
Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Microbiological Analysis
2.3. Identification of Isolates
2.4. Evaluation of Virulence Factors: Hemolysin and Gelatinase Production
2.5. Determination of the Minimum Inhibitory Concentration (MIC)
3. Results
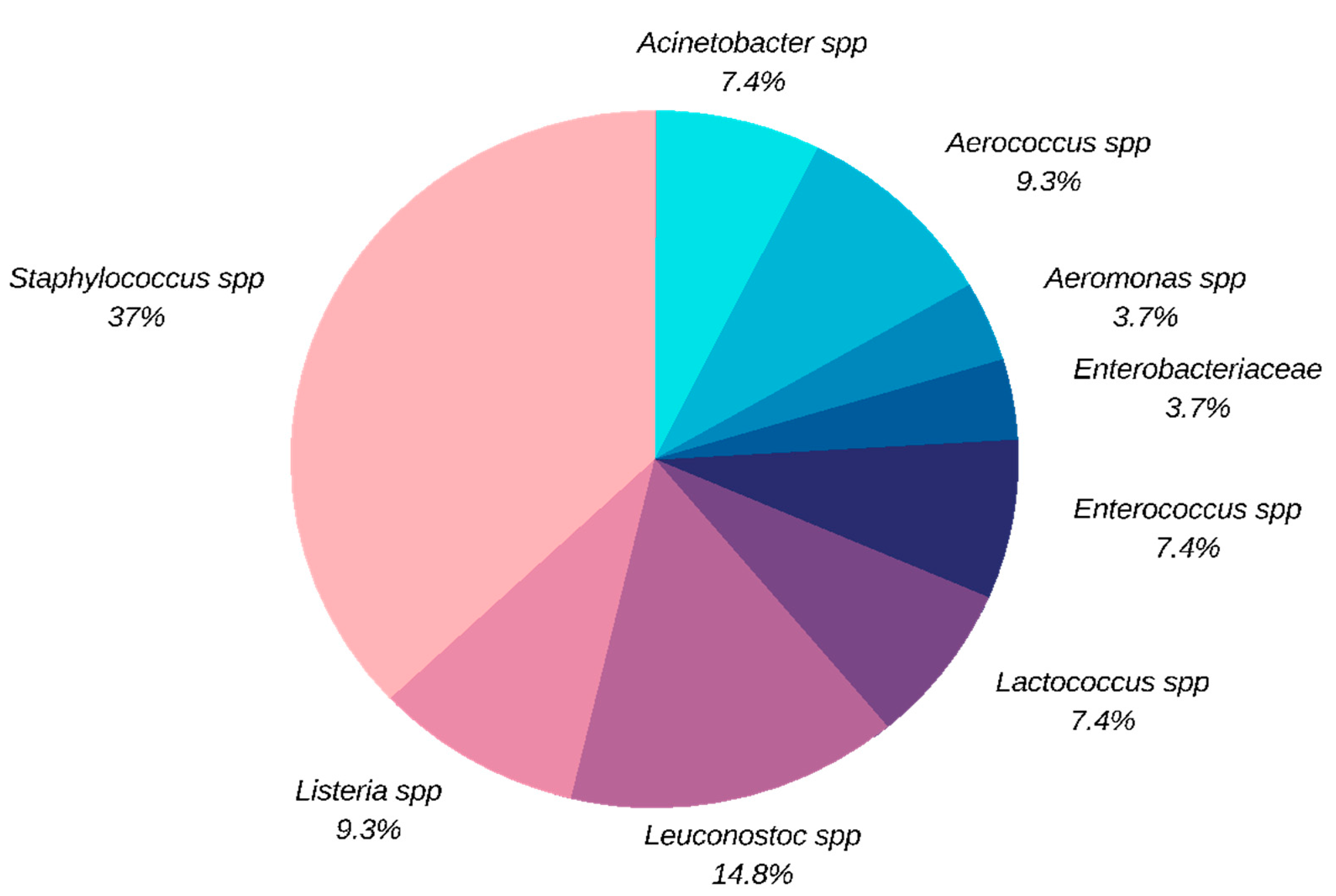
3.1. Quantification and Identification of the Isolates
3.2. Determination of the Minimum Inhibitory Concentration
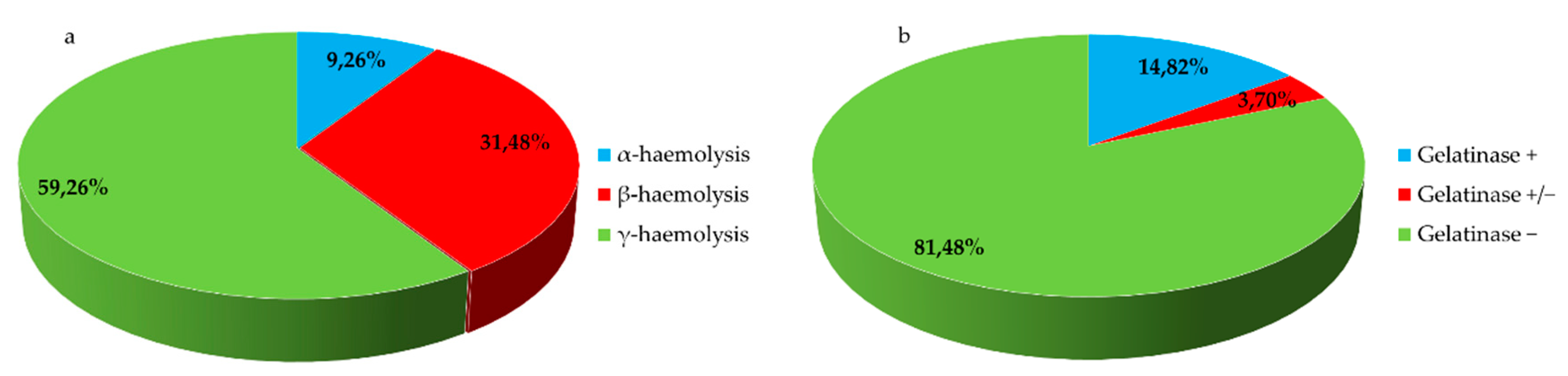
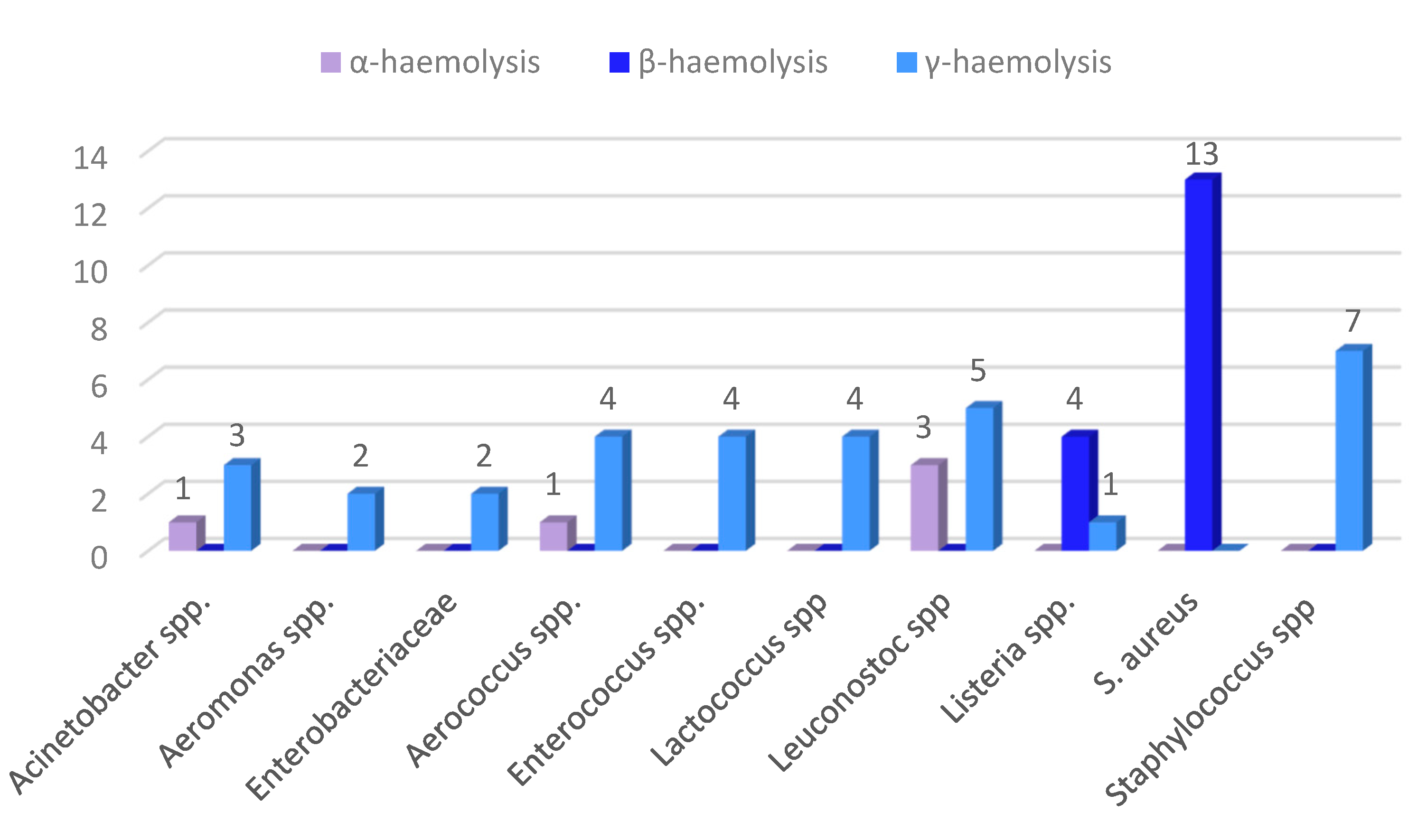
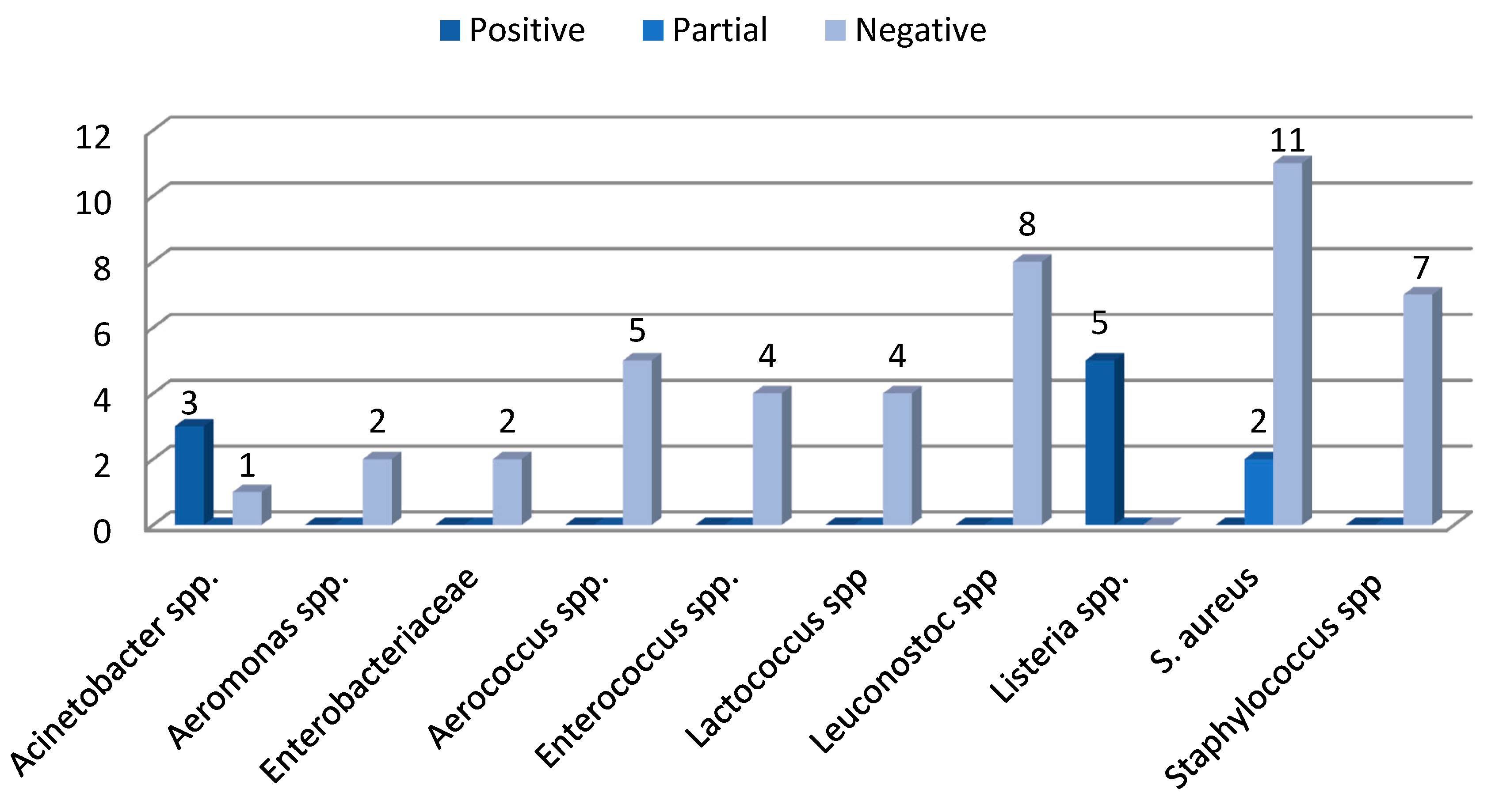
3.3. Phenotypical Virulence Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Callejón, R.M.; Rodríguez-Naranjo, M.I.; Ubeda, C.; Hornedo-Ortega, R.; Garcia-Parrilla, M.C.; Troncoso, A.M. Reported foodborne outbreaks due to fresh produce in the United States and European Union: Trends and causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef]
- Listeriosis in England and Wales: Summary for 2019. Available online: https://www.gov.uk/government/publications/listeria-monocytogenes-surveillance-reports/listeriosis-in-england-and-wales-summary-for-2019 (accessed on 13 July 2021).
- Guerra, M.M.; Mclauchlin, J.; Bernardo, F.A. Listeria in ready-to-eat and unprocessed foods produced in Portugal. Food Microbiol. 2001, 18, 423–429. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez,, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Scientific Opinion on the Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Tompkin, R.B. Control of Listeria monocytogenes in the food-processing environment. J Food Prot. 2002, 65, 709–725. [Google Scholar] [CrossRef]
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillet, C.; Join-Lambert, O.; Le Monnier, A.; Leclercq, A.; Mechaï, F.; Mamzer-Bruneel, M.F.; Bielecka, M.K.; Scortti, M.; Disson, O.; Berche, P.; et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis. 2010, 16, 136–138. [Google Scholar] [CrossRef]
- Hofer, E.; Ribeiro, R.; Feitosa, D.P. Species and serovars of the genus Listeria isolated from different sources in Brazil from 1971 to 1997. Mem. Inst. Oswaldo Cruz. 2000, 95, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Snapir, Y.M.; Vaisbein, E.; Nassar, F. Low virulence but potentially fatal outcome-Listeria ivanovii. Eur. J. Intern. Med. 2006, 17, 286–287. [Google Scholar] [CrossRef] [PubMed]
- EUCAST 2021. European Committee on Antimicrobial Susceptibility Testing European Committee on Antimicrobial Susceptibility Testing (EUCAST) (2021) Clinical-Breakpoints; Version 11.0; Växjö, Sweden. Wiley: Hoboken, NJ, USA, 2021; Available online: https://eucast.org/clinical_breakpoints (accessed on 13 July 2021).
- Davis, B.D. Mechanism of bactericidal action of aminoglycosides. Microbiol. Rev. 1987, 51, 341–350. [Google Scholar] [CrossRef]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.T. Wirkmechanismus der Chinolone [Mechanism of action of quinolones]. Infection 1986, 14, S3–S15. [Google Scholar] [CrossRef]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of quinolone action and resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef]
- Wilkowske, C.J. The penicillins. Mayo Clin. Proc. 1977, 52, 616–624. [Google Scholar] [PubMed]
- Peechakara, B.V.; Basit, H.; Gupta, M. Ampicillin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Frère, J.M. Mechanism of action of beta-lactam antibiotics at the molecular level. Biochem. Pharmacol. 1977, 26, 2203–2210. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Laskin, A.I.; Last, J.A. Tetracyclines. Antibiot. Chemother. 1971, 17, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Shutter, M.C.; Akhondi, H. Tetracycline. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Tanaka, S.; Otaka, T.; Kaji, A. Further studies on the mechanism of erythromycin action. Biochim. Biophys. Acta 1973, 331, 128–140. [Google Scholar] [CrossRef]
- Vázquez-Laslop, N.; Mankin, A.S. How Macrolide Antibiotics Work. Trends Biochem. Sci. 2018, 43, 668–684. [Google Scholar] [CrossRef]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved glycopeptide antibacterial drugs: Mechanism of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Preuss, C.V.; Bernice, F. Vancomycin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- European Food Safety Authority, European Centre for Disease Prevention and Control. The European Union Summary report on trends and sources of zoonoses, zoonotic agents and foodborne outbreaks in 2013. EFSA J. 2015, 13, 3991. [Google Scholar]
- El-Shenawy, M.; El-Shenawy, M.; Mañes, J.; Soriano, J.M. Listeria spp. in street-vended Ready-to-Eat foods. Interdiscip. Perspect. Infect. Dis. 2011, 2011, 968031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossu, F.; Spanu, C.; Deidda, S.; Mura, E.; Casti, D.; Pala, C.; Lamon, S.; Spanu, V.; Ibba, M.; Marrocu, E.; et al. Listeria Spp. and Listeria monocytogenes contamination in Ready-To-Eat sandwiches collected from vending machines. Ital. J. Food Saf. 2016, 5, 5500. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. 2005, 338, 1–26. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32005R2073 (accessed on 13 July 2021).
- Jang, H.G.; Kim, N.H.; Choi, Y.M.; Rhee, M.S. Microbiological quality and risk factors related to sandwiches served in bakeries, cafe’s and sandwich bars in South Korea. J. Food Protect. 2013, 76, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.; Moura, A.T.; García-Díez, J.; Oliveira, I.; Esteves, A.; Saraiva, C. Evaluation of hygienic quality of food served in Universities canteens of northem Portugal. Indian J. Microbiol. 2020, 60, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Fijałkowski, K.; Peitler, D.; Karakulska, J. Staphylococci isolated from ready-to-eat meat—Identification, antibiotic resistance and toxin gene profile. Int. J. Food Microbiol. 2016, 238, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Ordóñez, A.; Leong, D.; Morgan, C.A.; Hill, C.; Gahan, C.G.; Jordan, K. Occurrence, persistence, and virulence potential of Listeria ivanovii in foods and food processing environments in the Republic of Ireland. Biomed. Res. Int. 2015, 2015, 350526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwu, C.D.; Okoh, A.I. Preharvest transmission routes of fresh produce associated bacterial pathogens with outbreak potentials: A Review. Int. J. Environ. Res. Public Health 2019, 16, 4407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guern, A.S.; Martin, L.; Savin, C.; Carniel, E. Yersiniosis in France: Overview and potential sources of infection. Int. J. Infect. Dis. 2016, 46, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. The significance of mesophilic Aeromonas spp. in minimally processed Ready-to-Eat seafood. Microorganisms 2019, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, A.M.; Nascimento, J.D. Acinetobacter: An underrated foodborne pathogen? J. Infect. Dev. Ctries. 2017, 11, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selleck, E.M.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Bondi, M.; Laukova, A.; de Niederhausern, S.; Messi, P.; Papadopoulou, C.; Economou, V. Controversial aspects displayed by Enterococci: Probiotics or pathogens? Biomed. Res. Int. 2020, 2020, 9816185. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Enterococci from foods. FEMS Microbiol. Rev. 2002, 26, 163–171. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public. Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Lin, Y.T.; Wan, T.W.; Wang, D.Y.; Lin, H.Y.; Lin, C.Y.; Chen, Y.C.; Teng, L.J. Distribution of antibiotic resistance genes among Staphylococcus species isolated from ready-to-eat foods. J. Food Drug. Anal. 2019, 27, 841–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.; Savage, V.M.; Yeh, P.J. Intermediate levels of antibiotics may increase diversity of colony size phenotype in bacteria. Comput. Struct. Biotechnol. J. 2018, 16, 307–315. [Google Scholar] [CrossRef]
- Devlieghere, F.; Vermeiren, L.; Debevere, J. New preservation technologies: Possibilities and limitations. Int. Dairy J. 2004, 14, 273–285. [Google Scholar] [CrossRef]
- Antonio, C.M.; Abriouel, H.; López, R.L.; Omar, N.B.; Valdivia, E.; Gálvez, A. Enhanced bactericidal activity of enterocin AS-48 in combination with essential oils, natural bioactive compounds and chemical preservatives against Listeria monocytogenes in ready-to-eat salad. Food Chem. Toxicol. 2009, 47, 2216–2223. [Google Scholar] [CrossRef]
- Ben-Fadhel, Y.; Maherani, B.; Aragones, M.; Lacroix, M. Antimicrobial properties of encapsulated antimicrobial natural plant products for Ready-to-Eat carrots. Foods. 2019, 8, 535. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Regulation on the hygiene of foodstuffs, 852/2004/EC. Off. J. L 2004, 139, 54. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents | Group | Mechanism of Action | References |
|---|---|---|---|
| Amikacin | Aminoglycoside | Inhibits bacterial protein synthesis and also affects the barrier function of bacterial cell membranes. | [11,12] |
| Ciprofloxacin | Quinolone | Inhibit bacterial DNA gyrase and interfere with bacterial DNA replication. | [13,14] |
| Ampicillin | Penicillin (aminopenicillin) | Inhibition of bacterial cell wall synthesis by binding one or more of the penicillin binding proteins (PBPs). | [15,16] |
| Oxacillin | β-Lactam | Inhibits the synthesis of bacterial cell walls. | [17,18] |
| Imipenem | β-Lactam | Inhibits the synthesis of bacterial cell walls. | [17,18] |
| Tetracycline | Tetracycline | It specifically binds to the bacterial ribosomal 30S subunit at the A position, prevents the connection of aminoacyl-tRNA at this position, and inhibits protein synthesis. | [19,20] |
| Erythromycin | Macrolide | Combines with bacterial ribosomal 50S subunit to inhibit peptide acyltransferase and bacterial protein synthesis. | [21,22] |
| Vancomycin | Glycopeptide | Inhibits the synthesis of bacterial cell walls. | [23,24] |
| Strains | Ciprofloxacin | Tetracycline | Ampicillin | Erythromycin | Vancomycin |
|---|---|---|---|---|---|
| E. faecium EC1 | 4 µg/mL | 1 µg/mL | 16 µg/mL | 16 µg/mL | 8 µg/mL |
| E. faecium EC2 | 4 µg/mL | 1 µg/mL | 4 µg/mL | 4 µg/mL | 8 µg/mL |
| E. avium EC16 | / | / | 1 µg/mL | / | 2 µg/mL |
| E. durans EC17 | / | / | 2 µg/mL | / | 2 µg/mL |
| BREAKPOINT | ≥8 µg/mL | ≥4 µg/mL | ≥8 µg/mL | ≥4 µg/mL | ≥4 µg/mL |
| Strains | Amikacin | Ciprofloxacin | Erythromicin | Vancomiycin | Oxacillin |
|---|---|---|---|---|---|
| S. aureus S26 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 1 µg/mL | 4 µg/mL |
| S. aureus S28 | 1 µg/mL | 1 µg/mL | 1 µg/mL | 1 µg/mL | 2 µg/mL |
| S. aureus S29 | 1 µg/mL | 0.25 µg/mL | 0.5 µg/mL | 0.5 µg/mL | 2 µg/mL |
| S. aureus S31 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S32 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 4 µg/mL |
| S. aureus S33 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S34 | 1 µg/mL | 1 µg/mL | 1 µg/mL | 2µg/mL | 2 µg/mL |
| S. aureus S35 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S36 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S37 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S41 | 1 µg/mL | 0.25 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S42 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 1 µg/mL | 1 µg/mL |
| S. aureus S43 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 2 µg/mL |
| BREAKPOINT | ≥16 µg/mL | ≥1 µg/mL | ≥1 µg/mL | ≥2 µg/mL | ≥2 µg/mL |
| Strains | Tetracycline | Ampicillin | Erythromycin |
|---|---|---|---|
| L. ivanovii spp. londoniensis 46L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 47L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 48L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 49L | 1 µg/mL | 0.5 µg/mL | 2 µg/mL |
| L. welshimeri 50L | 4 µg/mL | 4 µg/mL | 2 µg/mL |
| BREAKPOINT | ≥1 µg/mL | ≥1 µg/mL | ≥1 µg/mL |
| Strain | Imipenem | Ciprofloxacin | Tetracyclin |
|---|---|---|---|
| A. hydrophila 52EB | 2 µg/mL | ||
| A. hydrophila 59EB | 2 µg/mL | ||
| BREAKPOINT | ≥1 µg/mL | ||
| Y. enterocolitica 58EB | 0.25 µg/mL | 4 µg/mL | |
| BREAKPOINT | ≥0.25 µg/mL | ≥4 µg/mL |
| Strains | Amikacin | Ciprofloxacin |
|---|---|---|
| A. Iwofii 54EB | 2 µg/mL | 0.25 µg/mL |
| A. calcoaceticus 55EB | 2 µg/mL | 0.25 µg/mL |
| A. Iwofii 56EB | 2 µg/mL | 0.25 µg/mL |
| A. Iwofii 57EB | 2 µg/mL | 0.25 µg/mL |
| BREAKPOINT | ≥8 µg/mL | ≥1 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camellini, S.; Iseppi, R.; Condò, C.; Messi, P. Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Appl. Sci. 2021, 11, 7177. https://doi.org/10.3390/app11167177
Camellini S, Iseppi R, Condò C, Messi P. Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Applied Sciences. 2021; 11(16):7177. https://doi.org/10.3390/app11167177
Chicago/Turabian StyleCamellini, Stefania, Ramona Iseppi, Carla Condò, and Patrizia Messi. 2021. "Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors" Applied Sciences 11, no. 16: 7177. https://doi.org/10.3390/app11167177
APA StyleCamellini, S., Iseppi, R., Condò, C., & Messi, P. (2021). Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Applied Sciences, 11(16), 7177. https://doi.org/10.3390/app11167177