
Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Sample Preparation
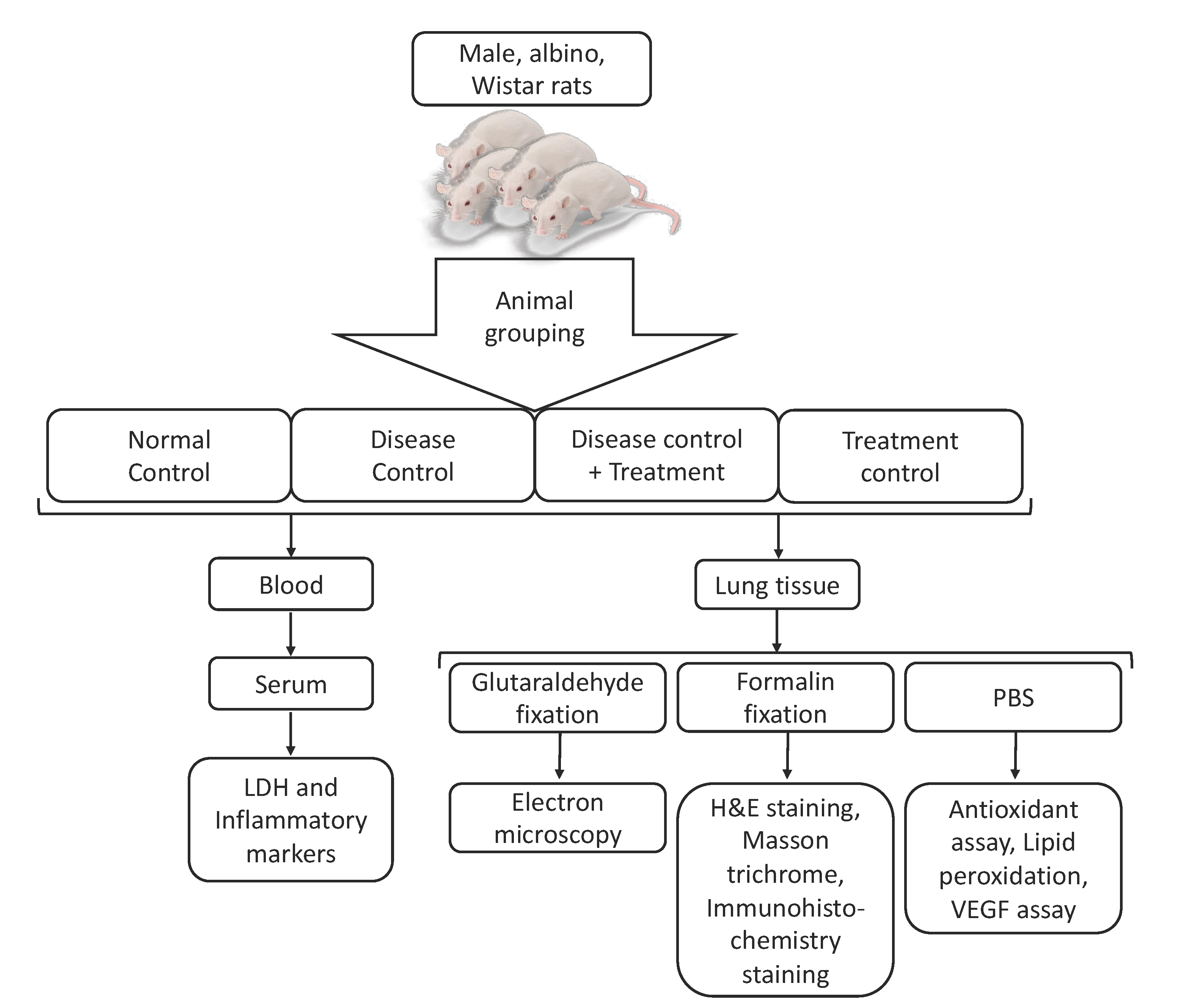
2.4. Animal Grouping and Treatment Plan
2.5. Measurement of Inflammatory Marker
2.6. Lactate Dehydrogenase (LDH) Assay
2.7. Tissue Sampling
2.8. Measurement of Lipid Peroxidation
2.9. Measurement of Antioxidant Enzymes
2.10. Angiogenic Marker Measurement
2.11. Histopathological Evaluation
2.12. Ultrastructural Assessment
2.13. Immunohistochemical Staining
2.14. Statistical Analysis
3. Results
3.1. The Effect of Quercetin on LDH Levels
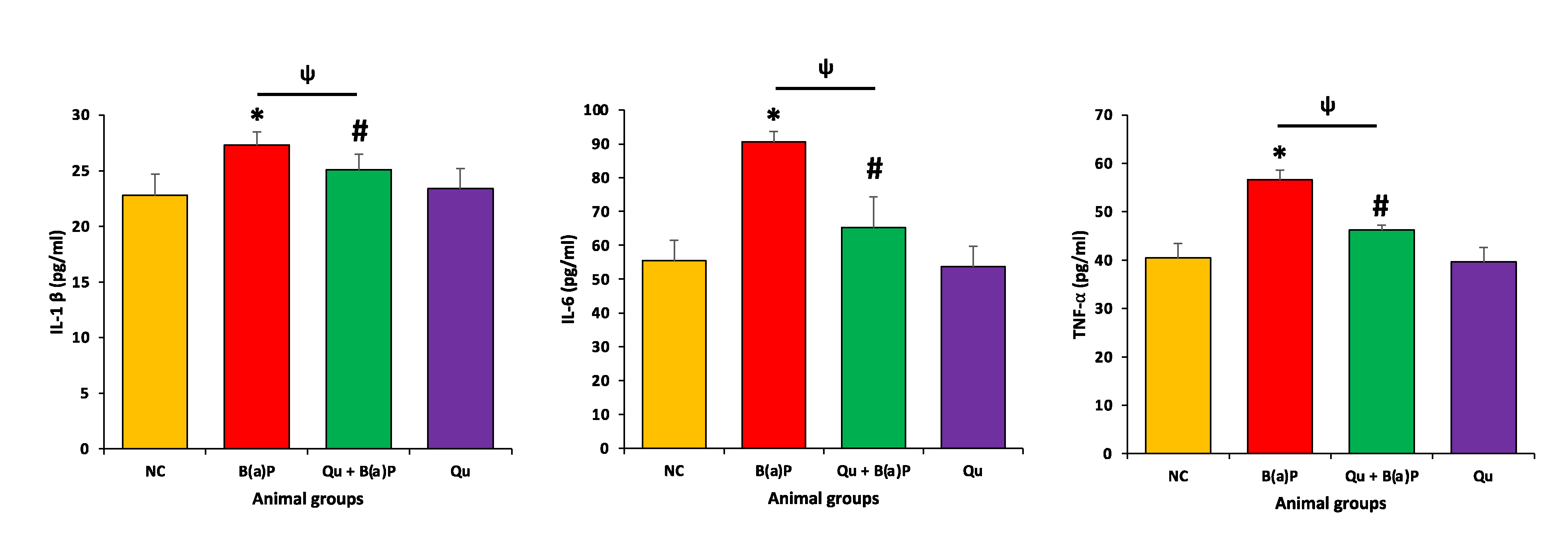
3.2. Effects of Qu on Inflammatory Markers
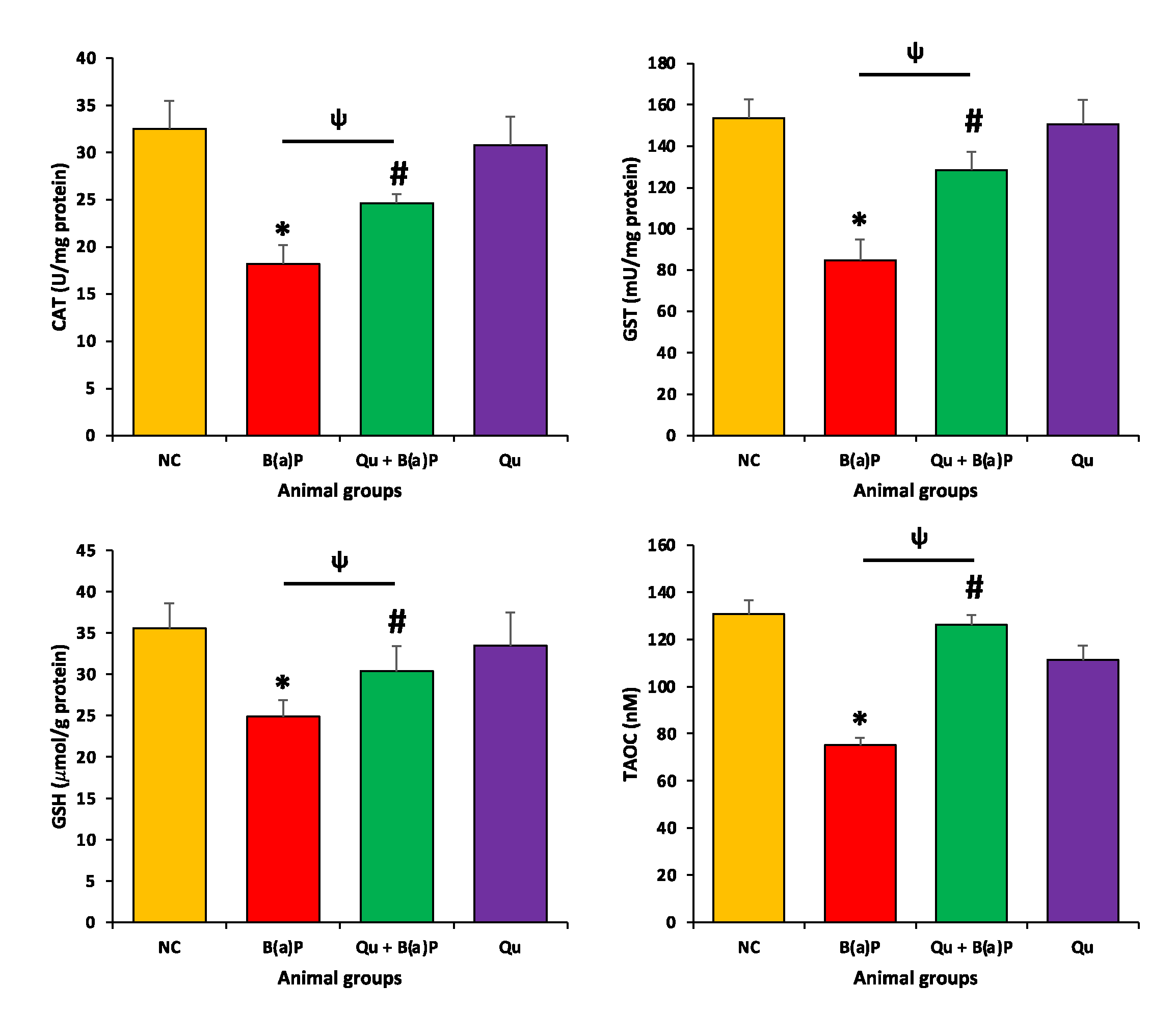
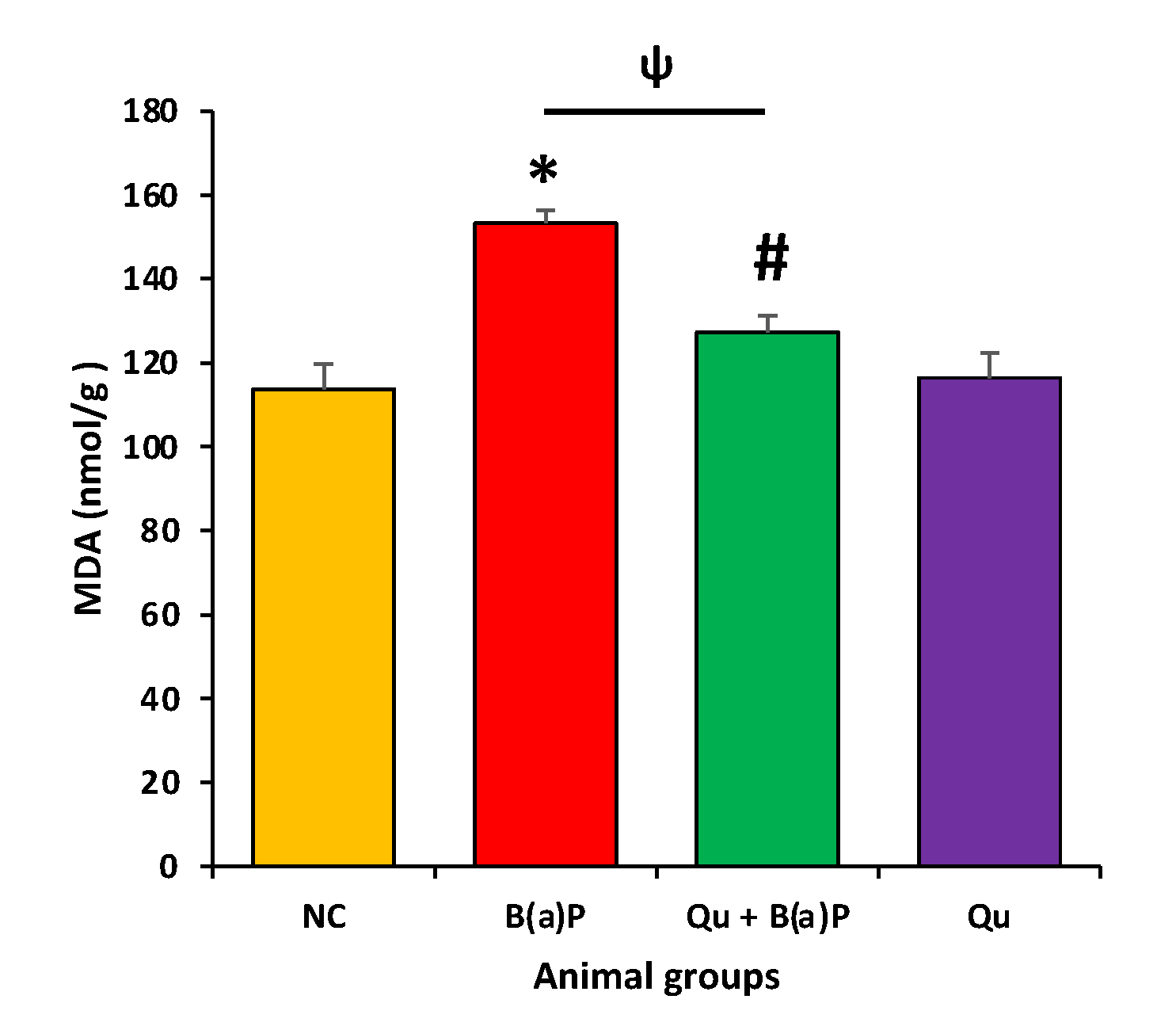
3.3. Quercetin Effect on Antioxidant Enzymes and Malondialdehyde (MDA)
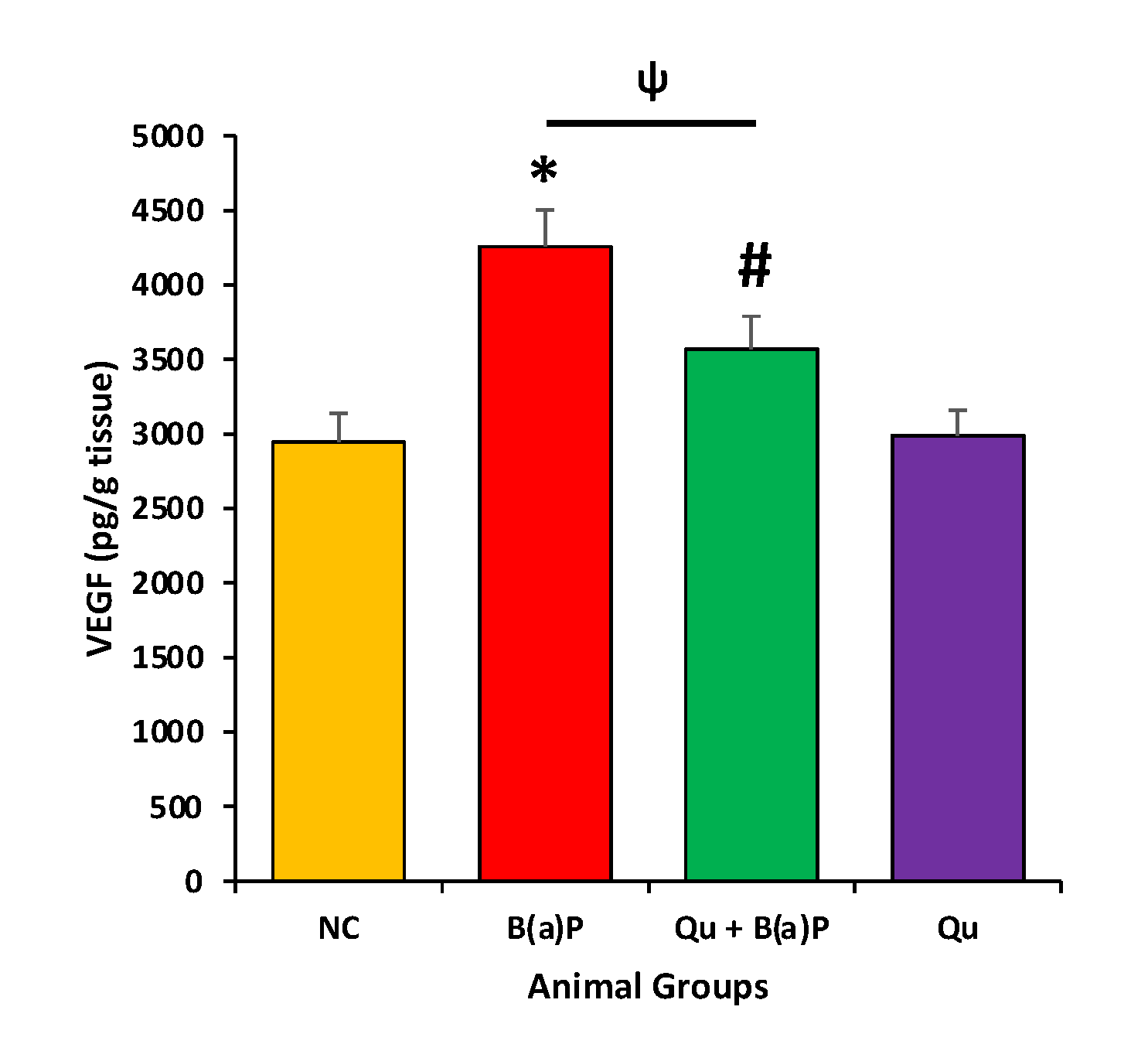
3.4. Effect of Quercetin on VEGF
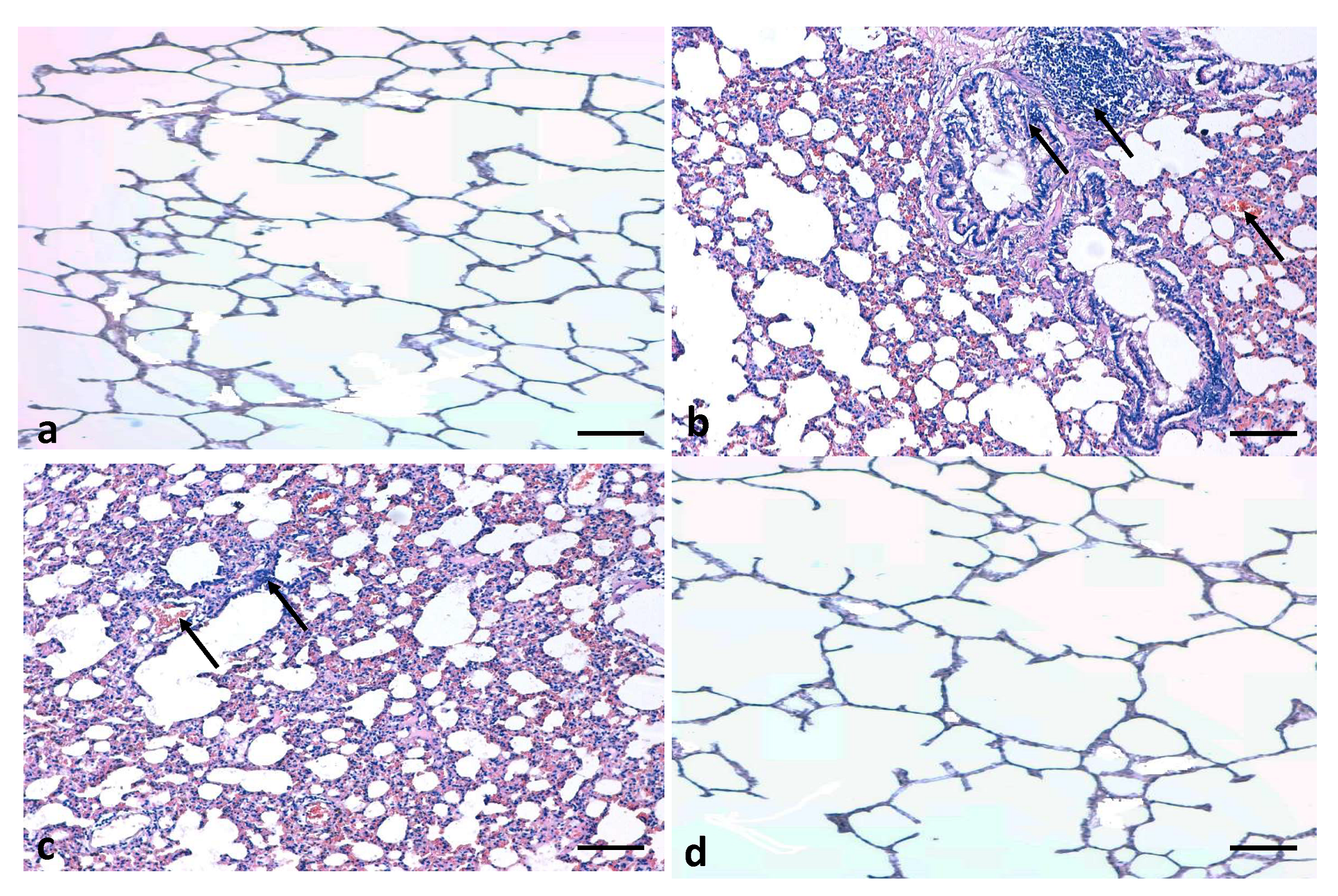
3.5. Effects of Quercetin on Lung Histology
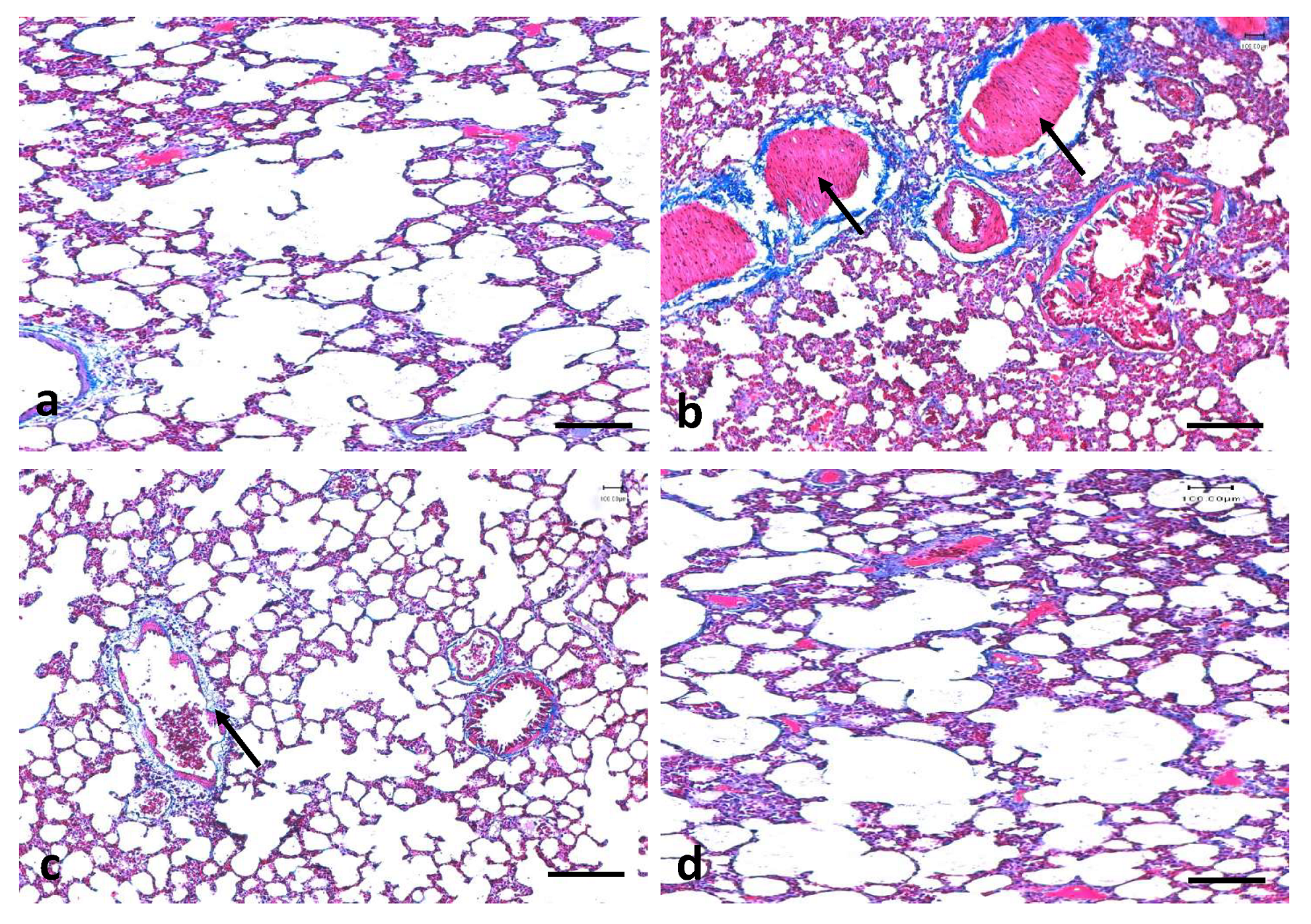
3.6. Effects of Quercetin on Lung Fibrosis
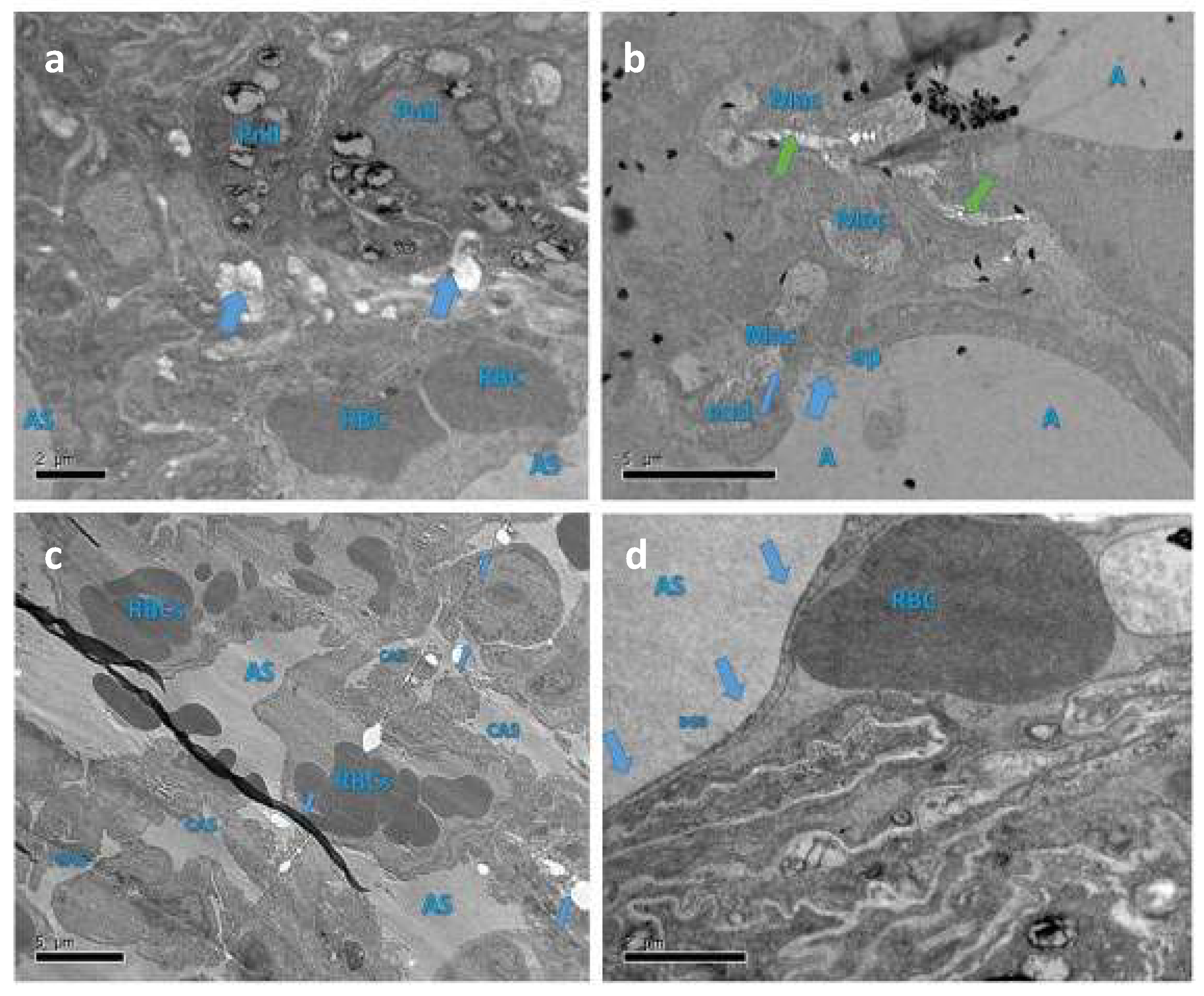
3.7. Protective Role of Qu on Lung Tissue Ultrastructure
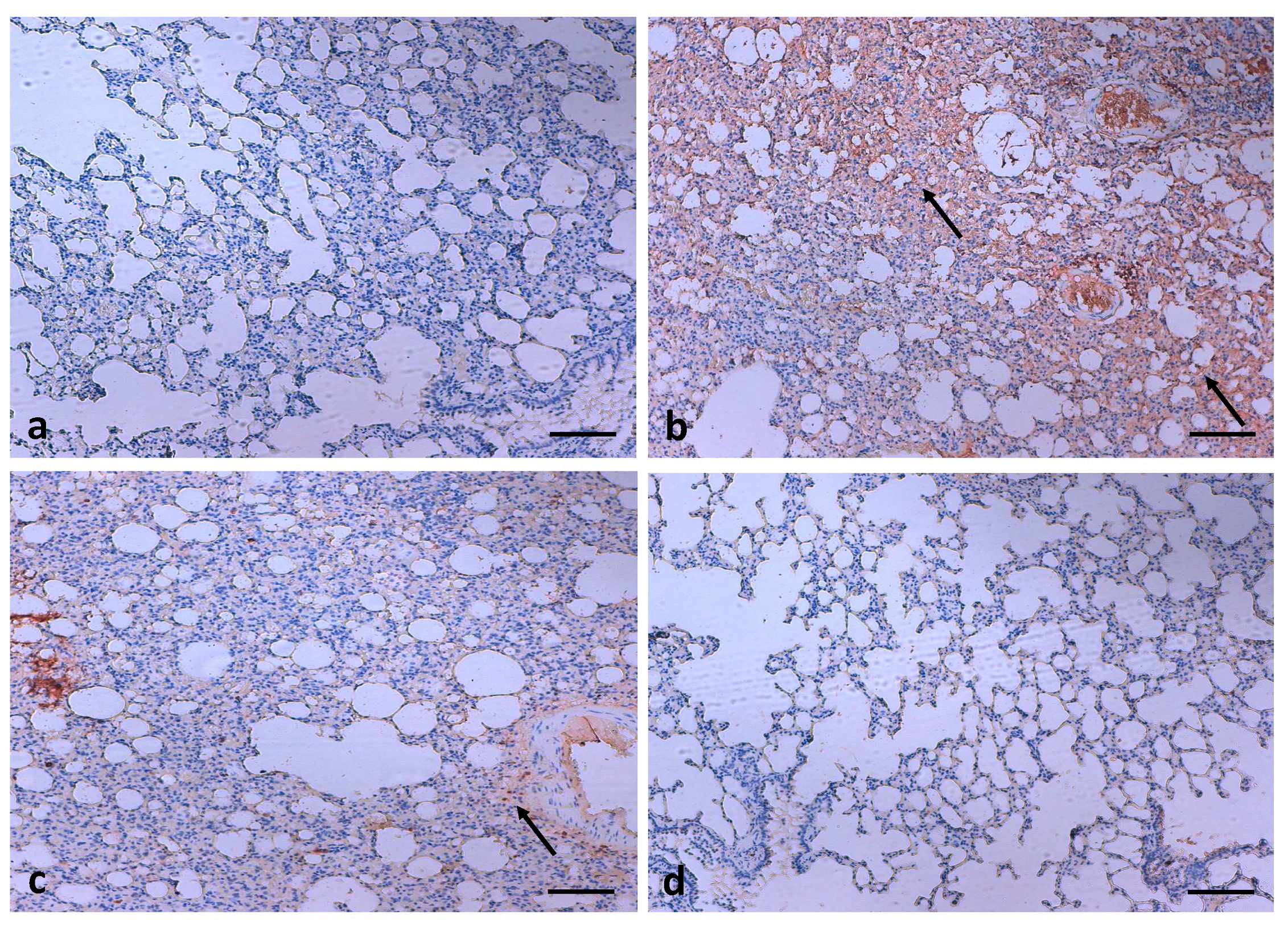
3.8. Effects of Quercetin on COX-2 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Organization WHO. WHO Report on the Global Tobacco Epidemic, 2017: Monitoring Tobacco use and Prevention Policies; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Organization WHO. WHO Report on the Global Tobacco Epidemic, 2011: Warning About the Dangers of Tobacco; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Islami, F.; Torre, L.A.; Jemal, A. Global trends of lung cancer mortality and smoking prevalence. Transl. Lung. Cancer Res. 2015, 4, 327–338. [Google Scholar]
- Szkup, M.; Jurczak, A.; Karakiewicz, B.; Kotwas, A.; Kopeć, J.; Grochans, E. Influence of cigarette smoking on hormone and lipid metabolism in women in late reproductive stage. Clin. Interv. Aging. 2018, 13, 109. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Peto, R.; Zhou, M.; Iona, A.; Smith, M.; Yang, L.; Guo, Y.; Chen, Y.; Bian, Z.; Lancaster, G.; et al. Contrasting male and female trends in tobacco-attributed mortality in China: Evidence from successive nationwide prospective cohort studies. Lancet 2015, 386, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Pirie, K.; Peto, R.; Reeves, G.K.; Green, J.; Beral, V.; Collaborators MWS. The 21st century hazards of smoking and benefits of stopping: A prospective study of one million women in the UK. Lancet 2013, 381, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Doll, R.; Peto, R.; Boreham, J.; Sutherland, I. Mortality in relation to smoking: 50 years’ observations on male British doctors. BMJ 2004, 328, 1519. [Google Scholar] [CrossRef] [Green Version]
- Doll, R.; Hill, A.B. The mortality of doctors in relation to their smoking habits. BMJ 1954, 1, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, D.H. Polycyclic aromatic hydrocarbons in the diet. Mutat. Res. 1999, 443, 139–147. [Google Scholar] [CrossRef]
- Jahangir, T.; Sultana, S. Benzo(a)pyrene induced genotoxicity: Attenuation by farnesol in a mouse model. J. Enzym. Inhib. Med. Chem. 2008, 23, 888–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, J.; Shree, A.; Vafa, A.; Afzal, S.M.; Sultana, S. Taxifolin ameliorates Benzo[a]pyrene-induced lung injury possibly via stimulating the Nrf2 signalling pathway. Int. Immunopharmacol. 2021, 96, 107566. [Google Scholar] [CrossRef]
- Baumgartner, K.B.; Samet, J.M.; Stidley, C.A.; Colby, T.V.; Waldron, J.A. Cigarette smoking: A risk factor for idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care. Med. 1997, 155, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Qamar, W.; Khan, A.Q.; Khan, R.; Lateef, A.; Tahir, M.; Rehman, M.U.; Ali, F.; Sultana, S. Benzo(a)pyrene-induced pulmonary inflammation, edema, surfactant dysfunction, and injuries in rats: Alleviation by farnesol. Exp. Lung. Res. 2012, 38, 19–27. [Google Scholar] [CrossRef]
- Diwanji, N.; Bergmann, A. An unexpected friend—ROS in apoptosis-induced compensatory proliferation: Implications for regeneration and cancer. Semin. Cell. Dev. Biol. 2018, 80, 74–82. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.H.; Chu, J.P.; Jiang, M.H.; Baek, S.H.; Park, H.D. Effects of fraction obtained from Korean Corni Fructus extracts causing anti-proliferation and p53-dependent apoptosis in A549 lung cancer cells. Nutr. Cancer 2011, 63, 121–129. [Google Scholar] [CrossRef]
- Ratovitski, E. Anticancer natural compounds as epigenetic modulators of gene expression. Curr. Genom. 2017, 18, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Sreekala, C.; Zhang, Z.; Budhraja, A.; Ding, S.; Son, Y.O.; Wang, X.; Hitron, A.; Hyun-Jung, K.; Wang, L.; et al. Cancer prevention with promising natural products: Mechanisms of action and molecular targets. Anti Cancer Agents Med. Chem. 2012, 12, 1159–1184. [Google Scholar] [CrossRef] [PubMed]
- Costea, T.; Hudiță, A.; Ciolac, O.A.; Gălățeanu, B.; Ginghină, O.; Costache, M.; Ganea, C.; Mocanu, M.M. Chemoprevention of colorectal cancer by dietary compounds. Int. J. Mol. Sci. 2018, 19, 3787. [Google Scholar] [CrossRef] [Green Version]
- Rostock, M.; Naumann, J.; Guethlin, C.; Guenther, L.; Bartsch, H.H.; Walach, H. Classical homeopathy in the treatment of cancer patients—A prospective observational study of two independent cohorts. BMC Cancer 2011, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Wang, Y.; Gao, L. Mechanism of antioxidant properties of quercetin and quercetin-DNA complex. J. Mol. Model. 2020, 26, 133. [Google Scholar] [CrossRef]
- Saeedi-Boroujeni, A.; Mahmoudian-Sani, M.R. Anti-inflammatory potential of Quercetin in COVID-19 treatment. J. Inflamm. 2021, 18, 3. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and cancer: New insights into its therapeutic effects on ovarian cancer cells. Cell. Biosci. 2020, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.H.; An, J.Y.; Kwon, Y.T.; Rhee, J.G.; Lee, Y.J. Effects of low dose quercetin: Cancer cell-specific inhibition of cell cycle progression. J. Cell. Biochem. 2009, 106, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, M.X.; Deng, X.H.; Ai, F.; Yuan, G.Y.; Song, H.Y. Effect of quercetin on the proliferation of the human ovarian cancer cell line SKOV-3 in vitro. Exp. Ther. Med. 2015, 10, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.K.; Mousa, A.M.; El-Sammad, N.M.; Abdel-Halim, A.H.; Khalil, W.K.; Elsayed, E.A.; Anwar, N.; Linscheid, M.W.; Moustafa, E.S.; Hashim, A.N.; et al. Antitumor activity of Cuphea ignea extract against benzo(a)pyrene-induced lung tumorigenesis in Swiss Albino mice. Toxicol. Rep. 2019, 6, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Kumar, B.; Sagi, S.S. Prophylactic efficacy of Quercetin in ameliorating the hypoxia induced vascular leakage in lungs of rats. PLoS ONE 2019, 14, e0219075. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M.A. Principles and Techniques of Electron. Microscopy: Biological Applications, 4th ed.; Cambridge University Press: Edinburg, UK, 2000; pp. 37–59. [Google Scholar]
- Rahmani, A.H.; Babiker, A.Y.; AlWanian, W.M.; Elsiddig, S.A.; Faragalla, H.E.; Aly, S.M. Association of cytokeratin and vimentin protein in the genesis of transitional cell carcinoma of urinary bladder patients. Dis. Markers 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, A.; Alzohairy, M.; Khadri, H.; Mandal, A.K.; Rizvi, M.A. Expressional evaluation of vascular endothelial growth factor (VEGF) protein in urinary bladder carcinoma patients exposed to cigarette smoke. Int. J. Clin. Exp. Pathol. 2012, 5, 195. [Google Scholar] [PubMed]
- Rahmani, A.; Alzohairy, M.; Babiker, A.Y.; Rizvi, M.A.; Elkarimahmad, H.G. Clinicopathological significance of PTEN and bcl2 expressions in oral squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2012, 5, 965. [Google Scholar]
- Wang, C.C.; Yuan, J.R.; Wang, C.F.; Yang, N.; Chen, J.; Liu, D.; Song, J.; Feng, L.; Tan, X.B.; Jia, X.B. Anti-inflammatory effects of Phyllanthus emblica L on benzopyrene-induced precancerous lung lesion by regulating the IL-1β/miR-101/Lin28B signaling pathway. Integr. Cancer Therapies 2017, 16, 505–515. [Google Scholar] [CrossRef]
- Gaweł, S.; Wardas, M.; Niedworok, E.; Wardas, P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek. 2004, 57, 453–455. [Google Scholar] [PubMed]
- Oke, G.O.; Abiodun, A.A.; Imafidon, C.E.; Monsi, B.F. Zingiber officinale (Roscoe) mitigates CCl4-induced liver histopathology and biochemical derangements through antioxidant, membrane-stabilizing and tissue-regenerating potentials. Toxicol. Rep. 2019, 6, 416–425. [Google Scholar] [CrossRef] [PubMed]
- ParathodiIllam, S.; Hussain, A.; Elizabeth, A.; Narayanankutty, A.; Raghavamenon, A.C. Natural combination of phenolic glycosides from fruits resists pro-oxidant insults to colon cells and enhances intrinsic antioxidant status in mice. Toxicol. Rep. 2019, 6, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Barnwal, P.; Vafa, A.; Afzal, S.M.; Shahid, A.; Hasan, S.K.; Alpashree Sultana, S. Benzo(a)pyrene induces lung toxicity and inflammation in mice: Prevention by carvacrol. Hum. Exp. Toxicol. 2018, 37, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, S.; Gao, C.; Deng, H.; Liu, Y.; Li, C.; Yuan, L.; Luo, Y. Isoorientin attenuates benzo [a] pyrene-induced liver injury by inhibiting autophagy and pyroptosis in vitro and vivo. Food Agric. Immunol. 2019, 30, 841–861. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Dang, F.; Gao, J.; Zhao, H.; Qi, S.; Gao, M. Acute benzo[a]pyrene treatment causes different antioxidant response and DNA damage in liver, lung, brain, stomach and kidney. Heliyon 2018, 4, e00898. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.; Shahid, A.; Ali, N.; Hasan, S.K.; Majed, F.; Sultana, S. Amelioration of benzo[a]pyrene-induced oxidative stress and pulmonary toxicity by naringenin in Wistar rats: A plausible role of COX-2 and NF-kappab. Hum. Exp. Toxicol. 2017, 36, 349–364. [Google Scholar] [CrossRef]
- Rahimi, V.B.; Shirazinia, R.; Fereydouni, N.; Zamani, P.; Darroudi, S.; Sahebkar, A.H.; Askari, V.R. Comparison of honey and dextrose solution on post-operative peritoneal adhesion in rat model. Biomed. Pharmacother. 2017, 92, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, Y.; Dong, L.; Wang, Z.; Chen, L.; Liang, D.; Shi, D.; Shan, X.; Liang, G. Design, Synthesis, and Biological Evaluation of Novel Quinazoline Derivatives as Anti-inflammatory Agents against Lipopolysaccharide-induced Acute Lung Injury in Rats. Chem. Biol. Drug Des. 2015, 85, 672–684. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev. 2007, 65, S140–S146. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhong, T.; Wu, H. Quercetin protects against lipopolysaccharide-induced acute lung injury in rats through suppression of inflammation and oxidative stress. Arch. Med. Sci. 2015, 11, 427–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almatroodi, S.A.; Alrumaihi, F.; Alsahli, M.A.; Alhommrani, M.F.; Khan, A.; Rahmani, A.H. Curcumin, an Active Constituent of Turmeric Spice: Implication in the Prevention of Lung Injury Induced by Benzo(a) Pyrene (BaP) in Rats. Molecules 2020, 25, 724. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, N.; Suresh, G.; Ramesh, B.; Siva, G.V. Fisetin, a novel flavonol attenuates benzo(a)pyrene-induced lung carcinogenesis in Swiss albino mice. Food Chem Toxicol. 2011, 49, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Islam, J.; Shree, A.; Afzal, S.M.; Vafa, A.; Sultana, S. Protective effect of Diosmin against benzo(a)pyrene-induced lung injury in Swiss Albino Mice. Environ. Toxicol. 2020, 35, 747–757. [Google Scholar] [CrossRef]
- Yeo, C.D.; Kim, Y.A.; Lee, H.Y.; Kim, J.W.; Kim, S.J.; Lee, S.H.; Kim, Y.K. Roflumilast treatment inhibits lung carcinogenesis in benzo (a) pyrene-induced murine lung cancer model. Eur. J. Pharmacol. 2017, 812, 189–195. [Google Scholar] [CrossRef]
- Li, F.; Bai, Y.; Zhao, M.; Huang, L.; Li, S.; Li, X.; Chen, Y. Quercetin inhibits vascular endothelial growth factor-induced choroidal and retinal angiogenesis in vitro. Ophthalmic. Res. 2015, 53, 109–116. [Google Scholar] [CrossRef]
- Alzohairy, M.A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effects of Thymoquinone, an Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation. Molecules 2021, 26, 3218. [Google Scholar] [CrossRef]
- Anandakumar, P.; Kamaraj, S.; Jagan, S.; Ramakrishnan, G.; Asokkumar, S.; Naveenkumar, C.; Raghunandhakumar, S.; Vanitha, M.K.; Devaki, T. The Anticancer Role of Capsaicin in Experimentally induced Lung Carcinogenesis. J. Pharmacopunct. 2015, 18, 19–25. [Google Scholar] [CrossRef]
- Shahid, A.; Ali, R.; Ali, N.; Hasan, S.K.; Rashid, S.; Majed, F.; Sultana, S. Attenuation of genotoxicity, oxidative stress, apoptosis and inflammation by rutin in benzo (a) pyrene exposed lungs of mice: Plausible role of NF-κB, TNF-α and Bcl-2. J. Complementary Integr. Med. 2016, 13, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, J.; Wang, B.; Wu, D.; Li, H.; Lu, H.; Wu, H.; Chai, Y. Protective effect of quercetin on lipopolysaccharide-induced acute lung injury in mice by inhibiting inflammatory cell influx. Exp. Biol. Med. 2014, 239, 1653–1662. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
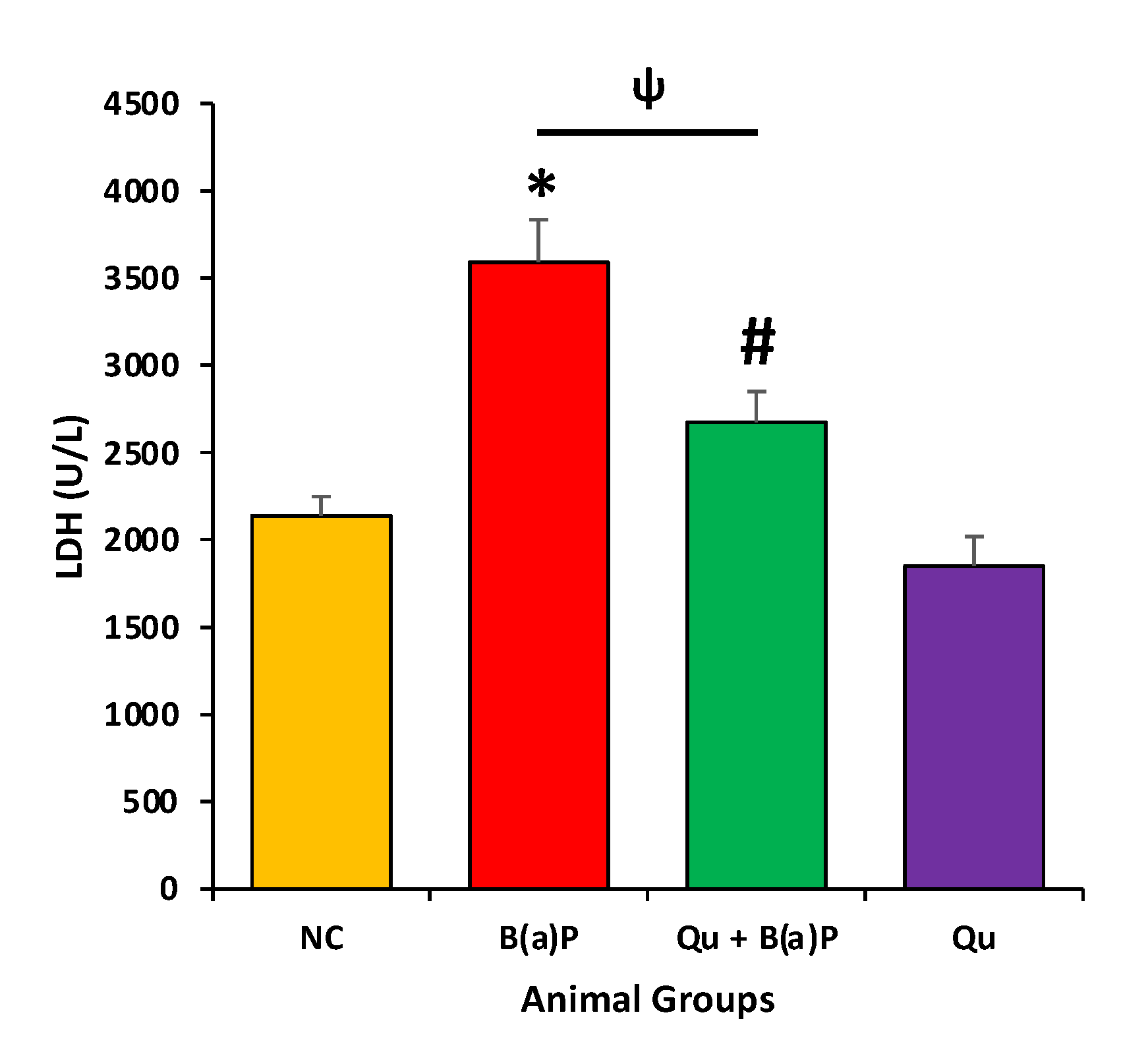{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Number | Group Description | Short Name | Treatment Plan |
|---|---|---|---|
| 1 | Normal Control | NC | The animals received normal saline solution (400 μL) through oral gavage for 8 consecutive weeks |
| 2 | Disease control | B(a)P | Oral administration of B(a)P in olive oil (50 mg/kg b.w.) (400 μL) 3 times/week for 8 consecutive weeks [29]. |
| 3 | Disease control + treatment | Qu + B(a)P | Qu (50 mg/kg b.w.), 400 μL was given orally 30 min before the oral administration of B(a)P in olive oil (50 mg/kg b.w.) 3 times/week for 8 consecutive weeks [30] |
| 4 | Treatment control | Qu | The rats received Qu (400 μL) 3 times a week at a dose of 50 mg/kg b.w. for 8 consecutive weeks |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzohairy, M.A.; Khan, A.A.; Ansari, M.A.; Babiker, A.Y.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule. Appl. Sci. 2021, 11, 8675. https://doi.org/10.3390/app11188675
Alzohairy MA, Khan AA, Ansari MA, Babiker AY, Alsahli MA, Almatroodi SA, Rahmani AH. Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule. Applied Sciences. 2021; 11(18):8675. https://doi.org/10.3390/app11188675
Chicago/Turabian StyleAlzohairy, Mohammad A., Amjad Ali Khan, Mohammad Azam Ansari, Ali Yousif Babiker, Mohammed A. Alsahli, Saleh A. Almatroodi, and Arshad Husain Rahmani. 2021. "Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule" Applied Sciences 11, no. 18: 8675. https://doi.org/10.3390/app11188675
APA StyleAlzohairy, M. A., Khan, A. A., Ansari, M. A., Babiker, A. Y., Alsahli, M. A., Almatroodi, S. A., & Rahmani, A. H. (2021). Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule. Applied Sciences, 11(18), 8675. https://doi.org/10.3390/app11188675