
Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Historic Overview of Salón de Reinos
2.2. Sampling
2.3. Isolation of Culturable Microorganisms
2.4. DNA Extraction and Amplification
2.5. DNA Sequencing
2.6. Biodeteriorative Activity of the Isolates
3. Results and Discussion
3.1. Ecology of Bacterial Isolates
3.2. Ecology of Fungal Isolates
3.3. Metabolic Activities of Bacterial and Fungal Isolates
3.4. Cleaning and Biocide Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosado, T.; Falé, A.; Gil, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Oxalate biofilm formation in mural paintings due to microorganisms—A comprehensive study. Int. Biodeter. Biodegr. 2013, 85, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rosado, T.; Gil, M.; Caldeira, A.T.; Martins, M.R.; Dias, C.B.; Carvalho, L.; Mirão, J.; Candeias, A.E. Material characterization and biodegradation assessment of mural paintings: Renaissance frescoes from Santo Aleixo Church, Southern Portugal. Int. J. Archit. Herit. 2014, 8, 835–852. [Google Scholar] [CrossRef]
- Rosado, T.; Falé, A.; Gil, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Understanding the influence of microbial contamination on colour alteration of pigments used in wall paintings—The case of red and yellow ochres and ultramarine blue. Color Res. Appl. 2019, 44, 783–789. [Google Scholar] [CrossRef]
- Unković, N.; Grbić, M.L.; Stupar, M.; Savković, Ž.; Jelikić, A.; Stanojević, D.; Vukojević, J. Fungal-induced deterioration of mural paintings: In situ and mock-model microscopy analyses. Microsc. Microanal. 2016, 22, 410–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trovão, J.; Portugal, A. Current knowledge on the fungal degradation abilities profiled through biodeteriorative plate essays. Appl. Sci. 2021, 11, 4196. [Google Scholar] [CrossRef]
- Piñar, G.; Saiz-Jimenez, C.; Schabereiter-Gurtner, C.; Blanco-Varela, M.T.; Lubitz, W.; Rölleke, S. Archaeal communities in two disparate deteriorated ancient wall paintings: Detection, identification and temporal monitoring by DGGE. FEMS Microbiol. Ecol. 2001, 37, 45–54. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Heyrman, J.; Dornieden, T.; Gonzalez-Delvalle, M.; Krumbein, W.E.; Laiz, L.; Petersen, K.; Saiz-Jimenez, C.; Swings, J. Bacterial and fungal diversity and biodeterioration problems in mural painting environments of St. Martins church (Greene-Kreiensen, Germany). Int. Biodeter. Biodegr. 2004, 53, 13–24. [Google Scholar] [CrossRef]
- Tsang, C.-C.; Chan, J.F.W.; Pong, W.-M.; Chen, J.H.K.; Ngan, A.H.Y.; Cheung, M.; Lai, C.K.C.; Tsang, D.N.C.; Lau, S.K.P.; Woo, P.C.Y. Cutaneous hyalohyphomycosis due to Parengyodontium album gen. et comb. nov. Med. Mycol. 2016, 54, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Jimenez, C.; Samson, R.A. Microorganisms and environmental pollution as deteriorating agents of the frescoes of “Santa María de la Rábida,” Huelva, Spain. In Proceedings of the 6th Triennial Meeting ICOM Committee for Conservation, Ottawa, Canada, 21–25 September 1981; p. 81/15/5. [Google Scholar]
- Karpovich-Tate, N.; Rebrikova, N.L. Microbial communities on damaged frescoes and building materials in the cathedral of the Nativity of the Virgin in the Pafnutii-Borovskii monastery, Russia. Int. Biodeter. 1990, 27, 281–296. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Petersen, K. Distribution of microorganisms on ancient wall paintings as related to associated faunal elements. Int. Biodeter. Biodegr. 2000, 46, 277–284. [Google Scholar] [CrossRef]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeter. Biodegr. 2008, 62, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, H.; Du, Y.; Tian, T.; Xiang, T.; Liu, X.; Wu, F.; An, L.; Wang, W.; Gu, J.-D.; et al. The community distribution of bacteria and fungi on ancient wall paintings of the Mogao Grottoes. Sci. Rep. 2015, 5, 7752. [Google Scholar] [CrossRef]
- Ma, W.; Wu, F.; Tian, T.; He, D.; Zhang, Q.; Gu, J.-D.; Duan, Y.; Ma, D.; Wang, W.; Feng, H. Fungal diversity and its contribution to the biodeterioration of mural paintings in two 1700-year-old tombs of China. Int. Biodeter. Biodegr. 2020, 152, 104972. [Google Scholar] [CrossRef]
- Duan, Y.; Wu, F.; Wang, W.; Gu, J.-D.; Li, Y.; Feng, H.; Chen, T.; Liu, G.; An, L. Differences of microbial community on the wall paintings preserved in situ and ex situ of the Tiantishan Grottoes, China. Int. Biodeter. Biodegr. 2018, 132, 102–113. [Google Scholar] [CrossRef]
- He, D.; Wu, F.; Ma, W.; Zhang, Y.; Gu, J.-D.; Duan, Y.; Xu, R.; Feng, H.; Wang, W.; Li, S.-W. Insights into the bacterial and fungal communities and microbiome that causes a microbe outbreak on ancient wall paintings in the Maijishan Grottoes. Int. Biodeter. Biodegr. 2021, 163, 105250. [Google Scholar] [CrossRef]
- Juretschko, S.; Timmermann, G.; Schmid, M.; Schleifer, K.-H.; Pommering-Röser, A.; Koops, H.-P.; Wagner, M. Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge: Nitrosococcus mobilis and Nitrospira-like bacteria as dominant populations. Appl. Environ. Microbiol. 1998, 64, 3042–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, U.; Rogall, T.; Bliicker, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 19, 7843–7853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR, Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, M.; Alisi, C.; Tasso, F.; Cecchini, A.; Marconi, P.; Sprocati, A.R. Laponite micro-packs for the selective celaning of multiple coherent deposits on wall paintings: The case study of Casina Farnese on the Palatine Hill (Rome-Italy). Int. Biodeter. Biodegr. 2014, 94, 1–11. [Google Scholar] [CrossRef]
- Lanyi, B. Classical and rapid identification methods for medically important bacteria. In Methods in Microbiology; Colwell, R.R., Grigorova, R., Eds.; Academic Press: London, UK, 1987; pp. 1–67. [Google Scholar]
- Unković, N.; Dimkić, I.; Supar, M.S.; Stanković, S.; Vukojević, J.; Grbić, M.L. Biodegradative potential of fungal isolates from sacral ambient: In vitro study as risk assessment implication for the conservation of wall paintings. PLoS ONE 2018, 13, e0190922. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, C.; Bakshi, U.; Mallick, I.; Mukherji, S.; Bera, B.; Ghosh, A. Genome-guided insights into the plant growth promotion capabilities of the physiologically versatile Bacillus aryabhattai strain AB211. Front. Microbiol. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Guan, Y.; Dong, Y.; Zhao, L.; Rong, S.; Chen, W.; Lv, M.; Xu, H.; Gao, X.; Chen, R.; et al. Isolation and evaluation of endophytic Bacillus tequilensis GYLH001 with potential application for biological control of Magnaporthe oryzae. PLoS ONE 2018, 13, e0203505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.H.; Liu, B.; Wang, J.P.; Che, J.M.; Li, P.F. Reclassification of Brevibacterium frigoritolerans DSM 8801T as Bacillus frigoritolerans comb. nov. based on genome analysis. Curr. Microbiol. 2020, 77, 1916–1923. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Kwon, S.-W.; Rooney, A.P.; Kim, S.-J. Bacillus paralicheniformis sp. nov., isolated from fermented soybean paste. Int. J. Syst. Evol. Microbiol. 2015, 65, 3487–3492. [Google Scholar] [CrossRef] [PubMed]
- Reva, O.N.; Smirnov, V.V.; Pettersson, B.; Priest, F.G. Bacillus endophyticus sp. nov., isolated from the inner tissues of cotton plants (Gossypium sp.). Int. J. Syst. Evol. Microbiol. 2002, 52, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Loqman, S.; Bouizgarne, B.; Barka, E.A.; Clément, C.; von Jan, M.; Spröer, C.; Klenk, H.-P.; Ouhdouch, Y. Streptomyces thinghirensis sp. nov., isolated from rhizosphere soil of Vitis vinifera. Int. J. Syst. Evol. Microbiol. 2009, 59, 3063–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepe, O.; Sannino, L.; Palomba, S.; Anastasio, M.; Blaiotta, G.; Villani, F.; Moschetti, G. Heterotrophic microorganisms in deteriorated medieval wall paintings in southern Italian churches. Microbiol. Res. 2010, 165, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, J.; Kiyuna, T.; Nishijima, M.; An, K.-D.; Nagatsuka, Y.; Tazato, N.; Handa, Y.; Hata-Yomita, J.; Sato, Y.; Kigawa, R.; et al. Polyphasic insights into the microbiomes of the Takamatsuzuka Tumulus and Kitora Tumulus. J. Gen. Appl. Microbiol. 2017, 63, 63–113. [Google Scholar] [CrossRef] [Green Version]
- Tomassetti, M.C.; Cirigliano, A.; Arrighi, C.; Negri, R.; Mura, F.; Maneschi, M.L.; Gentili, M.D.; Stipe, M.; Mazzoni, C.; Rinaldi, T. A role for microbial selection in frescoes’ deterioration in Tomba degli Scudi in Tarquinia, Italy. Sci. Rep. 2017, 7, 6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhagrassy, A.F. Isolation and characterization of actinomycetes from mural paintings of Snu-Sert-Ankh tomb, their antimicrobial activity, and their biodeterioration. Microbiol. Res. 2018, 216, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Caneva, G.; Isola, D.; Lee, H.J.; Chung, Y.J. Biological risk for hypogea: Shared data from Etruscan tombs in Italy and ancient tombs of the Baekje dynasty in Republic of Korea. Appl. Sci. 2020, 10, 6104. [Google Scholar] [CrossRef]
- Laiz, L.; Hermosin, B.; Caballero, B.; Saiz-Jimenez, C. Facultatively oligotrophic bacteria in Roman mural paintings. In Protection and Conservation of the Cultural Heritage of the Mediterranean Cities; Galan, E., Zezza, F., Eds.; Balkema: Lisse, The Netherlands, 2002; pp. 173–178. [Google Scholar]
- Dominguez-Moñino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne bacteria in show caves from Southern Spain. Microb. Cell 2021, in press. [Google Scholar]
- Srinath, B.S.; Namratha, K.; Byrappa, K. Green synthesis of biocompatible gold nanoparticles from gold mine bacteria Bacillus oceanisediminis and their antileukemic activity. Int. J. Pharm. Biol. Sci. 2018, 8, 169–177. [Google Scholar]
- Nugari, M.P.; Realini, M.; Roccardi, A. Contamination of mural paintings by indoor airborne fungal spores. Aerobiologia 1993, 9, 131–139. [Google Scholar] [CrossRef]
- An, K.-D.; Kiyuna, T.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. The identity of Penicillium sp. 1, a major contaminant of the stone chambers in the Takamatsuzuka and Kitora Tumuli in Japan, is Penicillium paneum. Anton. Leeuw. 2009, 96, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Ciferri, O. Microbial degradation of paintings. Appl. Environ. Microbiol. 1999, 65, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.A.; Shin, H.-D.; Dugan, F.M.; Schroers, H.-J.; Braun, U.; et al. Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef] [PubMed]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjevic, Z.; Andersen, B.; Houbraken, J.; Crous, P.W.; Samson, R.A. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef]
- Vaghefi, N.; Pethybridge, S.J.; Ford, R.; Nicolas, M.E.; Crous, P.W.; Taylor, P.W.J. Stagonosporopsis spp. associated with ray blight disease of Asteraceae. Australas. Plant Pathol. 2012, 41, 675–686. [Google Scholar] [CrossRef]
- Ropars, J.; Didiot, E.; Rodríguez de la Vega, R.C.; Bennetot, B.; Coton, M.; Poirier, E.; Coton, E.; Snirc, A.; Le Prieur, S.; Giraud, T. Domestication of the emblematic white cheese-making fungus Penicillium camemberti and its diversification into two varieties. Curr. Biol. 2020, 30, 4441–4453. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Michailides, T.J.; Xiao, C.L. Mucor rot–An emerging postharvest disease of mandarin fuit caused by Mucor piriformis and other Mucor spp. in California. Plant Dis. 2016, 100, 1054–1063. [Google Scholar] [CrossRef] [Green Version]
- Schultes, N.P.; Strzalkowski, N.; Li, D.-W. Botryotrichum domesticum sp. nov., a new hyphomycete from an indoor environment. Botany 2019, 87, 311–319. [Google Scholar] [CrossRef]
- Pangallo, D.; Kraková, L.; Chovanová, K.; Šimonovicová, A.; De Leo, F.; Urzì, C. Analysis and comparison of the microflora isolated from fresco surface and from surrounding air environment through molecular and biodegradative assays. World J. Microbiol. Biotechnol. 2012, 28, 2015–2027. [Google Scholar] [CrossRef] [PubMed]
- Pangallo, D.P.; Chovanová, K.C.; Šimonovicová, A.Š.; Ferianc, P.F. Investigation of microbial community isolated from indoor artworks and air environment: Identification, biodegradative abilities, and DNA typing. Can. J. Microbiol. 2009, 55, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Savković, Ž.; Stupar, M.; Unković, N.; Ivanović, Ž.; Blagojević, J.; Vukojević, J.; Grbić, M.L. In vitro biodegradation potential of airborne Aspergilli and Penicillia. Sci. Nat. 2019, 106, 8. [Google Scholar] [CrossRef] [PubMed]
- Trovão, J.; Tiago, I.; Catarino, L.; Gil, F.; Portugal, A. In vitro analyses of fungi and dolomitic limestone interactions: Bioreceptivity and biodeterioration assessment. Int. Biodeter. Biodegr. 2020, 155, 105107. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-pigmented biodeteriogenic fungi in etruscan hypogeal tombs: New data on their culture-dependent diversity, favouring conditions, and resistance to biocidal treatments. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, M.; Pietrini, A.M. La disinfezione delle patine biologiche sui manufatti lapidei: Biocidi chimici e naturali a confronto. Boll. ICR 2016, 33, 40–49. [Google Scholar]
- Vannini, A.; Contardo, T.; Paoli, L.; Scattoni, M.; Favero-Longo, S.E.; Loppi, S. Application of commercial biocides to lichens: Does a physiological recovery occur over time? Int. Biodeter. Biodegr. 2018, 129, 189–194. [Google Scholar] [CrossRef]
- Lo Schiavo, S.; De Leo, F.; Urzì, C. Present and future perspectives for biocides and antifouling products for stone-built cultural heritage: Ionic liquids as a challenging alternative. Appl. Sci. 2020, 10, 6568. [Google Scholar] [CrossRef]
- Sanmartín, P.; Rodríguez, A.; Aguiar, U. Medium-term field evaluation of several widely used cleaning-restoration techniques applied to algal biofilm formed on a granite-built historical monument. Int. Biodeter. Biodegr. 2020, 147, 104870. [Google Scholar] [CrossRef]
- Sanmartín, P.; Carballeira, R. Changes in heterotrophic microbial communities induced by biocidal treatments in the Monastery of San Martiño Pinario (Santiago de Compostela, NW Spain). Int. Biodeter. Biodegr. 2021, 156, 105130. [Google Scholar] [CrossRef]
- Coutinho, M.L.; Miller, A.Z.; Martin-Sanchez, P.M.; Mirao, J.; Gomez-Bolea, A.; Machado-Moreira, B.; Cerqueira-Alves, L.; Jurado, V.; Saiz-Jimenez, C.; Lima, A.; et al. A multiproxy approach to evaluate biocidal treatments on biodeteriorated majolica glaze tiles. Environ. Microbiol. 2016, 18, 4794–4816. [Google Scholar] [CrossRef] [PubMed]
- Romani, M.; Warscheid, T.; Nicole, L.; Marcon, L.; Di Martino, P.; Suzuki, M.T.; Lebaron, P.; Lami, R. Current and future chemical treatments to fight biodeterioration of outdoor building materials and associated biofilms: Moving away from ecotoxic and towards efficient, sustainable solutions. Sci. Total. Environ. 2021, 802, 149846. [Google Scholar] [CrossRef]
- Presentato, A.; Armetta, F.; Spinella, A.; Chillura Martino, D.F.; Alduina, R.; Saladino, M.L. Formulation of mesoporous silica nanoparticles for controlled release of antimicrobials for stone preventive conservation. Front. Chem. 2020, 8, 699. [Google Scholar] [CrossRef]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Impact of biocide treatments on the bacterial communities of the Lascaux Cave. Naturwissenschaften 2009, 96, 863–868. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Camaiti, M.; Borgioli, L.; Rosi, L. Photostability of innovative formulations for artworks restoration. Chim. Ind. 2011, 9, 100–195. [Google Scholar]
- Gheno, G.; Badetti, E.; Brunelli, A.; Ganzerla, R.; Marcomini, A. Consolidation of Vicenza, Arenaria and Istria stones: A comparison between nano-based products and acrylate derivatives. J. Cult. Herit. 2018, 32, 44–52. [Google Scholar] [CrossRef]
- Negri, A.; Nervo, M.; Di Marcello, S.; Castelli, D. Consolidation and adhesion of pictorial layers on a stone substrate. The study case of the Virgin with the Child from Palazzo Madama, in Turin. Coatings 2021, 11, 624. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
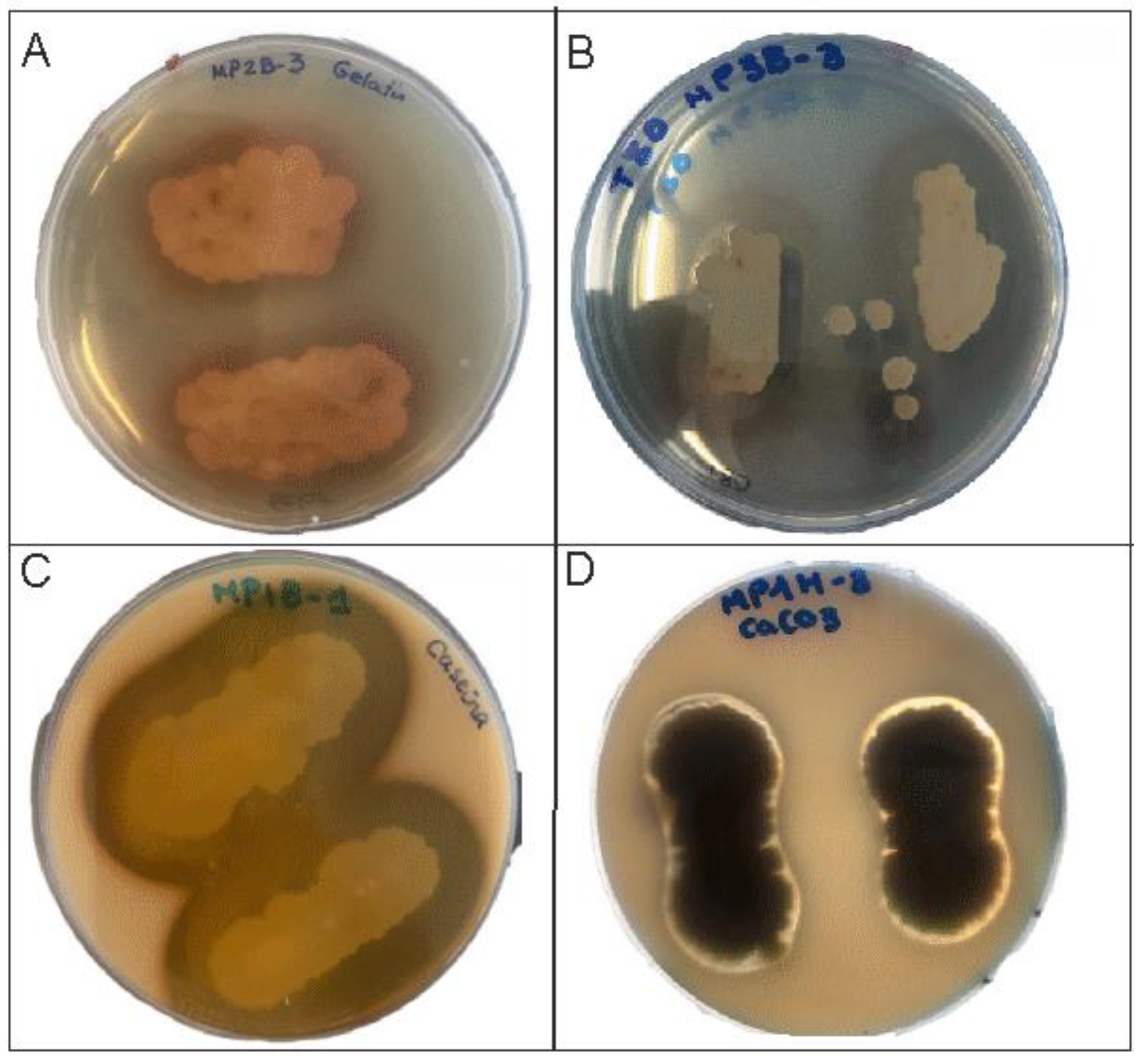{kind=link}
| Sample | Description | Location |
|---|---|---|
| M1 | Surface dust | S-3. Mexico shield |
| M2 | Black stains | S-4. Burgundy shield |
| M3 | Black stains | S-02. Between Galicia and Murcia shields |
| Medium | Composition * | Analysis | Reference |
|---|---|---|---|
| GH | TSA/MEA + 1% gelatin | Proteolytic activity | [24] |
| LH | 1% Tween 80 | Lipolytic activity | [25] |
| CH | Nutrient Casein Agar | Proteolytic activity | [26] |
| CaS | TSA/MEA + 1% CaCO3 | Calcite solubilization | [24] |
| GyS | TSA/MEA + 1% CaSO4 | Gypsum solubilization | [24] |
| AAP | Czapek-Dox broth | Acid/Alkali production | [26] |
| Samples | Accession Number | Strains | Identification and Accession Number (% Identity) |
|---|---|---|---|
| M1 | MZ827450 | MP1B-1 | Priestia aryabhattai EF114313 (99.7%) |
| MZ827451 | MP1B-2 | Bacillus tequilensis AYTO01000043 (99.9%) | |
| MZ827452 | MP1B-3 | Bacillus paralicheniformis KY694465 (99.7%) | |
| M2 | MZ827453 | MP2B-2 | Bacillus frigoritolerans AM747813 (99.7%) |
| MZ827454 | MP2B-3 | Bacillus paralicheniformis KY694465 (99.7%) | |
| MZ827455 | MP2B-4 | Bacillus cereus AE016877 (99.6%) | |
| M3 | MZ827456 | MP3B-1 | Cytobacillus oceanisediminis GQ292772 (99.4%) |
| MZ827457 | MP3B-2 | Streptomyces thinghirensis FM202482 (99.6%) | |
| MZ827458 | MP3B-3 | Priestia endophytica AF295302 (99.4%) | |
| MZ827459 | MP3B-6 | Bacillus frigoritolerans AM747813 (99.7%) |
| Samples | Accession Number | Strains | Identification and Accession Number (% Identity) |
|---|---|---|---|
| M1 | MZ827828 | MP1B-4 | Penicillium chrysogenum MF422150 (100%) |
| MZ827829 | MP1B-5 | Penicillium chrysogenum MF422150 (99.7%) | |
| MZ827830 | MP1H-1 | Alternaria angustiovoidea MK910070 (100%) | |
| MZ827831 | MP1H-2 | Cladosporium xylophilum MH863875 (100%) | |
| MZ827832 | MP1H-3 | Penicillium fuscoglaucum NR163669 (100%) | |
| MZ827833 | MP1H-4 | Mucor racemosus NR126135 (99.0%) | |
| MZ827834 | MP1H-5 | Cladosporium macrocarpum NR119657 (100%) | |
| MZ827835 | MP1H-6 | Penicillium chrysogenum MF422150 (100%) | |
| M2 | MZ827836 | MP2H-1 | Stagonosporopsis lupini NR160205 (98.3%) |
| M3 | MZ827837 | MP3B-5 | Penicillium chrysogenum MF422150 (99.7%) |
| MZ827838 | MP3H-1 | Mucor racemosus NR126135 (99.0%) | |
| MZ827839 | MP3H-2 | Penicillium chrysogenum MF422150 (100%) | |
| MZ827840 | MP3H-3 | Penicillium chrysogenum MF422150 (100%) | |
| MZ827841 | MP3H-5 | Botryotrichum domesticum NR169944(98.6%) |
| Strain Reference | GH * | LH | CH | GyS | CaS | AAP |
|---|---|---|---|---|---|---|
| Priestia aryabhattai, strain MP1B-1 | - | + | + | - | - | + |
| Bacillus tequilensis, strain MP1B-2 | - | + | - | + | - | - |
| Bacillus paralicheniformis, strain MP1B-3 | + | - | - | - | - | - |
| Penicillium chrysogenum, strain MP1B-4 | - | - | - | - | - | - |
| Penicillium chrysogenum, strain MP1B-5 | - | + | + | - | - | - |
| Alternaria angustiovoidea, strain MP1H-1 | - | - | - | - | - | - |
| Cladosporium xylophilum, strain MP1H-2 | - | + | - | - | - | - |
| Penicillium fuscoglaucum, strain MP1H-3 | - | + | + | - | + | - |
| Mucor racemosus, strain MP1H-4 | - | - | - | - | - | - |
| Cladosporium macrocarpum, strain MP1H-5 | - | + | - | - | - | - |
| Penicillium chrysogenum, strain MP1H-6 | - | - | + | - | + | - |
| Bacillus frigoritolerans, strain MP2B-2 | - | + | + | + | - | + |
| Bacillus paralicheniformis, strain MP2B-3 | + | - | - | - | - | - |
| Bacillus cereus, strain MP2B-4 | - | - | - | - | - | - |
| Stagonosporopsis lupini, strain MP2H-1 | - | - | + | - | - | - |
| Cytobacillus oceanisediminis, strain MP3B-1 | - | - | + | - | - | - |
| Streptomyces thinghirensis, strain MP3B-2 | - | + | + | - | - | - |
| Priestia endophytica, strain MP3B-3 | - | + | + | + | - | + |
| Bacillus frigoritolerans, strain MP3B-6 | - | + | + | - | - | + |
| Penicillium chrysogenum, strain MP3B-5 | - | - | + | - | - | - |
| Mucor racemosus, strain MP3H-1 | - | - | - | - | - | - |
| Penicillium chrysogenum, strain MP3H-2 | - | + | + | - | - | - |
| Penicillium chrysogenum, strain MP3H-3 | - | - | + | - | - | - |
| Botryotrichum domesticum, strain MP3H-5 | - | + | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. https://doi.org/10.3390/app11198858
Jurado V, Gonzalez-Pimentel JL, Hermosin B, Saiz-Jimenez C. Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Applied Sciences. 2021; 11(19):8858. https://doi.org/10.3390/app11198858
Chicago/Turabian StyleJurado, Valme, José Luis Gonzalez-Pimentel, Bernardo Hermosin, and Cesareo Saiz-Jimenez. 2021. "Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain" Applied Sciences 11, no. 19: 8858. https://doi.org/10.3390/app11198858
APA StyleJurado, V., Gonzalez-Pimentel, J. L., Hermosin, B., & Saiz-Jimenez, C. (2021). Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Applied Sciences, 11(19), 8858. https://doi.org/10.3390/app11198858