
Arthrospira platensis as a Feasible Feedstock for Bioethanol Production


Abstract
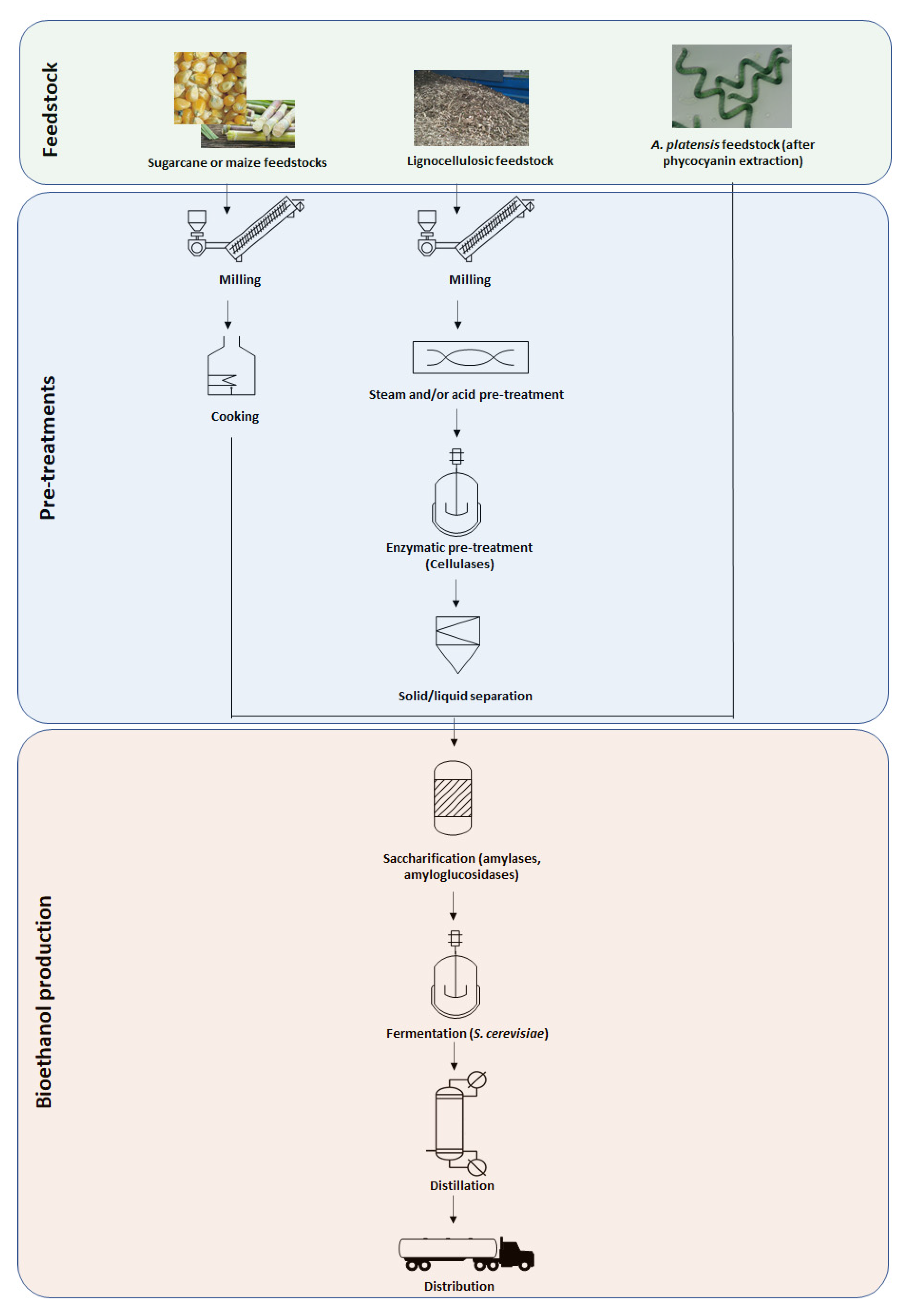
:1. Introduction
2. Growth Requirements for Biomass Production
3. Environmental Stresses for Glycogen Accumulation
3.1. Light Intensity

{kind=link}
| Enhancement Technique | Carbohydrates | Biomass | Culture Conditions | Observations | Reference |
|---|---|---|---|---|---|
| Light intensity | Glycogen (45% DW) | 1.6 g L−1 | Light intensity: 700 µmol/m2/s, temperature: 30 °C, volume: 250 mL, time: 5 days, 100 rpm | Effect of different light intensities on the production of glycogen of A. platensis 50, 270, 700 µmol/m2/s | [62] |
| Light intensity | EPS/glycogen ratios (3.1, 1.79, 1.54, 0.64) | 6.8 10−3, 1.8 10−2, 2.9 10−2, 3.1 10−2 g L−1 h−1 | Light intensity: 100, 400, 800 and 1200 µmol/m2/s, temperature: 35 °C, Time: 12 days, 1.3 L, Aeration: CO2 1%, Flow: 11 mL min−1 | Effect of different light intensities on the glycogen and EPS production of A. platensis, analysis of EPS composition. 100, 400, 800 and 1200 µmol/m2/s | [63] |
| Nitrogen deficiency | Glycogen (30% DW) | 0.6 g/L | Light intensity: 100 µmol/m2/s, temperature: 35 °C, volume: 500 mL, time: 96 h | The control amounts of glycogen and biomass were: 15% DW and 1.3 g/L respectively. Also, flocculation capability under nutrient starvation | [64] |
| Nitrogen deficiency | Glycogen (1.03 g L−1) | 1.6 g L−1 | Light intensity: 700 µmol/m2/s, temperature: 30 °C, volume: 250 mL, time: 5 days, 100 rpm | Nitrate: 3 mM, Light intensity (50, 270, 700 µmol/m2/s) | [61] |
| Nitrogen deficiency | Carbohydrates (73.2% DW) | 0.7 g L−1 | Light intensity: 40 µmol/m2/s, light:dark cycle 14 h:10 h, temperature: 28 °C, volume: 200 mL, time: 5 days | NaNO3: 15 mg/L | [31] |
| Nitrogen deficiency | Carbohydrates (74% DW) | 0.66 g L−1 | Light intensity: 100 µmol/m2/s, temperature: 20 °C, volume: 2000 mL, time: 9 days, light/dark cycle 16/8, bubbling: 0.2 µm-filtered air | NaNO3: 20 mg/L, biomass concentrated 15 times by gravity settling | [29] |
| Nitrogen deficiency | Carbohydrates (64.3%) | 27.5 g/m2/d | Natural daylight, temperature: 25–35 °C, volume: 180 m3, time: 90 h | Nitrate: 4.7 mg/L | [65] |
| Nitrogen deficiency | Glycogen (63.2%) | 1.1 g L−1 | Light intensity: 50 µmol/m2/s, temperature: 30 °C, volume: 250 mL, time: 7 days, agitation: 100 rpm | NaNO3: 29.4 mM, there was no difference between the medium with and without nitrogen depletion | [66] |
| Nitrogen deficiency | Total sugars (10 µg/mg DW) | -- | Light intensity: 43 µmol/m2/s, temperature: 30 °C, Time: 168 h | Nitrate free Zarrouk medium | [67] |
| Phosphorus deficiency | Glycogen (70% DW) | 0.6 g L−1 | Light intensity: 100 µmol/m2/s, temperature: 35 °C, volume: 500 mL, time: 96 h | The control amounts of glycogen and biomass were: 0.15 g/g DW and 1.3 g/L DW respectively. Also, flocculation capability under nutrient starvation | [64] |
| Phosphorus deficiency | Carbohydrates (63.1% DW) | 1.8 g L−1 | Light intensity: 120 µmol/m2/s, temperature: 30 °C, volume: 500 mL, time: 7 days, Aeration: Filtered air flow: 0.2 v/v min | K2HPO4: 20 mg/L | [32] |
| Phosphorus deficiency | Carbohydrates (65% DW) | -- | Light intensity: 20 µmol/m2/s, temperature: 30 °C, time: 27 days, volume: 250 mL, Aeration: Filtered air flow: 0.4 v/v min | K2HPO4: 2 mg/g of biomass (intracellular phosphorus), C/N ratio increased from 4.6 to 12.2 | [32] |
| Sodium stress | Carbohydrates (53.4% DW) | 2.1 g L−1 | Light intensity: 8000 lux, temperature: 27 °C, volume: 100 mL, time: 7 days, pH: 9.8, Aeration: 15% CO2 v/v flow: 30 mL/min | NaCl: 0.5 mol/L related to seawater concentration | [26] |
| Sodium stress | Carbohydrates (30 mg/L/d) | 128 mg/L/d | Light intensity: 2500 lux, temperature: 30 °C, volume: 750 mL, time: 20 days | Different parameters where evaluated: NaCl, nitrate, phosphorous concentrations | [68] |
| Sodium stress | Carbohydrates (31%DW) | 128 mg/L/d | Light intensity: 80 µmol/m2/s, temperature: 30 °C, Time: 10 days, volume: 8000 mL, Aeration: Filtered air, Flow: 0.1 v/v min, Air bubble size: 3 mm | NaCl: 40 g/L, NaNO3: 0.5 g/L, K2HPO4: 0.5 g/L | [61] |
| Sulphur deficiency | Glycogen (68% DW) | 1.3 g L−1 | Light intensity: 100 µmol/m2/s, temperature: 35 °C, volume: 500 mL, time: 96 h | The control amounts of glycogen and biomass were: 0.15 g/g DW and 1.3 g/L DW, respectively. Also, flocculation capability under nutrient starvation | [64] |
| Sulphur deficiency | Carbohydrates (61 mg/L/d) | 134 mg/L/d | Light intensity: 2500 lux, temperature: 30 °C, volume: 750 mL, time: 20 days | Different parameters where evaluated: sulfur, nitrate, phosphorous concentrations | [68] |
3.2. Nitrogen Depletion
3.3. Phosphorous Depletion
3.4. Sulfur Depletion
3.5. Salinity Effect
3.6. Combined Stresses
4. Techno-Economical Considerations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chiong, M.C.; Chong, C.T.; Ng, J.-H.; Lam, S.S.; Tran, M.-V.; Chong, W.W.F.; Jaafar, M.N.M.; Valera-Medina, A. Liquid biofuels production and emissions performance in gas turbines: A review. Energy Convers. Manag. 2018, 173, 640–658. [Google Scholar] [CrossRef] [Green Version]
- Bala, J.D.; Lalung, J.; Al-Gheethi, A.A.S.; Norli, I. A review on biofuel and bioresources for environmental applications. In Renewable Energy and Sustainable Technologies for Building and Environmental Applications: Options for a Greener Future; Ahmad, M.I., Ismail, M., Riffat, S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 205–225. [Google Scholar]
- Sudhakar, K.; Mamat, R.; Samykano, M.; Azmi, W.; Ishak, W.; Yusaf, T. An overview of marine macroalgae as bioresource. Renew. Sustain. Energy Rev. 2018, 91, 165–179. [Google Scholar] [CrossRef]
- Jambo, S.A.; Abdulla, R.; Azhar, S.H.M.; Marbawi, H.; Gansau, J.A.; Ravindra, P. A review on third generation bioethanol feedstock. Renew. Sustain. Energy Rev. 2016, 65, 756–769. [Google Scholar] [CrossRef]
- Lima, G.M.; Teixeira, P.C.N.; Teixeira, C.M.L.L.; Filócomo, D.; Lage, C.L.S. Influence of spectral light quality on the pigment concentrations and biomass productivity of Arthrospira platensis. Algal Res. 2018, 31, 157–166. [Google Scholar] [CrossRef]
- Scaife, M.A.; Merkx-Jacques, A.; Woodhall, D.L.; Armenta, R.E. Algal biofuels in Canada: Status and potential. Renew. Sustain. Energy Rev. 2015, 44, 620–642. [Google Scholar] [CrossRef]
- Merlo, S.; Gabarrell Durany, X.; Pedroso Tonon, A.; Rossi, S. Marine microalgae contribution to sustainable development. Water 2021, 13, 1373. [Google Scholar] [CrossRef]
- Vonshak, A. Physiology, Cell-Biology and Biotechnology; Taylor & Francis: London, UK, 1997. [Google Scholar]
- Masojídek, J.; Torzillo, G. Mass cultivation of freshwater microalgae. In Earth Systems and Environmental Sciences, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Cheng, J.; Zhu, Y.; Xu, X.; Zhang, Z.; Yang, W. Enhanced biomass productivity of Arthrospira platensis using zeolitic imidazolate framework-8 as carbon dioxide adsorbents. Bioresour. Technol. 2019, 294, 122118. [Google Scholar] [CrossRef] [PubMed]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A potential alternative to health supplementation for humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—A new bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Ma, Z.; Ahmed, F.; Yuan, B.; Zhang, W. Fresh living Arthrospira as dietary supplements: Current status and challenges. Trends Food Sci. Technol. 2019, 88, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Cheng, J.; Zhang, Z.; Liu, J. Mutation of Arthrospira platensis by gamma irradiation to promote phenol tolerance and CO2 fixation for coal-chemical flue gas reduction. J. CO2 Util. 2020, 38, 252–261. [Google Scholar] [CrossRef]
- Poughon, L.; Laroche, C.; Creuly, C.; Dussap, C.-G.; Paille, C.; Lasseur, C.; Monsieurs, P.; Heylen, W.; Coninx, I.; Mastroleo, F.; et al. Limnospira indica PCC8005 growth in photobioreactor: Model and simulation of the ISS and ground experiments. Life Sci. Space Res. 2020, 25, 53–65. [Google Scholar] [CrossRef]
- Nematollahi, M.A.; Laird, D.W.; Hughes, L.J.; Raeisossadati, M.; Moheimani, N.R. Effect of organic carbon source and nutrient depletion on the simultaneous production of a high value bioplastic and a specialty pigment by Arthrospira platensis. Algal Res. 2020, 47, 101844. [Google Scholar] [CrossRef]
- Rubio, C.; Dominik-Jakubiec, M.; Paz, S.; Gutiérrez, Á.J.; González-Weller, D.; Hardisson, A. Dietary exposure to trace elements (B, Ba, Li, Ni, Sr, and V) and toxic metals (Al, Cd, and Pb) from the consumption of commercial preparations of Spirulina platensis. Environ. Sci. Pollut. Res. Int. 2021, 28, 22146–22155. [Google Scholar] [CrossRef]
- Kałduńska, J.; Jakubczak, K.; Gutowska, I.; Dalewski, B.; Janda, K. Fluoride content in dietary supplements of spirulina (Arthrospira spp.) from conventional and organic cultivation. Fluoride 2020, 5, 469–476. [Google Scholar]
- Cheng, J.; Xia, A.; Song, W.; Su, H.; Zhou, J.; Cen, K. Comparison between heterofermentation and autofermentation in hydrogen production from Arthrospira (Spirulina) platensis wet biomass. Int. J. Hydrogen Energy 2012, 37, 6536–6544. [Google Scholar] [CrossRef]
- Lee, M.; Chen, Y.; Peng, T. Two-stage culture method for optimized polysaccharide production in Spirulina platensis. J. Sci. Food Agric. 2012, 92, 1562–1569. [Google Scholar] [CrossRef]
- Mendoza, Á.; Morales, V.; Sánchez-Bayo, A.; Rodríguez-Escudero, R.; González-Fernández, C.; Bautista, L.F.; Vicente, G. The effect of the lipid extraction method used in biodiesel production on the integrated recovery of biodiesel and biogas from Nannochloropsis gaditana, Isochrysis galbana and Arthrospira platensis. Biochem. Eng. J. 2020, 154, 107428. [Google Scholar] [CrossRef]
- Sydney, E.B.; Sturm, W.; de Carvalho, J.C.; Thomaz-Soccol, V.; Larroche, C.; Pandey, A.; Soccol, C.R. Potential carbon dioxide fixation by industrially important microalgae. Bioresour. Technol. 2010, 101, 5892–5896. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Patel, A.K.; Hong, M.E.; Chang, W.S.; Sim, S.J. Microalgae Bioenergy with Carbon Capture and Storage (BECCS): An emerging sustainable bioprocess for reduced CO2 emission and biofuel production. Bioresour. Technol. Rep. 2019, 7, 100270. [Google Scholar] [CrossRef]
- Ho, S.-H.; Zhang, C.; Tao, F.; Zhang, C.; Chen, W.-H. Microalgal torrefaction for solid biofuel production. Trends Biotechnol. 2020, 38, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Pourkarimi, S.; Hallajisani, A.; Alizadehdakhel, A.; Nouralishahi, A. Biofuel production through micro- and macroalgae pyrolysis—A review of pyrolysis methods and process parameters. J. Anal. Appl. Pyrolysis 2019, 142, 104599. [Google Scholar] [CrossRef]
- Ding, L.; Cheng, J.; Lu, H.; Yue, L.; Zhou, J.; Cen, K. Three-stage gaseous biofuel production combining dark hydrogen, photo hydrogen, and methane fermentation using wet Arthrospira platensis cultivated under high CO2 and sodium stress. Energy Convers. Manag. 2017, 148, 394–404. [Google Scholar] [CrossRef]
- Serrà, A.; Artal, R.; García-Amorós, J.; Gómez, E.; Philippe, L. Circular zero-residue process using microalgae for efficient water decontamination, biofuel production, and carbon dioxide fixation. Chem. Eng. J. 2020, 388, 124278. [Google Scholar] [CrossRef]
- Monshupanee, T.; Incharoensakdi, A. Enhanced accumulation of glycogen, lipids and polyhydroxybutyrate under optimal nutrients and light intensities in the cyanobacterium Synechocystis sp. PCC 6803. J. Appl. Microbiol. 2014, 116, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Depraetere, O.; Deschoenmaeker, F.; Badri, H.; Foubert, I.; Leys, N.; Wattiez, R.; Muylaert, K. Trade-Off between Growth and Carbohydrate Accumulation in Nutrient-Limited Arthrospira sp. PCC 8005 Studied by Integrating Transcriptomic and Proteomic Approaches. PLoS ONE 2015, 10, e0132461. [Google Scholar]
- Arias, D.M.; García, J.; Uggetti, E. Production of polymers by cyanobacteria grown in wastewater: Current status, challenges and future perspectives. New Biotechnol. 2020, 55, 46–57. [Google Scholar] [CrossRef]
- Yao, C.; Pan, Y.; Lu, H.; Wu, P.; Meng, Y.; Cao, X.; Xue, S. Utilization of recovered nitrogen from hydrothermal carbonization process by Arthrospira platensis. Bioresour. Technol. 2016, 212, 26–34. [Google Scholar] [CrossRef]
- Markou, G. Alteration of the biomass composition of Arthrospira (Spirulina) platensis under various amounts of limited phosphorus. Bioresour. Technol. 2012, 116, 533–535. [Google Scholar] [CrossRef]
- Nowicka-Krawczyk, P.; Mühlsteinová, R.; Hauer, T. Detailed characterization of the Arthrospira type species separating commercially grown taxa into the new genus Limnospira (Cyanobacteria). Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution à l’étude d’une Cyanophycée: Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthèse de Spirulina Maxima. Ph.D. Thesis, Université de Paris, Paris, France, 1966. [Google Scholar]
- Richmond, A.E. Microalgaculture. In CRC Critical Reviews in Biotechnology; CRC Press Inc.: Boca Raton, FL, USA, 1988; Volume 4. [Google Scholar]
- Chojnacka, K.; Noworita, A. Evaluation of Spirulina sp growth in phototrophic, heterotrophic and mixotrophic cultures. Enzyme Microb. Technol. 2004, 34, 461–465. [Google Scholar] [CrossRef]
- Cogne, G.; Lehmann, B.; Dussap, C.-G.; Gros, J.-B. Uptake of macrominerals and trace elements by the cyanobacterium Spirulina platensis (Arthrospira platensis PCC 8005) under photoautotrophic conditions: Culture medium optimization. Biotechnol. Bioeng. 2003, 81, 588–593. [Google Scholar] [CrossRef]
- Belkin, S.; Boussiba, S. Resistance of Spirulina platensis to ammonia at high pH values. Plant Cell Physiol. 1991, 32, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.R.; Colman, B. Inorganic carbon accumulation and photosynthesis in a blue-green alga as function of external pH. Plant Physiol. 1981, 67, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, W.D.; Mwatha, W.E.; Jones, B.E. Alkaliphiles: Ecology, diversity and applications. FEMS Microbiol. Rev. 1990, 75, 255–270. [Google Scholar] [CrossRef]
- Iltis, A. Tolérance de salinité de Spirulina platensis (Gom.) Geitler, (Cyanophyta) dans les mares natronées du Kanem (Tchad). Cah ORSTOM Sér Hydrobiol. 1968, 2, 119–125. [Google Scholar]
- Cornet, J.-F. Etude Cinétique et Energétique d’un Photobioréacteur. Etablissement d’un Modèle Structure. Applications à un Ecosystème clos Artificiel. Ph.D. Thesis, Université de Paris, Paris, France, 1992. [Google Scholar]
- Torzillo, G.; Vonshak, A. Effect of light and temperature on the photosynthetic activity of the cyanobacterium Spirulina platensis. Biomass Bioenergy 1994, 6, 399–405. [Google Scholar] [CrossRef]
- Bocci, F.; Ferrari, F.; Materassi, R.; Mannelli, D. Prime Ricerche Sulla Crescita di Spirulina platensis e Spirulina Maxima in Coltura Fotolimitata; Atti XIX Congr Naz Soc Ital Microbiol: Catania, Italy, 1980. [Google Scholar]
- Chen, C.-Y.; Kao, P.-C.; Tsai, C.-J.; Lee, D.-J.; Chang, J.-S. Engineering strategies for simultaneous enhancement of C-phycocyanin production and CO2 fixation with Spirulina platensis. Bioresour. Technol. 2013, 145, 307–312. [Google Scholar] [CrossRef]
- Soletto, D.; Binaghi, L.; Ferrari, L.; Lodi, A.; Carvalho, J.C.M.; Zilli, M.; Converti, A. Effects of carbon dioxide feeding rate and light intensity on the fed-batch pulse-feeding cultivation of Spirulina platensis in helical photobioreactor. Biochem. Eng. J. 2008, 39, 369–375. [Google Scholar] [CrossRef]
- Kaplan, A. Photoinhibition in Spirulina platensis: Response of photosynthesis and HCO3− uptake capability to CO2—depleted conditions. J. Exp. Bot. 1981, 32, 669. [Google Scholar] [CrossRef]
- Flores, E.; Frías, J.E.; Rubio, L.M.; Herrero, A. Photosynthetic nitrate assimilation in cyanobacteria. Photosyn. Res. 2005, 83, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Boussiba, S. Ammonia uptake in the alkalophilic cyanobacterium Spirulina platensis. Plant Cell Physiol. 1989, 30, 303–308. [Google Scholar] [CrossRef]
- Boussiba, S.; Gibson, J. Ammonia translocation in cyanobacteria. FEMS Microbiol. Rev. 1991, 88, 1–14. [Google Scholar]
- Markou, G.; Vandamme, D.; Muylaert, K. Ammonia inhibition on Arthrospira platensis in relation to the initial biomass density and pH. Bioresour. Technol. 2014, 166, 259–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeliovitch, A.; Azov, Y. Toxicity of ammonia to algae in sewage oxidation pounds. Appl. Environ. Microbiol. 1976, 31, 801–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, G.-Z.; Qiu, B.-S.; Forchhammer, K. Ammonium tolerance in the cyanobacterium Synechocystis sp. strain PCC 6803 and the role of the psbA multigene family. Plant Cell Environ. 2014, 37, 840–851. [Google Scholar] [CrossRef]
- Drath, M.; Kloft, N.; Batschauer, A.; Marin, K.; Novak, J.; Forchhammer, K. Ammonia triggers photodamage of photosystem II in the cyanobacterium Synechocystis sp. strain PCC 6803. Plant Physiol. 2008, 147, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Avila-Leon, I.; Chuei-Matsudo, M.; Sato, S.; de Carvalho, J.C.M. Arthrospira platensis biomass with high protein content cultivated in continuous process using urea as nitrogen source. J Appl. Microbiol. 2012, 112, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Phélippé, M.; Gonçalves, O.; Thouand, G.; Cogne, G.; Laroche, C. Characterization of the polysaccharides chemical diversity of the cyanobacteria Arthrospira platensis. Algal Res. 2019, 38, 101426. [Google Scholar] [CrossRef]
- Chentir, I.; Doumandji, A.; Ammar, J.; Zili, F.; Jridi, M.; Markou, G.; Ben Ouada, H. Induced change in Arthrospira sp. (Spirulina) intracellular and extracellular metabolites using multifactor stress combination approach. J. Appl. Phycol. 2018, 30, 1563–1574. [Google Scholar] [CrossRef]
- Klähn, S.; Hagemann, M. Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 2011, 13, 551–562. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Aikawa, S.; Kojima, Y.; Toya, Y.; Furusawa, C.; Kondo, A.; Shimizu, H. Construction of a Genome-Scale Metabolic Model of Arthrospira platensis NIES-39 and Metabolic Design for Cyanobacterial Bioproduction. PLoS ONE 2015, 10, e0144430. [Google Scholar]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef]
- Aikawa, S.; Izumi, Y.; Matsuda, F.; Hasunuma, T.; Chang, J.-S.; Kondo, A. Synergistic enhancement of glycogen production in Arthrospira platensis by optimization of light intensity and nitrate supply. Bioresour. Technol. 2012, 108, 211–215. [Google Scholar] [CrossRef]
- Klanchui, A.; Dulsawat, S.; Chaloemnegam, K.; Cheevadhanarak, S.; Prommeenate, P.; Meechai, A. An improved Genome-scale metabolic model of Arthrospira platensis C1 (iAK888) and its application in glycogen overproduction. Metabolites 2018, 8, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Yao, C.; Sun, Y.; Chen, W.; Tan, H.; Cao, X.; Xue, S.; Yin, H. Production and structural characterization of a new type of polysaccharide from nitrogen-limited Arthrospira platensis cultivated in outdoor industrial-scale open raceway ponds. Biotechnol. Biofuels 2019, 12, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Hasunuma, T.; Kikuyama, F.; Matsuda, M.; Aikawa, S.; Izumi, Y.; Kondo, A. Dynamic metabolic profiling of cyanobacterial glycogen biosynthesis under conditions of nitrate depletion. J. Exp. Bot. 2013, 64, 2943–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschoenmaeker, F.; Bayon-Vicente, G.; Sachdeva, N.; Depraetere, O.; Cabrera Pino, J.C.; Leroy, B.; Muylaert, K.; Wattiez, R. Impact of different nitrogen sources on the growth of Arthrospira sp. PCC 8005 under batch and continuous cultivation—A biochemical, transcriptomic and proteomic profile. Bioresour. Technol. 2017, 237, 78–88. [Google Scholar] [CrossRef]
- El-Shouny, W.; Sharaf, M.; Abomohra, A.E.-F.; Abo-Eleneen, M. Production enhancement of some valuable compounds of Arthrospira platensis. J. Bas. E Environ. Sci. 2015, 2, 74–85. [Google Scholar]
- Kashith, M.; Keerthana, B.; Sriram, S.; Ramamurthy, V. Adenylate cyclase in Arthrospira platensis responds to light through transcription. Biochem. Biophy. Res. Com. 2016, 477, 297–301. [Google Scholar] [CrossRef]
- Karniol, B.; Wagner, J.R.; Walker, J.M.; Vierstra, R.D. Phylogenetic analysis of the phytochrome superfamily reveals distinct microbial subfamilies of photoreceptors. Biochem. J. 2005, 392, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Strange, R.E. Bacterial glycogen and survival. Nature 1968, 220, 606–607. [Google Scholar] [CrossRef] [PubMed]
- Grossman, A.R.; Schaefer, M.R.; Chiang, G.G.; Collier, J.L. Environmental effects on the light-harvesting complex of cyanobacteria. J. Bacteriol. 1993, 175, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Ballicora, M.A.; Iglesias, A.A.; Preiss, J. ADP-Glucose pyrophosphorylase, a regulatory enzyme for bacterial glycogen synthesis. MMBR 2003, 67, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, D.M.; Fradinho, J.C.; Uggetti, E.; García, J.; Oehmen, A.; Reis, M.A. Polymer accumulation in mixed cyanobacterial cultures selected under the feast and famine strategy. Algal Res. 2018, 33, 99–108. [Google Scholar] [CrossRef]
- Cheng, D.; He, Q. Assessment of environmental stresses for enhanced microalgal biofuel production–an overview. Front. Energy Res. 2014, 2, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Kumaresan, V.; Nizam, F.; Ravichandran, G.; Viswanathan, K.; Palanisamy, R.; Bhatt, P.; Arasu, M.V.; Al-Dhabi, N.A.; Mala, K.; Arockiaraj, J. Transcriptome changes of blue-green algae, Arthrospira sp. in response to sulfate stress. Algal Res. 2017, 23, 96–103. [Google Scholar] [CrossRef]
- Hincha, D.K.; Hagemann, M. Stabilization of model membranes during drying by compatible solutes involved in the stress tolerance of plants and microorganisms. Biochem. J. 2004, 383, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Carrieri, D.; Ananyev, G.; Lenz, O.; Bryant, D.A.; Dismukes, G.C. Contribution of a sodium ion gradient to energy conservation during fermentation in the cyanobacterium Arthrospira (Spirulina) maxima CS-328. Appl. Environ. Microbiol. 2011, 77, 7185–7194. [Google Scholar] [CrossRef] [Green Version]
- Trewavas, A.J.; Rodrigues, C.; Rato, C.; Malhó, R. Cyclic nucleotides: The current dilemma. Curr. Opin. Plant Biol. 2002, 5, 425–429. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.S.; Costa, B.; Woiciechowski, A.L.; de Carvalho, J.C.; Medeiros, A.B.P.; Francisco, A.M.; Bonomi, L.J. Brazilian biofuel program: An overview. J. Sci. Ind. Res. 2005, 64, 897–904. [Google Scholar]
- Gulati, M.; Kohlmann, K.; Ladisch, M.R.; Hespell, R.; Bothast, R.J. Assessment of ethanol production options for corn products. Bioresour. Technol. 1996, 58, 253–264. [Google Scholar] [CrossRef]
- Mohanty, S.K.; Swain, M.R. Bioethanol production from corn and wheat: Food, fuel, and future. Bioethanol Prod. Food Crops 2019, 45–59. [Google Scholar] [CrossRef]
- Bothast, R.J.; Schlicher, M.A. Biotechnological processes for conversion of corn into ethanol. Appl. Microbiol. Biotechnol. 2005, 67, 19–25. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Freitas, B.C.B.; Rosa, G.M.; Moraes, L.; Morais, M.G.; Mitchell, B.G. Operational and economic aspects of Spirulina-based biorefinery. Bioresour. Technol. 2019, 292, 121946. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez Bautista, E.; Laroche, C. Arthrospira platensis as a Feasible Feedstock for Bioethanol Production. Appl. Sci. 2021, 11, 6756. https://doi.org/10.3390/app11156756
Gonzalez Bautista E, Laroche C. Arthrospira platensis as a Feasible Feedstock for Bioethanol Production. Applied Sciences. 2021; 11(15):6756. https://doi.org/10.3390/app11156756
Chicago/Turabian StyleGonzalez Bautista, Enrique, and Céline Laroche. 2021. "Arthrospira platensis as a Feasible Feedstock for Bioethanol Production" Applied Sciences 11, no. 15: 6756. https://doi.org/10.3390/app11156756
APA StyleGonzalez Bautista, E., & Laroche, C. (2021). Arthrospira platensis as a Feasible Feedstock for Bioethanol Production. Applied Sciences, 11(15), 6756. https://doi.org/10.3390/app11156756