Secreted Protein Acidic and Rich in Cysteine: Metabolic and Homeostatic Properties beyond the Extracellular Matrix Structure



{kind=link}
{kind=link}
{kind=link}
Abstract
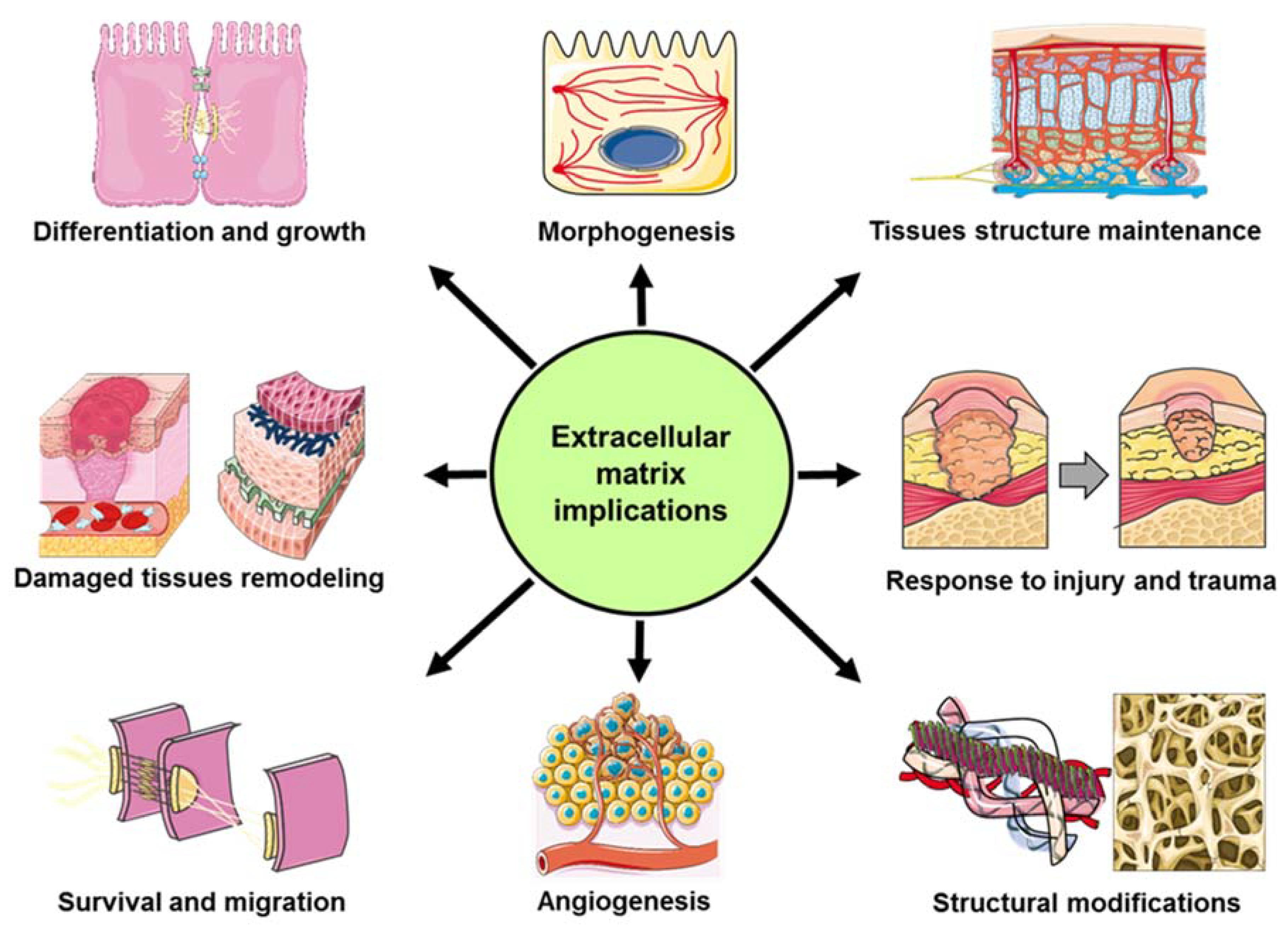
:1. Extracellular Matrix (ECM) and Secreted Protein Acidic and Rich in Cysteine (SPARC)
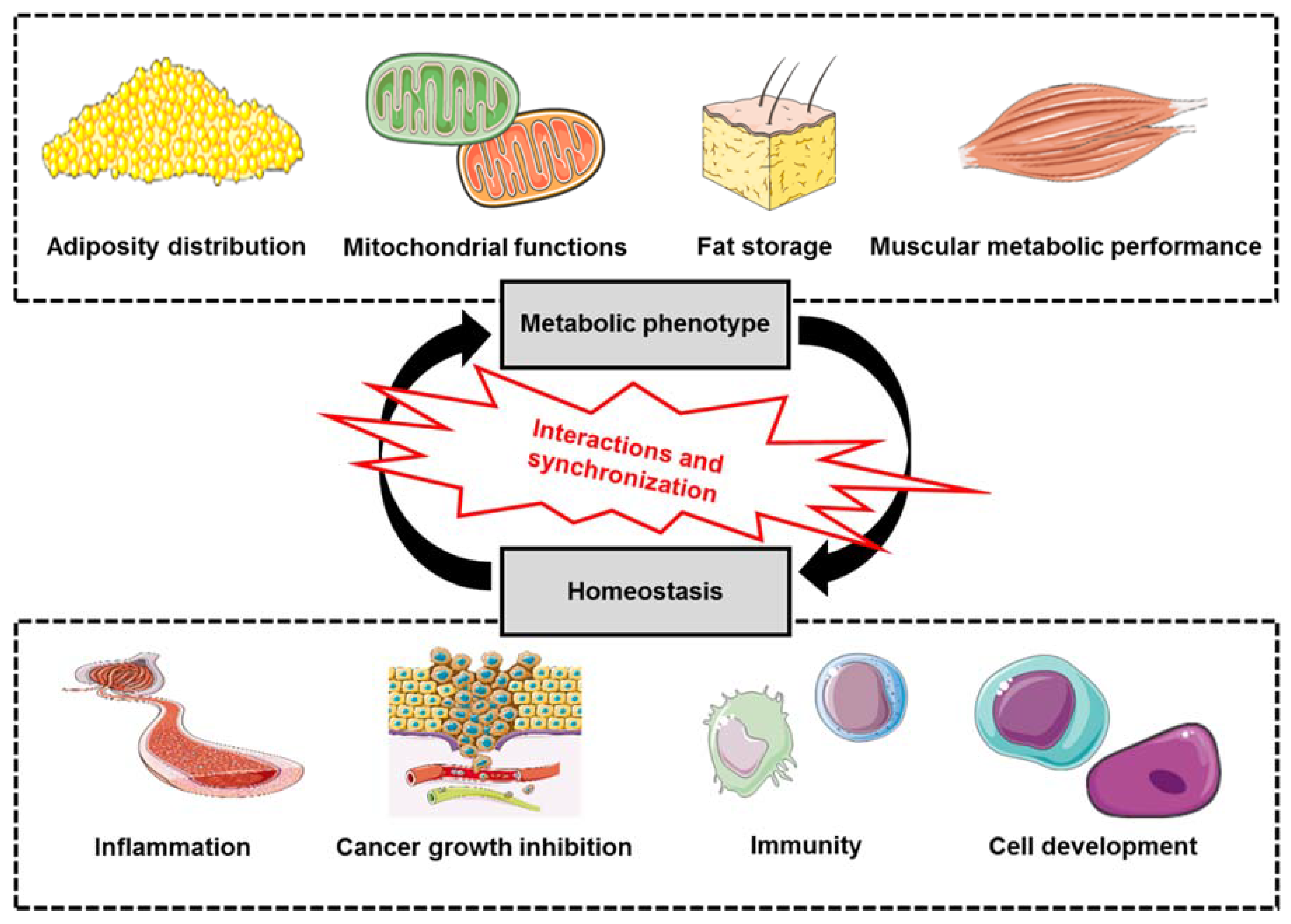
2. SPARC: Metabolics and Homeostasis
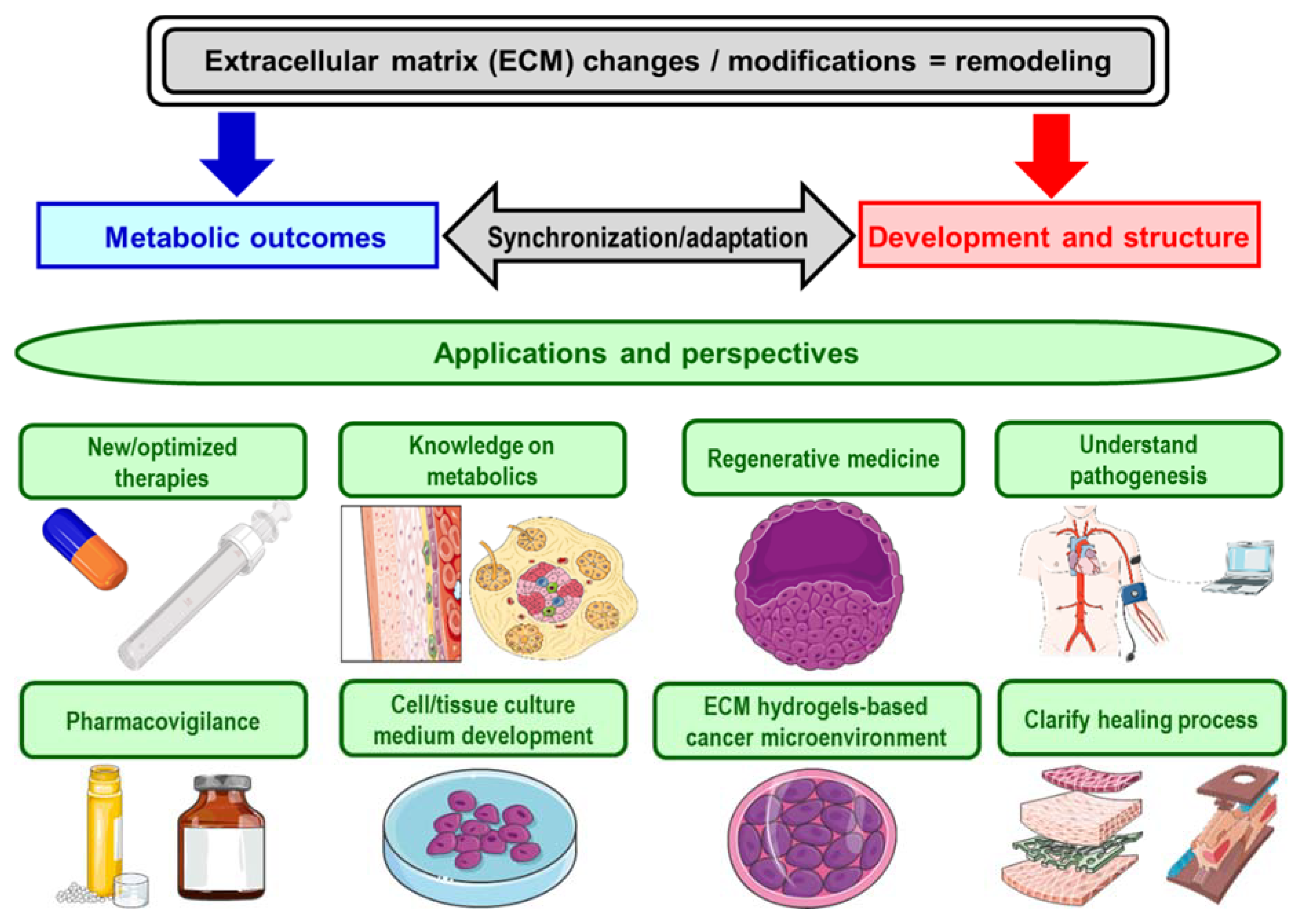
3. Perspective and Implications
Funding
Acknowledgments
Conflicts of Interest
References
- Riazi, A.M.; Kwon, S.Y.; Stanford, W.L. Stem Cell Sources for Regenerative Medicine. Adv. Struct. Saf. Stud. 2009, 482, 55–90. [Google Scholar]
- Mizuno, H. Adipose-derived stem and stromal cells for cell-based therapy: Current status of preclinical studies and clinical trials. Curr. Opin. Mol. Ther. 2010, 12, 442–449. [Google Scholar]
- Aboushwareb, T.; Atala, A. Stem cells in urology. Nat. Clin. Pract. Urol. 2008, 5, 621–631. [Google Scholar] [CrossRef]
- Horton, E.R.; Vallmajo-Martin, Q.; Martin, I.; Snedeker, J.G.; Ehrbar, M.; Blache, U. Extracellular Matrix Production by Mesenchymal Stromal Cells in Hydrogels Facilitates Cell Spreading and Is Inhibited by FGF-2. Adv. Heal. Mater. 2020, 1901669. [Google Scholar] [CrossRef] [PubMed]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldin, L.T.; Cramer, M.C.; Velankar, S.S.; White, L.; Badylak, S.F. Extracellular matrix hydrogels from decellularized tissues: Structure and function. Acta Biomater. 2017, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swinehart, I.T.; Badylak, S.F. Extracellular matrix bioscaffolds in tissue remodeling and morphogenesis. Dev. Dyn. 2016, 245, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Vigetti, D.; Viola, M.; Karousou, E.; Deleonibus, S.; Karamanou, K.; De Luca, G.; Passi, A. Epigenetics in extracellular matrix remodeling and hyaluronan metabolism. FEBS J. 2014, 281, 4980–4992. [Google Scholar] [CrossRef]
- Sonbol, H.S. Extracellular Matrix Remodeling in Human Disease. J. Microsc. Ultrastruct. 2018, 6, 123–128. [Google Scholar] [CrossRef]
- Rauch, U. Extracellular matrix components associated with remodeling processes in brain. Cell. Mol. Life Sci. 2004, 61, 2031–2045. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Lewis, D.; Gerecht, S. Bioinspired Hydrogels to Engineer Cancer Microenvironments. Annu. Rev. Biomed. Eng. 2017, 19, 109–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Termine, J.D.; Kleinman, H.K.; Whitson, S.; Conn, K.M.; McGarvey, M.L.; Martin, G.R. Osteonectin, a bone-specific protein linking mineral to collagen. Cell 1981, 26, 99–105. [Google Scholar] [CrossRef]
- Bradshaw, A.D. The role of SPARC in extracellular matrix assembly. J. Cell Commun. Signal. 2009, 3, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanemi, A.; Melouane, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and bioenergetics: Extracellular matrix, adipocytes remodeling and skeletal muscle metabolism. Int. J. Biochem. Cell Boil. 2019, 117, 105627. [Google Scholar] [CrossRef]
- Kelleher, C.M.; McLean, S.; Mecham, R.P. Vascular Extracellular Matrix and Aortic Development, in Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2004; pp. 153–188. [Google Scholar]
- Maeda, H.; Wada, N.; Tomokiyo, A.; Monnouchi, S.; Akamine, A. Chapter Six-Prospective Potency of TGF-β1 on Maintenance and Regeneration of Periodontal Tissue, in International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 283–367. [Google Scholar]
- McKee, D.M.; Cole, W.G. Chapter 2-Bone Matrix and Mineralization, in Pediatric Bone, 2nd ed.; Glorieux, F.H., Pettifor, J.M., Jüppner, H., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 9–37. [Google Scholar]
- Workman, G.; Bradshaw, A.D. Chapter 19-Production and Purification of Recombinant Human SPARC, in Methods in Cell Biology; Mecham, R.P., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 335–345. [Google Scholar]
- Humphrey, E.L.; Morris, G.E.; Fuller, H. Valproate reduces collagen and osteonectin in cultured bone cells. Epilepsy Res. 2013, 106, 446–450. [Google Scholar] [CrossRef]
- Cerilli, A.L.; Wick, M.R. Chapter 3-Immunohistology of Soft Tissue and Osseous Neoplasms, in Diagnostic Immunohistochemistry, 2nd ed.; Dabbs, D.J., Ed.; Churchill Livingstone: London, UK, 2006; pp. 65–120. [Google Scholar]
- Heinegard, D.; Oldberg, A. Structure and biology of cartilage and bone matrix noncollagenous macromolecules. Faseb J. 1989, 3, 2042–2051. [Google Scholar] [CrossRef] [Green Version]
- Jundt, G.; Termine, J.D.; Schulz, A. Osteonectin--a differentiation marker of bone cells. Cell Tissue Res. 1987, 248, 409–415. [Google Scholar] [CrossRef]
- Badylak, S.F. The extracellular matrix as a scaffold for tissue reconstruction. Semin. Cell Dev. Boil. 2002, 13, 377–383. [Google Scholar] [CrossRef]
- Hodde, J.P. Extracellular matrix as a bioactive material for soft tissue reconstruction. ANZ J. Surg. 2006, 76, 1096–1100. [Google Scholar] [CrossRef]
- Spang, M.T.; Karen, C. Extracellular matrix hydrogel therapies: In vivo applications and development. Acta Biomater. 2018, 68, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D.; Sage, E.H. SPARC, a matricellular protein that functions in cellular differentiation and tissue response to injury. J. Clin. Investig. 2001, 107, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brekken, R.A.; Sage, E. SPARC, a matricellular protein: At the crossroads of cell–matrix communication. Matrix Boil. 2001, 19, 815–827. [Google Scholar] [CrossRef]
- Bella, J.; Hulmes, D.J. Fibrillar Collagens. Subcell Biochem. 2017, 82, 457–490. [Google Scholar]
- Mortimer, G.M.; Minchin, R.F. Cryptic epitopes and functional diversity in extracellular proteins. Int. J. Biochem. Cell Boil. 2016, 81, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.L.; Sukkar, M.B. The SPARC protein: An overview of its role in lung cancer and pulmonary fibrosis and its potential role in chronic airways disease. Br. J. Pharmacol. 2016, 174, 3–14. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Boil. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Rosset, E.M.; Bradshaw, A.D. SPARC/osteonectin in mineralized tissue. Matrix Boil. 2016, 52, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.W.H.; Harper, S.J.; McVey, J.H.; Hogan, B.L. In vivo expression of mRNA for the Ca++-binding protein SPARC (osteonectin) revealed by in situ hybridization. J. Cell Boil. 1987, 105, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Broken Energy Homeostasis and Obesity Pathogenesis: The Surrounding Concepts. J. Clin. Med. 2018, 7, 453. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.J.; Lagerpusch, M.; Enderle, J.; Schautz, B.; Heller, M.; Bosy-Westphal, A. Beyond the body mass index: Tracking body composition in the pathogenesis of obesity and the metabolic syndrome. Obes. Rev. 2012, 13, 6–13. [Google Scholar]
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef]
- Després, J.-P.; Fias, F. Body Fat Distribution and Risk of Cardiovascular Disease. Circulation 2012, 126, 1301–1313. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, A.D.; Graves, D.C.; Motamed, K.; Sage, E.H. SPARC-null mice exhibit increased adiposity without significant differences in overall body weight. Proc. Natl. Acad. Sci. USA 2003, 100, 6045–6050. [Google Scholar] [CrossRef] [Green Version]
- Riedl, I.; Yoshioka, M.; Nishida, Y.; Tobina, T.; Paradis, R.; Shono, N.; Tanaka, H.; St-Amand, J. Regulation of skeletal muscle transcriptome in elderly men after 6weeks of endurance training at lactate threshold intensity. Exp. Gerontol. 2010, 45, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Melouane, A.; Yoshioka, M.; St-Amand, J. Extracellular matrix/mitochondria pathway: A novel potential target for sarcopenia. Mitochondrion 2019, 50, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Romani, P.; Brian, I.; Santinon, G.; Pocaterra, A.; Audano, M.; Pedretti, S.; Mathieu, S.; Forcato, M.; Bicciato, S.; Manneville, J.-B.; et al. Extracellular matrix mechanical cues regulate lipid metabolism through Lipin-1 and SREBP. Nature 2019, 21, 338–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, W.J.; Mullen, P.J.; Schmid, E.W.; Flores, A.; Momcilovic, M.; Sharpley, M.S.; Jelinek, D.; Whiteley, A.E.; Maxwell, M.B.; Wilde, B.; et al. Extracellular Matrix Remodeling Regulates Glucose Metabolism through TXNIP Destabilization. Cell 2018, 175, 117–132.e21. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Guan, Y.; Zhang, L.; Li, K.; Dong, C. SPARC interacts with AMPK and regulates GLUT4 expression. Biochem. Biophys. Res. Commun. 2010, 396, 961–966. [Google Scholar] [CrossRef]
- Omi, S.; Yamanouchi, K.; Nakamura, K.; Matsuwaki, T.; Nishihara, M. Reduced fibrillar collagen accumulation in skeletal muscle of secreted protein acidic and rich in cysteine (SPARC)-null mice. J. Veter- Med Sci. 2019, 81, 1649–1654. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.H.; Bradshaw, A.D. The Influence of the Extracellular Matrix in Inflammation: Findings from the SPARC-Null Mouse; Anat Rec: Hoboken, NJ, USA, 2019. [Google Scholar]
- Gilmour, D.T.; Lyon, G.J.; Carlton, M.B.; Sanes, J.R.; Cunningham, J.M.; Anderson, J.R.; Hogan, B.L.; Evans, M.J.; Colledge, W.H. Mice deficient for the secreted glycoprotein SPARC/osteonectin/BM40 develop normally but show severe age-onset cataract formation and disruption of the lens. EMBO J. 1998, 17, 1860–1870. [Google Scholar] [CrossRef]
- Swaminathan, S.S.; Oh, D.-J.; Kang, M.H.; Ren, R.; Jin, R.; Gong, H.; Rhee, U.J. Secreted Protein Acidic and Rich in Cysteine (SPARC)-Null Mice Exhibit More Uniform Outflow. Investig. Opthalmol. Vis. Sci. 2013, 54, 2035–2047. [Google Scholar] [CrossRef] [PubMed]
- Sangaletti, S.; Tripodo, C.; Cappetti, B.; Casalini, P.; Chiodoni, C.; Piconese, S.; Santangelo, A.; Parenza, M.; Arioli, I.; Miotti, S.; et al. SPARC Oppositely Regulates Inflammation and Fibrosis in Bleomycin-Induced Lung Damage. Am. J. Pathol. 2011, 179, 3000–3010. [Google Scholar] [CrossRef] [PubMed]
- Rempel, S.A.; Hawley, R.C.; Gutiérrez, J.A.; Mouzon, E.; Bobbitt, K.R.; Lemke, N.; Schultz, C.R.; Schultz, L.R.; Golembieski, W.; Koblinski, J.; et al. Splenic and immune alterations of the Sparc-null mouse accompany a lack of immune response. Genes Immun. 2007, 8, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Pichler, R.H.; Bassuk, J.A.; Hugo, C.; Reed, M.J.; Eng, E.; Gordon, K.L.; Pippin, J.; Alpers, C.E.; Couser, W.G.; Sage, E.H.; et al. SPARC is expressed by mesangial cells in experimental mesangial proliferative nephritis and inhibits platelet-derived-growth-factor-medicated mesangial cell proliferation in vitro. Am. J. Pathol. 1996, 148, 1153–1167. [Google Scholar]
- Francki, A.; Motamed, K.; McClure, T.D.; Kaya, M.; Murri, C.; Blake, D.J.; Carbon, J.G.; Sage, E.H. SPARC regulates cell cycle progression in mesangial cells via its inhibition of IGF-dependent signaling. J. Cell. Biochem. 2003, 88, 802–811. [Google Scholar] [CrossRef]
- Ghanemi, A.; St-Amand, J. Interleukin-6 as a “metabolic hormone”. Cytokine 2018, 112, 132–136. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Y.; Ye, J. IL-6: A Potential Role in Cardiac Metabolic Homeostasis. Int. J. Mol. Sci. 2018, 19, 2474. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Liu, J.; Lau, C.W.; Huang, Y. IL-6 in diabetes and cardiovascular complications. Br. J. Pharmacol. 2014, 171, 3595–3603. [Google Scholar] [CrossRef] [Green Version]
- Erdogan, B.; Webb, D.J. Cancer-associated fibroblasts modulate growth factor signaling and extracellular matrix remodeling to regulate tumor metastasis. Biochem. Soc. Trans. 2017, 45, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Moriggi, M.; Giussani, M.; Torretta, E.; Capitanio, D.; Sandri, M.; Leone, R.; De Palma, S.; Vasso, M.; Vozzi, G.; Tagliabue, E.; et al. ECM Remodeling in Breast Cancer with Different Grade: Contribution of 2D-DIGE Proteomics. Proteom. 2018, 18, 1800278. [Google Scholar] [CrossRef] [PubMed]
- Yuzhalin, A.E.; Lim, S.Y.; Kutikhin, A.; Gordon-Weeks, A.N. Dynamic matrisome: ECM remodeling factors licensing cancer progression and metastasis. Biochim. Biophys. Acta (BBA) Bioenerg. 2018, 1870, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, P.; Huang, P. Cancer stem cells, metabolism, and therapeutic significance. Tumor Boil. 2016, 37, 5735–5742. [Google Scholar] [CrossRef] [PubMed]
- Lyssiotis, C.A.; Kimmelman, A.C. Metabolic Interactions in the Tumor Microenvironment. Trends Cell Boil. 2017, 27, 863–875. [Google Scholar] [CrossRef] [Green Version]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and cancer: A homeostatic hormone? Cytokine 2020, 127, 154996. [Google Scholar] [CrossRef]
- Irminger-Finger, I.; Kargul, J.; Laurent, G.J.; Laurent, G.J. Extra cellular matrix a modular soil for stem cells. Int. J. Biochem. Cell Boil. 2016, 81, 164. [Google Scholar] [CrossRef]
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Boil. 2014, 37, 1–14. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted Protein Acidic and Rich in Cysteine: Metabolic and Homeostatic Properties beyond the Extracellular Matrix Structure. Appl. Sci. 2020, 10, 2388. https://doi.org/10.3390/app10072388
Ghanemi A, Yoshioka M, St-Amand J. Secreted Protein Acidic and Rich in Cysteine: Metabolic and Homeostatic Properties beyond the Extracellular Matrix Structure. Applied Sciences. 2020; 10(7):2388. https://doi.org/10.3390/app10072388
Chicago/Turabian StyleGhanemi, Abdelaziz, Mayumi Yoshioka, and Jonny St-Amand. 2020. "Secreted Protein Acidic and Rich in Cysteine: Metabolic and Homeostatic Properties beyond the Extracellular Matrix Structure" Applied Sciences 10, no. 7: 2388. https://doi.org/10.3390/app10072388
APA StyleGhanemi, A., Yoshioka, M., & St-Amand, J. (2020). Secreted Protein Acidic and Rich in Cysteine: Metabolic and Homeostatic Properties beyond the Extracellular Matrix Structure. Applied Sciences, 10(7), 2388. https://doi.org/10.3390/app10072388