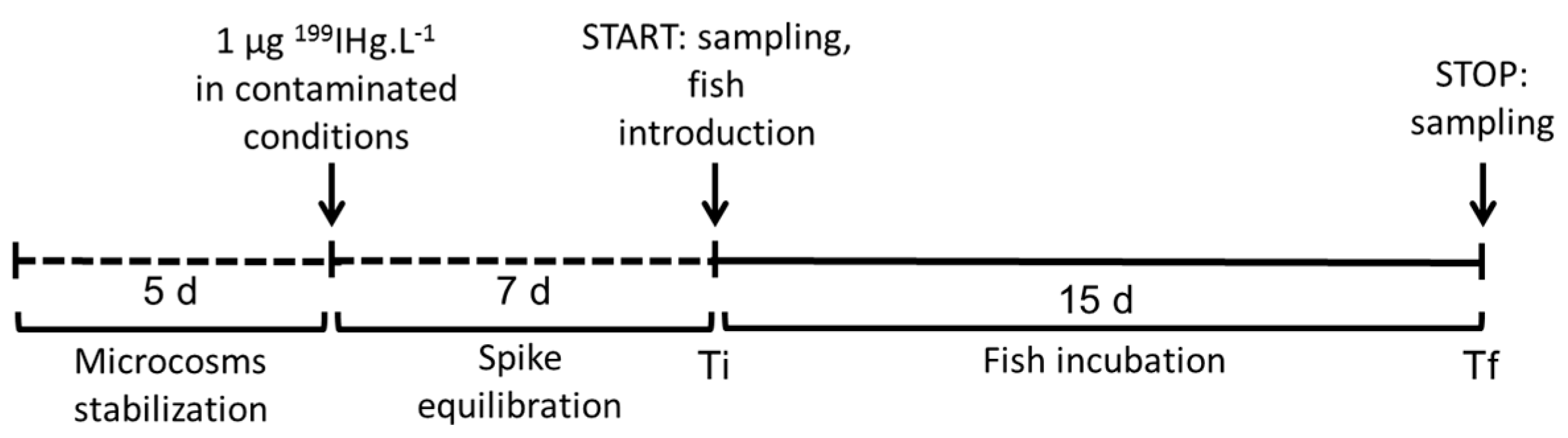
2.2. Experimental Design
2.3. Total Mercury Analysis
2.4. GC-ICP-MS(Gas Chromatography-Inductively Coupled Plasma-Mass Spectrometry) Analysis
2.5. MiSEQ Analysis
2.6. Sulfate-Reducing Prokaryotes (SRP) Culture and Identification
2.7. Hg Methylation Capacity by Isolated Strains
2.8. Statistical Analysis
3. Results
3.1. Organisms’ Health
3.2. Mercury Speciation in Control Units
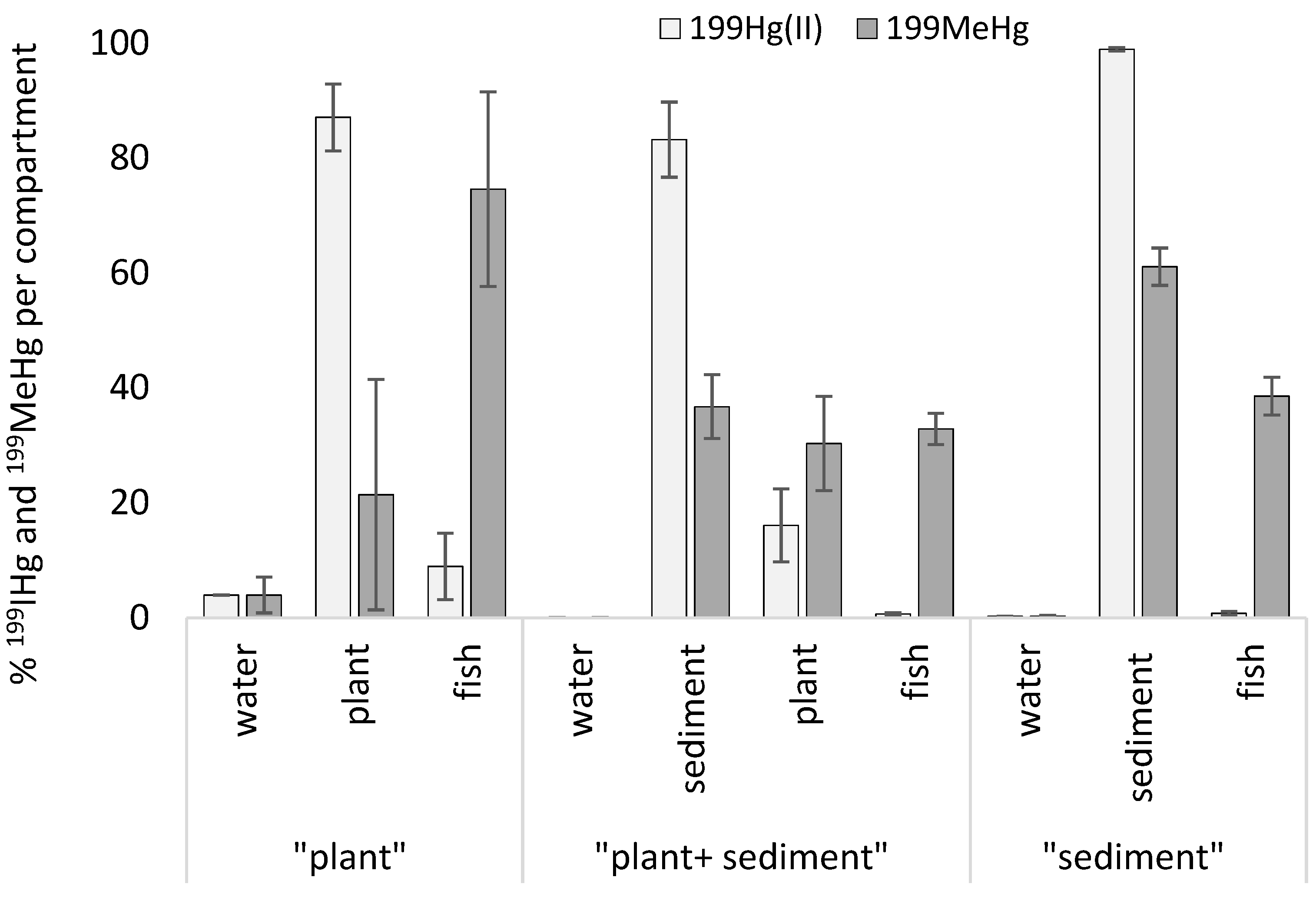
3.3. Distribution and Transformation of Isotopic Tracer 199Hg in Contaminated Units
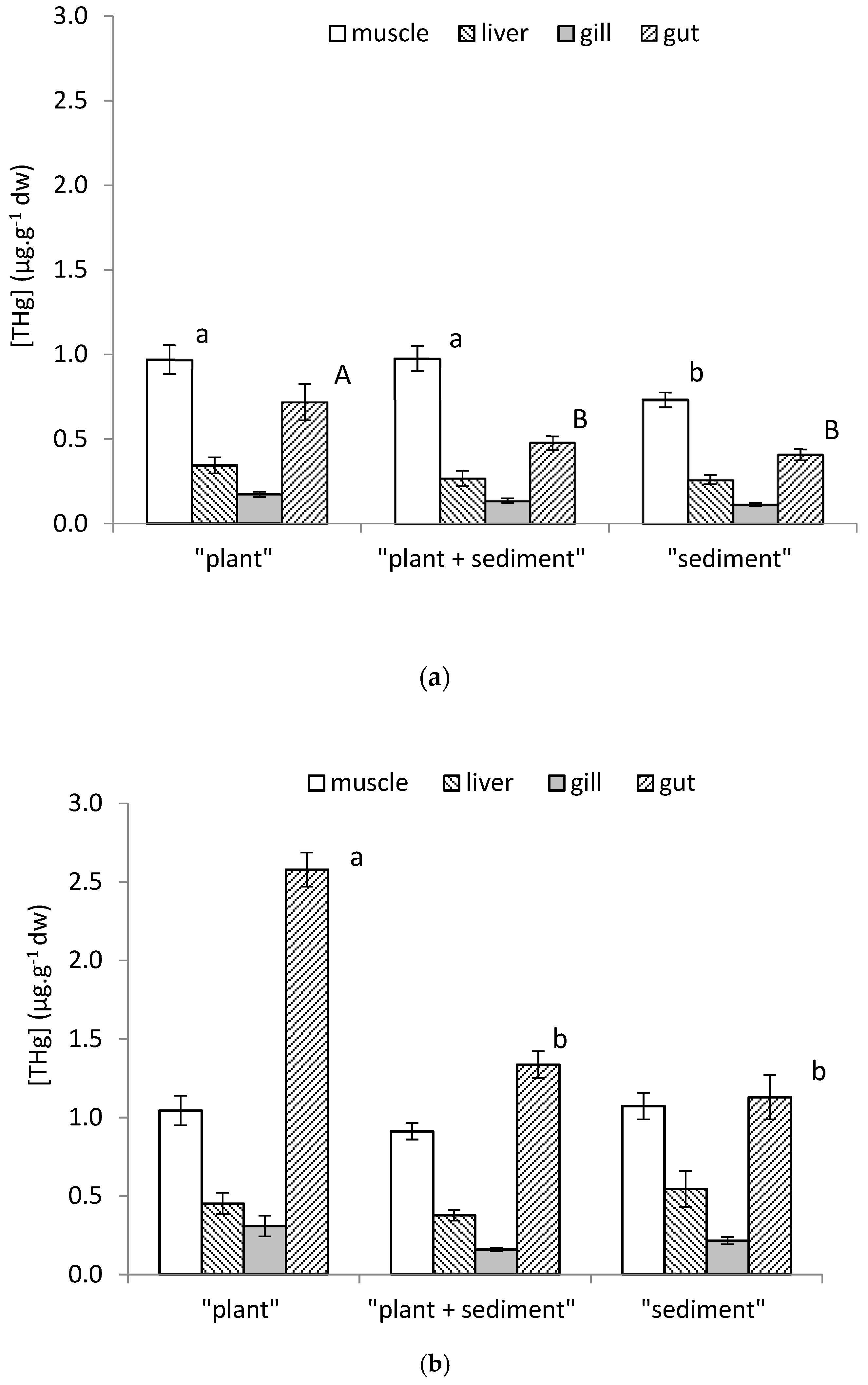
3.4. Organostropism of THg in Control and Contaminated Fish
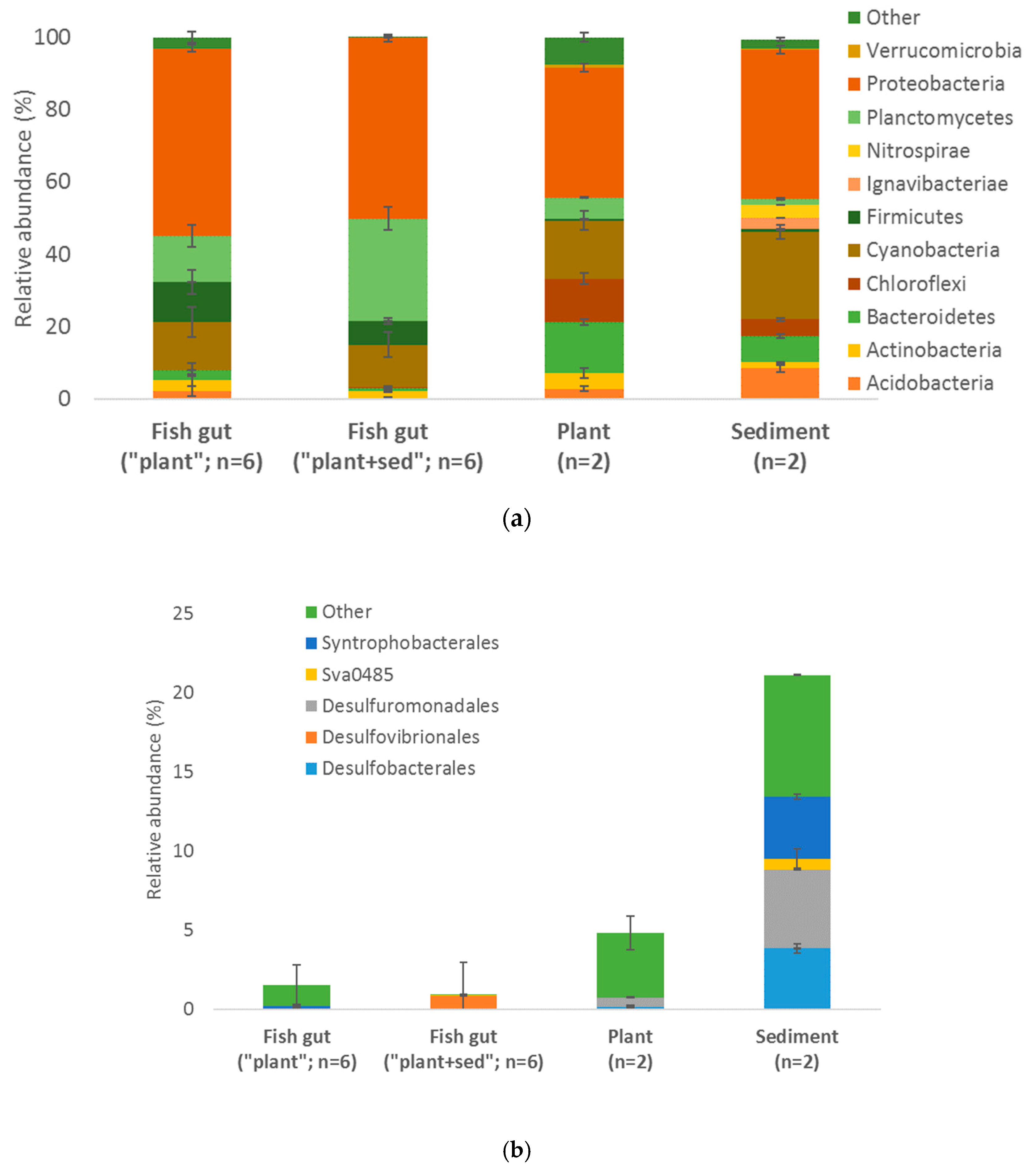
3.5. Microorganisms Global Diversity
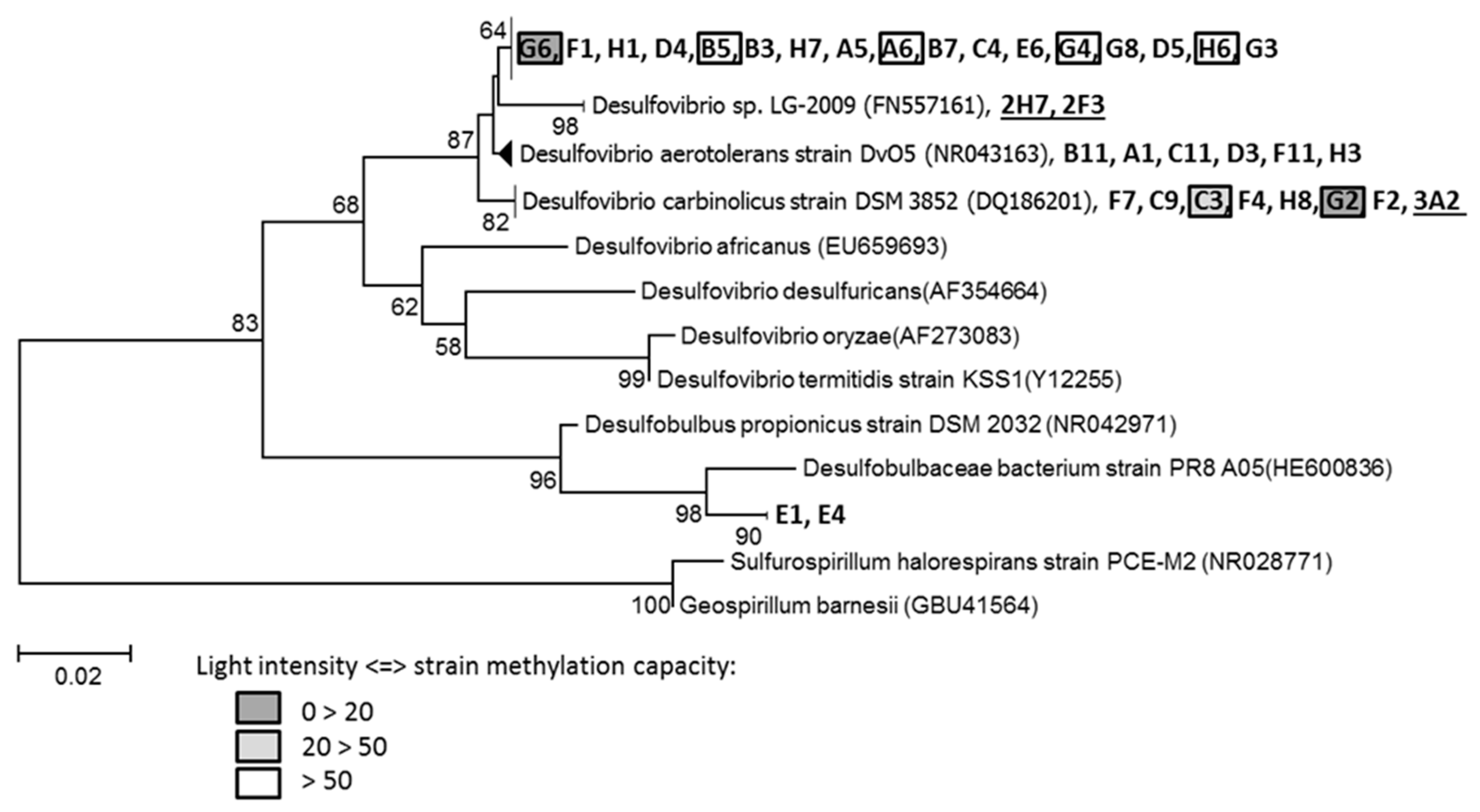
3.6. Sulfate Reducers Isolation and Hg Methylation Capacity
4. Discussion
4.1. Periphyton Associated with Root’s Macrophytes: Main Source of MeHg for Organisms
4.1.1. High Methylation Potential in Aquatic Rhizosphere of Invasive Macrophyte
4.1.2. MeHg Produced in Aquatic Rhizosphere is Bioavailable for Organisms
4.2. Involvement of Fish Gut Microbiota in Hg Methylation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Guimaraes, J.R.D.; Meili, M.; Hylander, L.D.; Castro e Silva, E.; Roulet, M.; Mauro, J.B.N.; Lemos, R.A. Net mercury methylation in five tropical flood plain regions of Brazil, high in the root zone of floating macrophyte mats but low in surface sediments and flooded soils. Sci. Total Environ. 2000, 261, 99–107. [Google Scholar] [CrossRef]
- Mauro, J.B.; Guimaraes, J.R.; Hintelmann, H.; Watras, C.J.; Haack, E.A.; Coelho-Souza, S.A. Mercury methylation in macrophytes, periphyton, and water, comparative studies with stable and radio-mercury additions. Anal. Bioanal. Chem. 2002, 374, 983–989. [Google Scholar]
- Achá, D.; Iñiguez, V.; Roulet, M.; Guimarães, J.R.D.; Luna, R.; Alanoca, L.; Sanchez, S. Sulfate-reducing bacteria in floating macrophyte rhizospheres from an Amazonian floodplain lake in Bolivia and their association with Hg methylation. Appl. Environ. Microbiol. 2005, 71, 7531–7535. [Google Scholar] [CrossRef] [Green Version]
- Achá, D.; Hintelmann, H.; Yee, J. Importance of sulfate reducing bacteria in mercury methylation and demethylation in periphyton from Bolivian Amazon Region. Chemosphere 2011, 82, 911–916. [Google Scholar] [CrossRef]
- Correia, R.R.S.; Miranda, M.R.; Guimaraes, J.R.D. Mercury methylation and the microbial consortium in periphyton of tropical macrophytes, Effect of different inhibitors. Environ. Res. 2012, 112, 86–91. [Google Scholar] [CrossRef]
- Desrosiers, M.; Planas, D.; Mucci, A. Mercury methylation in the epilithon of boreal shield aquatic ecosystems. Environ. Sci. Technol. 2006, 40, 1540–1546. [Google Scholar] [CrossRef]
- Gentès, S.; Monperrus, M.; Legeay, A.; Maury-Brachet, R.; Davail, S.; André, J.M.; Guyoneaud, R. Incidence of invasive macrophytes on methylmercury budget in temperate lakes, Central role of bacterial periphytic communities. Environ. Pollut. 2013, 172, 116–123. [Google Scholar] [CrossRef]
- Gentès, S.; Taupiac, J.; Colin, Y.; André, J.M.; Guyoneaud, R. Bacterial periphytic communities related to mercury methylation within aquatic plant roots from a temperate freshwater lake (South-Western France). Environ. Sci. Pollut. Res. 2017, 24, 19223–19233. [Google Scholar] [CrossRef]
- Flemming, E.J.; Mack, E.E.; Green, P.G.; Nelson, D.C. Mercury methylation from unexpected sources, Molybdate-inhibited freshwater sediments and an iron-reducing bacterium. Appl. Environ. Microbiol. 2006, 72, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Kerin, E.J.; Gilmour, C.C.; Roden, E.; Suzuki, M.T.; Coates, J.D.; Mason, R.P. Mercury methylation by dissimilatory iron-reducing bacteria. Appl. Environ. Microbiol. 2006, 72, 7919–7921. [Google Scholar] [CrossRef] [Green Version]
- Hamelin, S.; Amyot, M.; Barkay, T.; Wang, Y.; Planas, D. Methanogens, Principal methylators of mercury in lake periphyton. Environ. Sci. Technol. 2011, 45, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, C.C.; Podar, M.; Bullock, A.L.; Graham, A.M.; Brown, S.D.; Somenahally, A.C.; Elias, D. Mercury methylation by novel microorganisms from new environments. Environ. Sci. Technol. 2013, 47, 11810–11820. [Google Scholar] [CrossRef] [PubMed]
- Parks, J.M.; Johs, A.; Podar, M.; Bridou, R.; Hurt, R.A., Jr.; Smith, S.D.; Tomanicek, S.J.; Qian, Y.; Brown, S.D.; Brandt, C.C.; et al. The genetic basis for bacterial mercury methylation. Science 2013, 339, 1332–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podar, M.; Gilmour, C.C.; Brandt, C.C.; Soren, A.; Brown, S.D.; Crable, B.R.; Palumbo, A.V.; Somenahally, A.C.; Elias, D.A. Global prevalence and distribution of genes and microorganisms involved in mercury methylation. Sci. Adv. 2015, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Goñi-Urriza, M.; Corsellis, Y.; Lanceleur, L.; Tessier, E.; Gury, J.; Monperrus, M.; Guyoneaud, R. Relationships between bacterial energetic metabolism, mercury methylation potential, and hgcA/hgcB gene expression in Desulfovibrio dechloroacetivorans BerOc1. Environ. Sci. Pollut. Res. 2015, 22, 13764–13771. [Google Scholar] [CrossRef]
- Bravo, A.G.; Loizeau, J.L.; Dranguet, P.; Makri, S.; Björn, E.; Ungureanu, V.G.; Cosio, C. Persistent Hg contamination and occurrence of Hg-methylating transcript (hgcA) downstream of a chlor-alkali plant in the Olt River (Romania). Environ. Sci. Pollut. Res. 2016, 23, 10529–10541. [Google Scholar] [CrossRef] [Green Version]
- Guasch, H.; Admiraal, W.; Sabater, S. Contrasting effects of organic and inorganic toxicants on freshwater periphyton. Aquat. Toxicol. 2003, 64, 165–175. [Google Scholar] [CrossRef]
- King, J.K.; Harmon, S.M.; Fu, T.T.; Gladden, J.B. Mercury removal, methylmercury formation, and sulfate-reducing bacteria profiles in wetland mesocosms. Chemosphere 2002, 46, 859–870. [Google Scholar] [CrossRef]
- Göthberg, A.; Greger, M. Formation of methyl mercury in an aquatic macrophyte. Chemosphere 2006, 65, 2096–2105. [Google Scholar]
- Roulet, M.; Lucotte, M.; Guimarães, J.R.; Rheault, I. Methylmercury in water, seston, and epiphyton of an Amazonian river and its floodplain, Tapajós River, Brazil. Sci. Total Environ. 2000, 261, 43–59. [Google Scholar] [CrossRef]
- Molina, C.I.; Gibon, F.M.; Duprey, J.L.; Dominguez, E.; Guimaraes, J.R.; Roulet, M. Transfer of mercury and methylmercury along macroinvertebrate food chains in a floodplain lake of the Beni River, Bolivian Amazonia. Sci. Total Environ. 2010, 408, 3382–3391. [Google Scholar] [CrossRef] [PubMed]
- Leite, R.G.; Araújo-Lima, C.A.R.M.; Victoria, R.L.; Martinelli, L.A. Stable isotope analysis of energy sources for larvae of eight fish species from the Amazon floodplain. Ecol. Freshw. Fish. 2002, 11, 56–63. [Google Scholar] [CrossRef]
- Dutartre, A. De la régulation des plantes aquatiques envahissantes à la gestion des hydrosystèmes. Ingénierie Ecol. 2004, 87–100. [Google Scholar]
- Lambert, E.; Dutartre, A.; Coudreuse, J.; Haury, J. Relationships between the biomass production of invasive Ludwigia species and physical properties of habitats in France. Hydrobiologia 2010, 656, 173–186. [Google Scholar] [CrossRef]
- Hintelmann, H.; Keppel-Jones, K.; Evans, R.D. Constants of mercury methylation and demethylation rates in sediments and comparison of tracer and ambient mercury availability. Environ. Toxicol. Chem. 2000, 19, 2204–2211. [Google Scholar] [CrossRef]
- Monperrus, M.; Tessier, E.; Amouroux, D.; Leynaert, A.; Huonnic, P.; Donard, O.F.X. Mercury methylation, demethylation and reduction rates in coastal and marine surface waters of the Mediterranean Sea. Mar. Chem. 2007, 107, 49–63. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Biddinger, S.B. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Dietert, R.R.; Silbergeld, E.K. Biomarkers for the 21st century, listening to the microbiome. Toxicol. Sci. 2015, 144, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Feng, X.B.; Wang, W.X. In vivo mercury methylation and demethylation in freshwater tilapia quantified by mercury stable isotopes. Environ. Sci. Technol. 2013, 47, 7949–7957. [Google Scholar] [CrossRef]
- Rudd, J.W.; Furutani, A.; Turner, M.A. Mercury methylation by fish intestinal contents. Appl. Environ. Microbiol. 1980, 40, 777–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colin, Y.; Gury, J.; Monperrus, M.; Gentès, S.; Ayala, P.; Goñi-Urriza, M.; Guyoneaud, R. Biosensor for screening bacterial mercury methylation, example within the Desulfobulbaceae. Res. Microbiol. 2018, 169, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Ciutat, A.; Boudou, A. Bioturbation effects on cadmium and zinc transfers from contaminated sediment and on metal bioavailability of benthic bivalves. Environ. Toxicol. Chem. 2003, 22, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Canredon, A.; Anschutz, P.; Buquet, D.; Charbonnier, C.; Gentès, S.; Legeay, A.; Feurtet-Mazel, A.; Poirier, D.; Bujan, S.; Devaux, L.; et al. Sulphate and organic matter control on mercury biogeochemistry and production of methylmercury in Aquitaine coastal lakes. Appl. Geochem. 2019, 104, 135–145. [Google Scholar] [CrossRef]
- Rodriguez Martin-Doimeadios, R.C.; Krupp, E.; Amouroux, D.; Donard, O.F.X. Application of isotopically labeled methylmercury for isotope dilution analysis of biological samples using gas chromatography/ICPMS. Anal. Chem. 2002, 74, 2505–2512. [Google Scholar] [CrossRef]
- Clémens, S.; Monperrus, M.; Donard, O.F.; Amouroux, D.; Guérin, T. Mercury speciation analysis in seafood by species-specific isotope dilution, method validation and occurrence data. Anal. Bioanal. Chem. 2011, 401, 2699. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, P.; Monperrus, M.; Alonso, J.I.G.; Amouroux, D.; Donard, O.F.X. Comparison of different numerical approaches for multiple spiking species-specific isotope dilution analysis exemplified by the determination of butyltin species in sediments. J. Anal. At. Spectrom. 2007, 22, 1373–1382. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS, Find; Rapidly; OTUs with Galaxy Solution. Bioinformatics 2017, 34, 1287–1294. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm, robust and fast clustering method for amplicon-based studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2012, 10, 57–59. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project, improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Colin, Y.; Goñi-Urriza, M.; Caumette, P.; Guyoneaud, R. Combination of high throughput cultivation and dsrA sequencing for assessment of sulfate-reducing bacteria diversity in sediments. FEMS Microbiol. Ecol. 2012, 83, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4, Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbour-joining method, a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Ivask, A.; Hakkila, K.; Virta, M. Detection of organomercurials with sensor bacteria. Anal. Chem. 2001, 73, 5168–5171. [Google Scholar] [CrossRef]
- Ranchou-Peyruse, M.; Monperrus, M.; Bridou, R.; Duran, R.; Amouroux, D.; Salvado, J.C.; Guyoneaud, R. Overview of mercury methylation capacities among anaerobic bacteria including representatives of the sulphate-reducers, Implications for environmental studies. Geomicrobiol. J. 2009, 26, 1–8. [Google Scholar] [CrossRef]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Photodecomposition of methylmercury in an arctic Alaskan lake. Environ. Sci. Technol. 2006, 40, 1212–1216. [Google Scholar] [CrossRef]
- Engstrom, D.R.; Balogh, S.J.; Swain, E.B. History of mercury inputs to Minnesota lakes, influences of watershed disturbance and localized atmospheric deposition. Limnol. Oceanogr. 2007, 52, 2467–2683. [Google Scholar] [CrossRef]
- Muir, D.; Wang, X.; Yang, F.; Nguyen, N.; Jackson, T.A.; Evans, M.S.; Douglas, M.; Köck, G.; Lamoureux, S.F.; Pienitz, R.; et al. Spatial trends and historical deposition of mercury in eastern and northern Canada inferred from lake sediment cores. Environ. Sci. Technol. 2009, 43, 4802–4809. [Google Scholar] [CrossRef]
- Christensen, P.B.; Revsbech, N.P.; Sand-Jensen, K. Microsensor analysis of oxygen in the rhizosphere of the aquatic macrophyte littorella uniflora. Plant. Physiol. 1994, 105, 847–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gemerden, H. Microbial mats, a joint-venture. Mar. Geol 1993, 113, 3–25. [Google Scholar] [CrossRef]
- Hamelin, S.; Planas, D.; Amyot, M. Mercury methylation and demethylation by periphyton biofilms and their host in a fluvial wetland of the St. Lawrence River (QC, Canada). Sci. Total Environ. 2015, 512–513, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.Q.; Adatto, I.; Montesdeoca, M.R.; Driscoll, C.T.; Hines, M.E.; Barkay, T. Mercury methylation in Sphagnum moss mats and its association with sulfate-reducing bacteria in an acidic Adirondack forest lake wetland. FEMS Microbiol. Ecol. 2010, 74, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.D.; Bodaly, R.A.; Fudge, R.J.P.; Rudd, J.W.M.; Rosenberg, D.M. Food as the dominant pathway of methylmercury uptake by fish. Water Air Soil Pollut. 1997, 100, 13–24. [Google Scholar]
- Harris, R.; Bodaly, R.A. Temperature, growth and dietary effects on fish mercury dynamics in two Ontario lakes. Biogeochemistry 1998, 40, 175–187. [Google Scholar] [CrossRef]
- Pickhardt, P.C.; Stepanova, M.; Fisher, N.S. Contrasting uptake routes and tissue distributions of inorganic and methylmercury in mosquitofish (Gambusia affinis) and redear sunfish (Lepomis microlophus). Environ. Toxicol. Chem. 2006, 25, 2132–2142. [Google Scholar] [CrossRef]
- Junk, W.J.; Piedade, M.T.F. Plant life in the floodplain with special reference to herbaceous plants. In The Central Amazon Floodplain; Springer: Berlin, Germany, 1997; pp. 147–185. [Google Scholar]
- Gentès, S.; Maury-Brachet, R.; Guyoneaud, R.; Monperrus, M.; André, J.M.; Davail, S.; Legeay, A. Mercury bioaccumulation along food webs in temperate aquatic ecosystems colonized by aquatic macrophytes in south western France. Ecotoxicol. Environ. Saf. 2013, 91, 180–187. [Google Scholar] [CrossRef]
- Maury-Brachet, R.; Durrieu, G.; Dominique, Y.; Boudou, A. Mercury distribution in fish organs and food regimes, significant relationships from twelve species collected in French Guiana (Amazonian basin). Sci. Total Environ. 2006, 368, 262–270. [Google Scholar]
- Silva, L.F.F.; Machado, W.; Filho, S.D.L.; Lacerda, L.D. Mercury accumulation in sediments of a mangrove ecosystem in SE Brazil. Water Air Soil Pollut. 2003, 145, 67–77. [Google Scholar] [CrossRef]
- Kaschak, E.; Knopf, B.; Petersen, J.H.; Bings, N.H.; König, H. Biotic methylation of mercury by intestinal and sulfate-reducing bacteria and their potential role in mercury accumulation in the tissue of the soil-living Eisenia foetida. Soil Biol. Biochem. 2014, 69, 202–211. [Google Scholar] [CrossRef]
- Martín-Doimeadios, R.R.; Mateo, R.; Jiménez-Moreno, M. Is gastrointestinal microbiota relevant for endogenous mercury methylation in terrestrial animals? Environ. Res. 2017, 152, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, F.; Wang, W. In vivo mercury demethylation in a marine fish (Acanthopagrus schlegeli). Environ. Sci. Technol. 2017, 51, 6441–6451. [Google Scholar] [CrossRef] [PubMed]
- Boudou, A.; Ribeyre, F. Experimental study of trophic contamination of Salmogairdneri by two mercury compounds—HgCl2 and CH3HgCl—Analysis at the organism and organ levels. Water Air Soil Pollut. 1985, 26, 137–148. [Google Scholar] [CrossRef]
- Ribeiro, O.C.A.; Belger, L.; Pelletier, E.; Rouleau, C. Histopathological evidence of inorganic mercury and methyl mercury toxicity in the arctic charr (Salvelinus alpinus). Environ. Res. 2002, 90, 217–225. [Google Scholar] [CrossRef]
- Berntssen, M.H.G.; Hylland, K.; Julshamn, K.; Lundebye, A.K.; Waagbo, R. Maximum limits of organic and inorganic mercury in fish feed. Aquacult. Nutr. 2004, 10, 83–97. [Google Scholar] [CrossRef]
- Hoyle, I.; Handy, R.D. Dose-dependent inorganic mercury absorption by isolated perfused intestine of rainbow trout, Oncorhynchus mykiss, involves both amiloride-sensitive and energy-dependent pathways. Aquat. Toxicol. 2005, 72, 147–159. [Google Scholar] [CrossRef]
- Pak, K.; Bartha, R. Products of mercury demethylation by sulfidogens and methanogens. Bull. Environ. Contam. Toxicol. 1998, 61, 690–694. [Google Scholar] [CrossRef]
- Bridou, R.; Monperrus, M.; Rodriguez Gonzalez, P.; Guyoneaud, R.; Amouroux, D. Simultaneous determination of mercury methylation and demethylation capacities of various sulfate reducing bacteria using species-specific isotopic tracers. Environ. Toxicol. Chem. 2011, 30, 337–344. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Matrix | [IHg] | [MeHg] | % MeHg | n |
|---|---|---|---|---|---|
| Plant-C | Water | 1.1 ± 0.3 | 0.1 ± 0.1 | 7.7 ± 3.8 | 4 |
| Plant | 38.9 ± 3.7 | 6.2 ± 3.6 | 11.3 ± 5.1 | 4 | |
| Fish | 345.2 ± 127.4 | 1152.6 ± 136.4 | 77.3 ± 7.9 | 6 | |
| Plant+sediment-C | Water | 0.8 ± 0.1 | 0.1 ± 0 | 13.4 ± 1.3 | 6 |
| Sediment | 5.7 ± 1.2 | 0.1 ± 0 | 1.6 ± 0.8 | 4 | |
| Plant | 24.7 ± 7.3 | 1.4 ± 0.4 | 5.6 ± 0.5 | 6 | |
| Fish | 220.3 ± 24.4 | 1083.2 ± 68.8 | 83.2 ± 1.3 | 9 | |
| Sediment-C | Water | 0.9 ± 0.2 | 0.1 ± 0 | 21.3 ± 10 | 6 |
| Sediment | 5 ± 0.4 | 0.1 ± 0 | 1.1 ± 0.2 | 5 | |
| Fish | 174.5 ± 15 | 948.1 ± 63.9 | 84.3 ± 1.2 | 9 |
| Condition | Matrix | [199IHg] | [199MMHg] | % Methylation | n | |
|---|---|---|---|---|---|---|
| Ti | Plant-Hg | Water | 109.2–41.8 | 24.1–9.2 | 18.1–18.1 | 2 |
| Plant | 2061.1–2572.8 | 311.4–22.9 | 13.1–0.9 | 2 | ||
| Plant+sediment-Hg | Water | 41 ± 15.4 | 0.8 ± 0.7 | 3.7 ± 3.4 | 3 | |
| Sediment | 2.9 ± 1.5 | 0.6 ± 0.3 | 13.9 ± 6.1 | 3 | ||
| Plant | 1362 ± 272.2 | 92.8 ± 36 | 6.2 ± 2.4 | 3 | ||
| Sediment-Hg | Water | 244.2 ± 27.7 | 0.2 ± 0 | 0.1 ± 0 | 3 | |
| Sediment | 1.8 ± 0.1 | 0 ± 0 | 2.3 ± 1.6 | 3 | ||
| Tf | Plant-Hg | Water | 10.9–9.5 | 8–0.6 | 42.4 - 6.4 | 2 |
| Plant | 587.2–397.1 | 55.8 | 12.3 | 2-1 | ||
| Fish | 63.7 ± 25.1 | 180.7 ± 69.1 | 77.3 ± 4.9 | 6 | ||
| Plant+sediment-Hg | Water | 0.4 ± 0.1 | 0 ± 0 | 13 ± 6 | 3 | |
| Sediment | 3.4 ± 0.3 | 0.3 ± 0.1 | 8.2 ± 3.2 | 3 | ||
| Plant | 44.4 ± 4 | 16.8 ± 2.3 | 27.4 ± 0.9 | 3 | ||
| Fish | 15 ± 3.1 | 153.9 ± 39.7 | 90 ± 1.4 | 7 | ||
| Sediment-Hg | Water | 3.9 ± 1.7 | 0.5 ± 0.2 | 12.9 ± 5.9 | 3 | |
| Sediment | 5.3 ± 0.2 | 0.5 ± 0 | 7.9 ± 1.1 | 3 | ||
| Fish | 26.8 ± 5.1 | 172.5 ± 14 | 89.3 ± 1.7 | 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentès, S.; Monperrus, M.; Legeay, A.; Gassie, C.; Maury-Brachet, R.; André, J.-M.; Guyoneaud, R. Influence of Macrophyte and Gut Microbiota on Mercury Contamination in Fish: A Microcosms Study. Appl. Sci. 2020, 10, 1500. https://doi.org/10.3390/app10041500
Gentès S, Monperrus M, Legeay A, Gassie C, Maury-Brachet R, André J-M, Guyoneaud R. Influence of Macrophyte and Gut Microbiota on Mercury Contamination in Fish: A Microcosms Study. Applied Sciences. 2020; 10(4):1500. https://doi.org/10.3390/app10041500
Chicago/Turabian StyleGentès, Sophie, Mathilde Monperrus, Alexia Legeay, Claire Gassie, Régine Maury-Brachet, Jean-Marc André, and Rémy Guyoneaud. 2020. "Influence of Macrophyte and Gut Microbiota on Mercury Contamination in Fish: A Microcosms Study" Applied Sciences 10, no. 4: 1500. https://doi.org/10.3390/app10041500
APA StyleGentès, S., Monperrus, M., Legeay, A., Gassie, C., Maury-Brachet, R., André, J. -M., & Guyoneaud, R. (2020). Influence of Macrophyte and Gut Microbiota on Mercury Contamination in Fish: A Microcosms Study. Applied Sciences, 10(4), 1500. https://doi.org/10.3390/app10041500