Functional Roles of Saccades for a Hand Movement


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Apparatus
2.2. Behavioral Tasks
2.3. Data Recording and Analysis
2.3.1. Saccade
2.3.2. Hand Movement
3. Results
3.1. Hitting Task
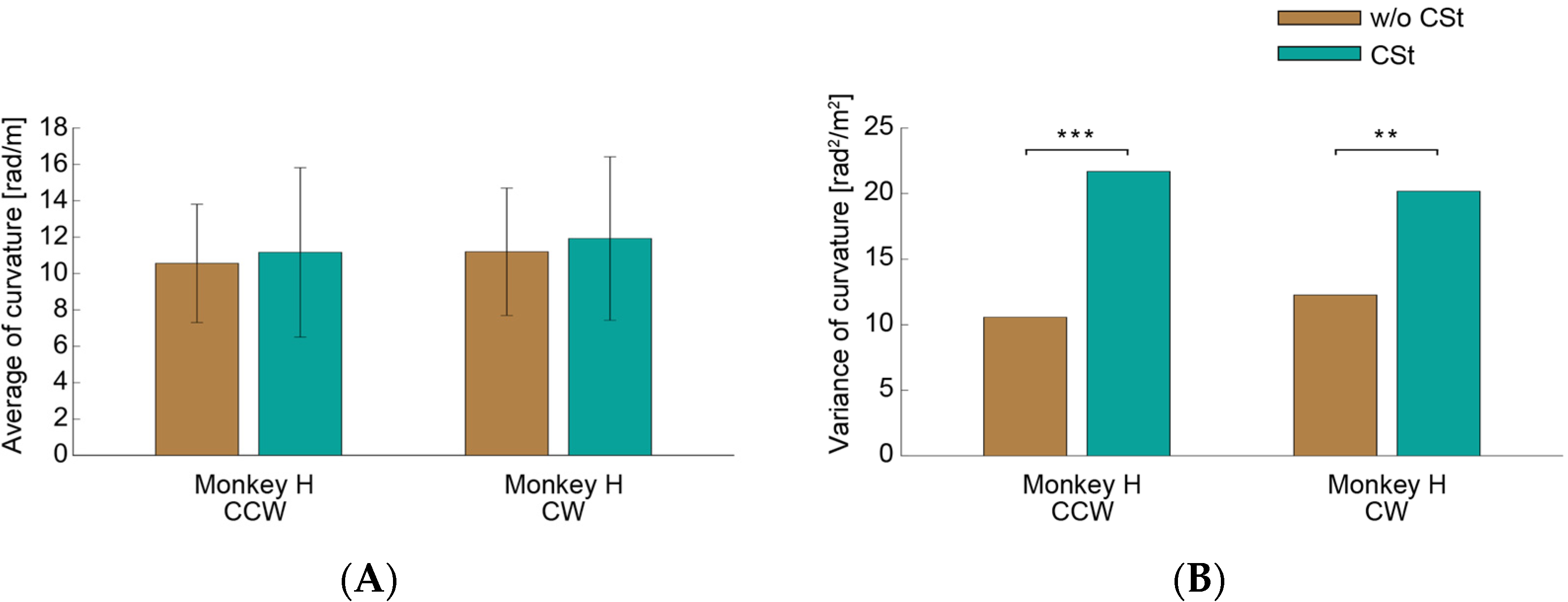
3.2. Circle-Drawing Task
4. Discussion
4.1. Functional Roles of the Saccades
4.2. The Saccades and the Control Model of an Arm Movement
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morasso, P.; Mussa Ivaldi, F.A. Trajectory formation and handwriting: A computational model. Biol. Cybern. 1982, 45, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Viviani, P.; Cenzato, M. Segmentation and coupling in complex movements. J. Exp. Psychol. Hum. Percept. Perform. 1985, 11, 828–845. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Kawato, M. A via-point time optimization algorithm for complex sequential trajectory formation. Neural Netw. 2004, 17, 353–364. [Google Scholar] [CrossRef]
- Morasso, P. Spatial Control of Arm Movements. Exp. Brain Res. 1981, 42, 223–227. [Google Scholar] [CrossRef]
- Gomi, H.; Kawato, M. Human arm stiffness and equilibrium-point trajectory during multi-joint movement. Biol. Cybern. 1997, 76, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Georgopoulos, A.P.; Kalaska, J.F.; Caminiti, R.; Massey, J.T. On the Relations between the Direction of Two-Dimensional Arm Movements and Cell Discharge in Primate Motor Cortex. J. Neurosci. 1982, 2, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.H.; Kalaska, J.F. Changes in motor cortex activity during reaching movements with similar hand paths but different arm postures. J. Neurophysiol. 1995, 73, 2563–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein Breteler, M.D.; Hondzinski, J.M.; Flanders, M. Drawing sequences of segments in 3D: Kinetic influences on arm configuration. J. Neurophysiol. 2003, 89, 3253–3263. [Google Scholar] [CrossRef]
- Abrams, R.A.; Meyer, D.E.; Kornblum, S. Eye-hand coordination: Oculomotor control in rapid aimed limb movements. J. Exp. Psychol. Hum. Percept. Perform. 1990, 16, 248–267. [Google Scholar] [CrossRef]
- Frens, M.A.; Erkelens, C.J. Coordination of hand movements and saccades: Evidence for a common and a separate pathway. Exp. Brain Res. 1991, 85, 682–690. [Google Scholar] [CrossRef]
- Bekkering, H.; Adam, J.J.; van den Aarssen, A.; Kingma, H.; Whiting, H.T. Interference between saccadic eye and goal-directed hand movements. Exp. Brain Res. 1995, 106, 475–484. [Google Scholar] [CrossRef]
- Helsen, W.F.; Elliott, D.; Starkes, J.L.; Ricker, K.L. Temporal and spatial coupling of point of gaze and hand movements in aiming. J. Motor Behav. 1998, 30, 249–259. [Google Scholar] [CrossRef]
- Lunenburger, L.; Kutz, D.F.; Hoffmann, K.P. Influence of arm movements on saccades in humans. Eur. J. Neurosci. 2000, 12, 4107–4116. [Google Scholar] [CrossRef]
- Diaz, G.; Cooper, J.; Rothkopf, C.; Hayhoe, M. Saccades to future ball location reveal memory-based prediction in a virtual-reality interception task. J. Vis. 2013, 13, 20. [Google Scholar] [CrossRef]
- Mann, D.L.; Nakamoto, H.; Logt, N.; Sikkink, L.; Brenner, E. Predictive eye movements when hitting a bouncing ball. J. Vis. 2019, 19, 28. [Google Scholar] [CrossRef] [Green Version]
- Beaubaton, D.; Hay, L. Contribution of Visual Information to Feedforward and Feedback Processes in Rapid Pointing Movements. Hum. Mov. Sci. 1986, 5, 19–34. [Google Scholar] [CrossRef]
- Desmurget, M.; Grafton, S. Forward modeling allows feedback control for fast reaching movements. Trends Cogn. Sci. 2000, 4, 423–431. [Google Scholar] [CrossRef]
- Saunders, J.A.; Knill, D.C. Humans use continuous visual feedback from the hand to control fast reaching movements. Exp. Brain Res. 2003, 152, 341–352. [Google Scholar] [CrossRef]
- Thaler, L.; Goodale, M.A. The role of online visual feedback for the control of target-directed and allocentric hand movements. J. Neurophysiol. 2011, 105, 846–859. [Google Scholar] [CrossRef] [Green Version]
- Tchalenko, J. Eye movements in drawing simple lines. Perception 2007, 36, 1152–1167. [Google Scholar] [CrossRef]
- Ueyama, Y.; Miyashita, E. Devising a Robotic Arm Manipulandum for Normal and Altered Reaching Movements to Investigate Brain Mechanisms of Motor Control. Instrum. Sci. Technol. 2013, 41, 251–273. [Google Scholar] [CrossRef]
- Hinder, M.R.; Tresilian, J.R.; Riek, S.; Carson, R.G. The contribution of visual feedback to visuomotor adaptation: How much and when? Brain Res. 2008, 1197, 123–134. [Google Scholar] [CrossRef]
- Milner, T.E. A model for the generation of movements requiring endpoint precision. Neuroscience 1992, 49, 487–496. [Google Scholar] [CrossRef]
- Plamondon, R.; Alimi, A.M. Speed/accuracy trade-offs in target-directed movements. Behav. Brain Sci. 1997, 20, 279. [Google Scholar] [CrossRef]
- Matthis, J.S.; Yates, J.L.; Hayhoe, M.M. Gaze and the Control of Foot Placement When Walking in Natural Terrain. Curr. Biol. 2018, 28, 1224–1233. [Google Scholar] [CrossRef] [Green Version]
- Todorov, E.; Jordan, M.I. Optimal feedback control as a theory of motor coordination. Nat. Neurosci. 2002, 5, 1226–1235. [Google Scholar] [CrossRef]
- Liu, D.; Todorov, E. Evidence for the flexible sensorimotor strategies predicted by optimal feedback control. J. Neurosci. 2007, 27, 9354–9368. [Google Scholar] [CrossRef]
- Izawa, J.; Rane, T.; Donchin, O.; Shadmehr, R. Motor adaptation as a process of reoptimization. J. Neurosci. 2008, 28, 2883–2891. [Google Scholar] [CrossRef]
- Nagengast, A.J.; Braun, D.A.; Wolpert, D.M. Optimal control predicts human performance on objects with internal degrees of freedom. PLoS Comput. Biol. 2009, 5, e1000419. [Google Scholar] [CrossRef]
- Ueyama, Y.; Miyashita, E. Optimal Feedback Control for Predicting Dynamic Stiffness During Arm Movement. IEEE Trans. Ind. Electron. 2014, 61, 1044–1052. [Google Scholar] [CrossRef]
- Shadmehr, R.; Mussa-Ivaldi, F.A. Adaptive representation of dynamics during learning of a motor task. J. Neurosci. 1994, 14, 3208–3224. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, J.W.; Pine, Z.M.; Ghilardi, M.F.; Ghez, C. Learning of visuomotor transformations for vectorial planning of reaching trajectories. J. Neurosci. 2000, 20, 8916–8924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osu, R.; Burdet, E.; Franklin, D.W.; Milner, T.E.; Kawato, M. Different mechanisms involved in adaptation to stable and unstable dynamics. J. Neurophysiol. 2003, 90, 3255–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, N.; Gomi, H. Multiple motor learning strategies in visuomotor rotation. PLoS ONE 2010, 5, e9399. [Google Scholar] [CrossRef]
- Wang, J.X.; Kurth-Nelson, Z.; Kumaran, D.; Tirumala, D.; Soyer, H.; Leibo, J.Z.; Hassabis, D.; Botvinick, M. Prefrontal cortex as a meta-reinforcement learning system. Nat. Neurosci. 2018, 21, 860–868. [Google Scholar] [CrossRef]
- Sugiyama, T.; Schweighofer, N.; Izawa, J. Reinforcement meta-learning optimizes visuomotor learning. bioRxiv 2020. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Number of Trials | Saccade [Times/Trial] (Number of Trials) | ||||
|---|---|---|---|---|---|---|
| Total | LS | Total CS | CSa 1 | CSd 2 | ||
| Monkey H | 762 | 2.51 (762) | 1.77 (762) | 0.35 (242) | 0.27 (199) 3 | 0.09 (67) 3 |
| Monkey U | 556 | 1.49 (556) | 1.16 (540) | 0.01 (8) | 0.01(8) | 0.00 (0) |
| Subject | Direction 1 | Trial | Saccade [Times/Trial] (Number of Trials) All/Midway 2 | ||
|---|---|---|---|---|---|
| Total 3 | LS | CS | |||
| Monkey H | CCW | 743 | 5.22 (741)/ 1.81 (723) | 2.33 (735)/ 0.83 (457) | 2.27 (694)/ 0.86 (447) |
| CW | 751 | 4.77 (751)/ 1.72 (648) | 1.78 (714)/ 0.75 (439) | 2.22 (668)/ 0.88 (441) | |
| Monkey U | CCW | 749 | 4.70 (749)/ 3.04 (749) | 1.70 (691)/ 0.89 (531) | 1.80 (638)/ 1.35 (624) |
| CW | 769 | 4.60 (769)/ 2.29 (769) | 1.10 (679)/ 0.16 (110) | 2.60 (744)/ 1.73 (729) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakazume, Y.; Furubayashi, S.; Miyashita, E. Functional Roles of Saccades for a Hand Movement. Appl. Sci. 2020, 10, 3066. https://doi.org/10.3390/app10093066
Sakazume Y, Furubayashi S, Miyashita E. Functional Roles of Saccades for a Hand Movement. Applied Sciences. 2020; 10(9):3066. https://doi.org/10.3390/app10093066
Chicago/Turabian StyleSakazume, Yuki, Sho Furubayashi, and Eizo Miyashita. 2020. "Functional Roles of Saccades for a Hand Movement" Applied Sciences 10, no. 9: 3066. https://doi.org/10.3390/app10093066
APA StyleSakazume, Y., Furubayashi, S., & Miyashita, E. (2020). Functional Roles of Saccades for a Hand Movement. Applied Sciences, 10(9), 3066. https://doi.org/10.3390/app10093066