
Impacts of Diverse Natural Products on Honey Bee Viral Loads and Health

 , ,
, , 
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. RNA Extraction, cDNA Synthesis, and qPCR
2.3. Data Analysis
3. Results
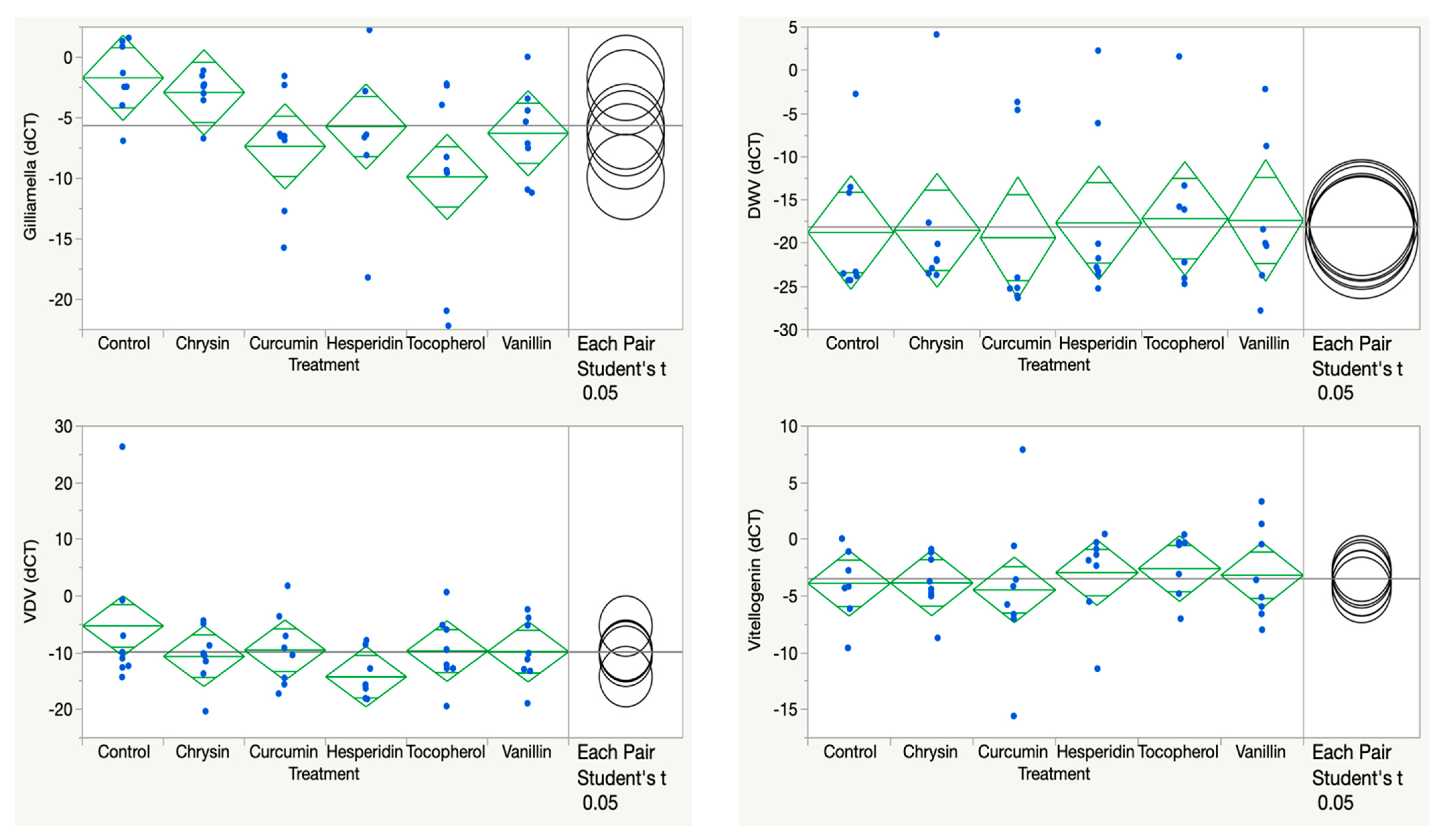
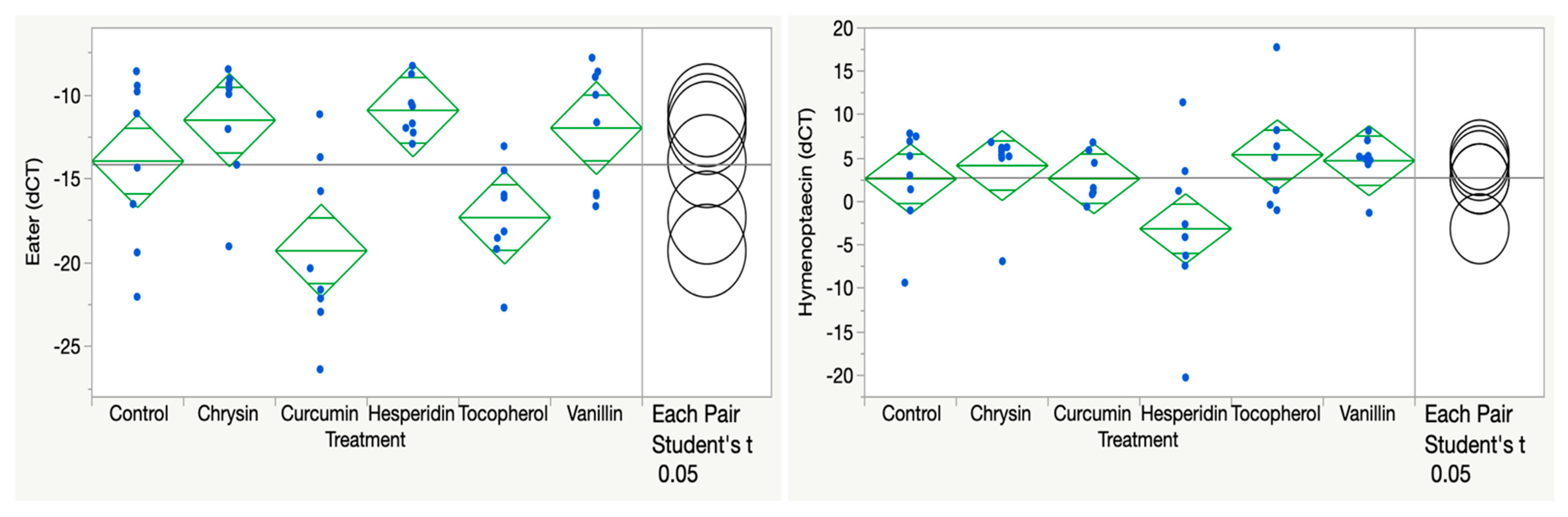
3.1. Chrysin, Curcumin, Hesperidin, Vanillin, and Tocopherol
3.2. Raw Cacao, Roasted Cacao, Limonene, and Tyrosine
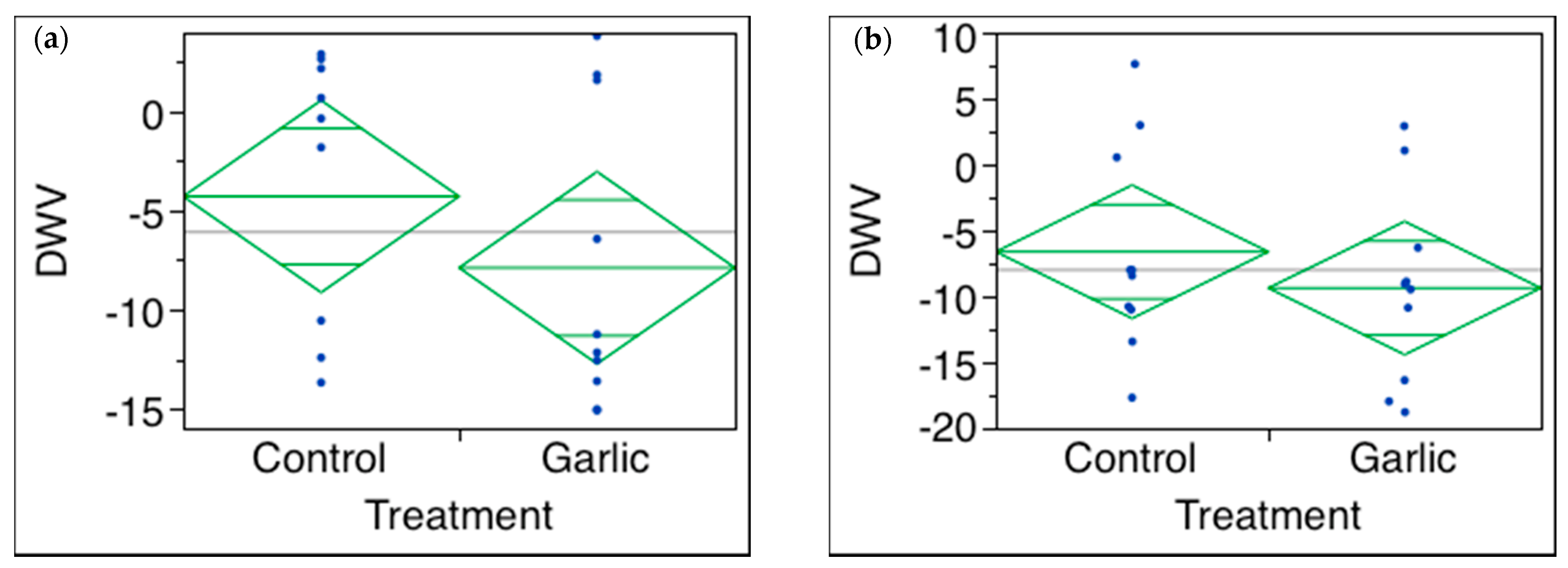
3.3. Cat’s Claw, Garlic Oil, Olive Leaf, Oregano Oil, and Elderberry
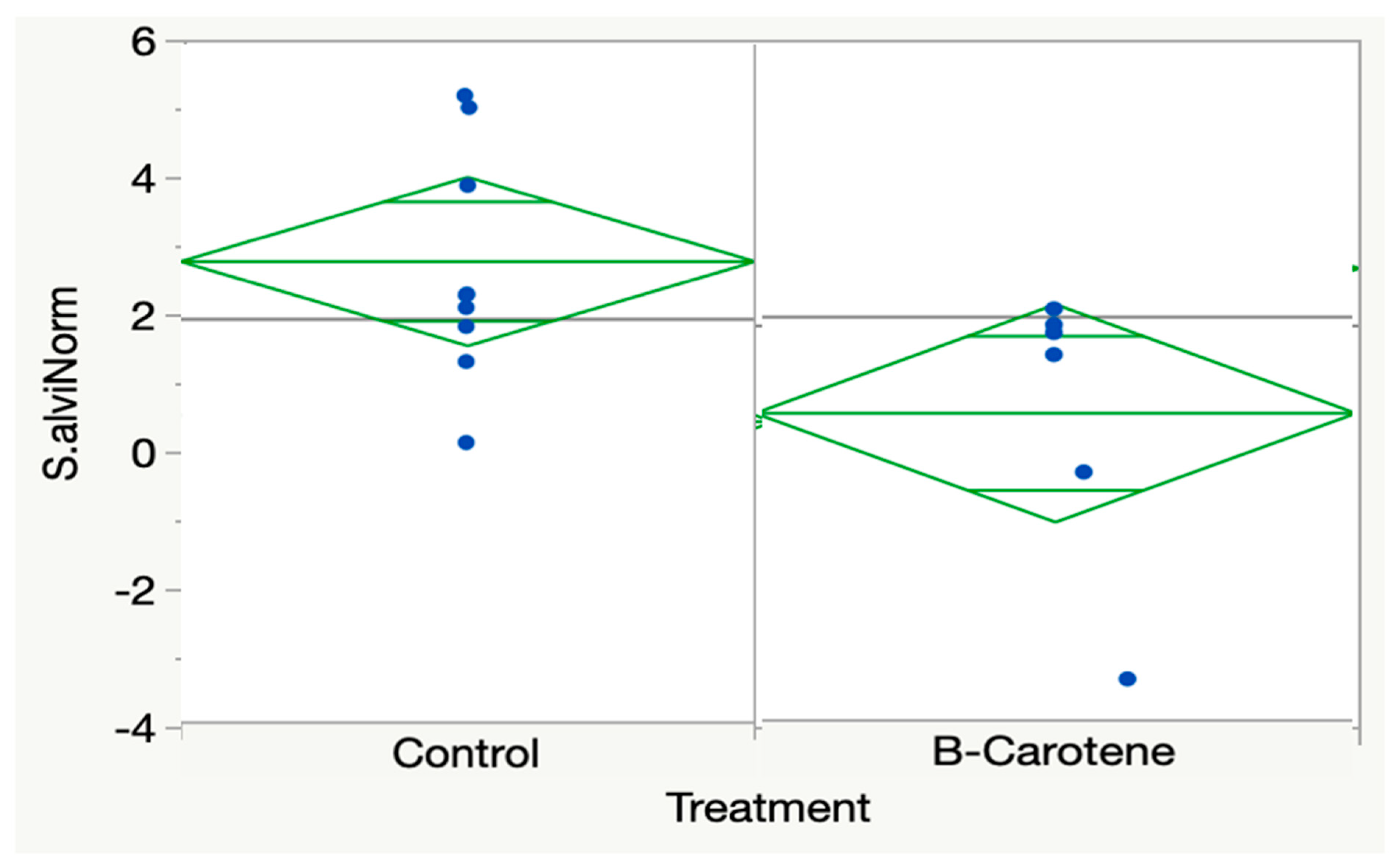
3.4. Beta-Carotene
3.5. Carvacrol, Cinnamic Acid, and Ginger
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jordan, A.; Patch, H.M.; Grozinger, C.M.; Khanna, V. Economic Dependence and Vulnerability of United States Agricultural Sector on Insect-Mediated Pollination Service. Environ. Sci. Technol. 2021, 55, 2243–2253. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Huang, Q.; Evans, J.D. Hologenome theory and the honey bee pathosphere. Curr. Opin. Insect Sci. 2015, 10, 1–7. [Google Scholar] [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.P.; vanEngelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Noël, A.; Le Conte, Y.; Mondet, F. Varroa destructor: How does it harm Apis mellifera honey bees and what can be done about it? Emerg. Top. Life Sci. 2020, 4, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the Major Ectoparasite of the Western Honey Bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Highfield, A.C.; El Nagar, A.; Mackinder, L.C.M.; Noël, L.M.L.J.; Hall, M.J.; Martin, S.J.; Schroeder, D.C. Deformed wing virus implicated in overwintering honeybee colony losses. Appl. Environ. Microbiol. 2009, 75, 7212–7220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barroso-Arévalo, S.; Fernández-Carrión, E.; Goyache, J.; Molero, F.; Puerta, F.; Sánchez-Vizcaíno, J.M. High Load of Deformed Wing Virus and Varroa destructor Infestation Are Related to Weakness of Honey Bee Colonies in Southern Spain. Front. Microbiol. 2019, 10, 1331. [Google Scholar] [CrossRef] [PubMed]
- Benaets, K.; Van Geystelen, A.; Cardoen, D.; De Smet, L.; De Graaf, D.C.; Schoofs, L.; Larmuseau, M.H.D.; Brettell, L.E.; Martin, S.J.; Wenseleers, T. Covert deformed wing virus infections have long-term deleterious effects on honeybee foraging and survival. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162149. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Grozinger, C.M.; Flenniken, M.L. Bee viruses: Ecology, pathogenicity, and impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Chen, Y.P.; Ribière, M.; Gauthier, L. Varroa and Viruses. In Varroa—Still a Problem in the 21st Century? Carreck, N.L., Ed.; International Bee Research Association: Cardiff, UK, 2011; pp. 11–31. [Google Scholar]
- Boncristiani, H.; Ellis, J.D.; Bustamante, T.; Graham, J.; Jack, C.; Kimmel, C.B.; Mortensen, A.; Schmehl, D.R. World Honey Bee Health: The Global Distribution of Western Honey Bee (Apis mellifera L.) Pests and Pathogens. Bee World 2021, 98, 2–6. [Google Scholar] [CrossRef]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumanna, P. Dead or alive: Deformed wing virus and varroa destructor reduce the life span of winter honeybees. Appl. Environ. Microbiol. 2012, 78, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryabov, E.V.; Childers, A.K.; Lopez, D.; Grubbs, K.; Posada-Florez, F.; Weaver, D.; Girten, W.; vanEngelsdorp, D.; Chen, Y.; Evans, J.D. Dynamic evolution in the key honey bee pathogen deformed wing virus: Novel insights into virulence and competition using reverse genetics. PLoS Biol. 2019, 17, e3000502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauber, J.P.; Collins, W.R.; Schwarz, R.S.; Chen, Y.; Grubbs, K.; Huang, Q.; Lopez, D.; Peterson, R.; Evans, J.D. Natural Product Medicines for Honey Bees: Perspective and Protocols. Insects 2019, 10, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honeybee products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [Green Version]
- Mani, J.S.; Johnson, J.B.; Steel, J.C.; Broszczak, D.A.; Neilsen, P.M.; Walsh, K.B.; Naiker, M. Natural product-derived phytochemicals as potential agents against coronaviruses: A review. Virus Res. 2020, 284, 197989. [Google Scholar] [CrossRef] [PubMed]
- Byler, K.G.; Ogungbe, I.V.; Setzer, W.N. In-silico screening for anti-Zika virus phytochemicals. J. Mol. Graph. Model. 2016, 69, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.C.; Kim, Y.W.; Ryu, S.; Lee, A.; Lee, J.S.; Song, M.J. Suppression of norovirus by natural phytochemicals from Aloe vera and Eriobotryae Folium. Food Control 2017, 73, 1362–1370. [Google Scholar] [CrossRef]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Vijayasri, S.; Hopper, W. Towards the identification of novel phytochemical leads as macrodomain inhibitors of Chikungunya virus using molecular docking approach. J. Appl. Pharm. Sci. 2017, 7, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Kamei, M.; Nishimura, H.; Takahashi, T.; Takahashi, N.; Inokuchi, K.; Mato, T.; Takahashi, K. Anti-influenza virus effects of cocoa. J. Sci. Food Agric. 2016, 96, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef] [Green Version]
- Simone-Finstrom, M.D.; Spivak, M. Increased resin collection after parasite challenge: A case of self-medication in honey bees? PLoS ONE 2012, 7, e34601. [Google Scholar]
- Simone-Finstrom, M.; Borba, R.S.; Wilson, M.; Spivak, M. Propolis Counteracts Some Threats to Honey Bee Health. Insects 2017, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erler, S.; Moritz, R.F.A. Pharmacophagy and pharmacophory: Mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie 2016, 47, 389–411. [Google Scholar] [CrossRef] [Green Version]
- Stamets, P.E.; Naeger, N.L.; Evans, J.D.; Han, J.O.; Hopkins, B.K.; Lopez, D.; Moershel, H.M.; Nally, R.; Sumerlin, D.; Taylor, A.W.; et al. Extracts of Polypore Mushroom Mycelia Reduce Viruses in Honey Bees. Sci. Rep. 2018, 8, 13936. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Young, E.C.; Tozkar, C.O.; Schwarz, R.S.; Chen, Y.; Irwin, R.E.; Adler, L.S.; Evans, J.D. Nectar and pollen phytochemicals stimulate honey bee (Hymenoptera: Apidae) immunity to viral infection. J. Econ. Entomol. 2017, 110, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its’ Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Mikail, P.; Maadirad, S.; Moloudizargari, M.; Aghajanshakeri, S.; Sarahroodi, S. Therapeutic Uses and Pharmacological Properties of Garlic, Shallot, and Their Biologically Active Compounds. Iran J. Basic Med. Sci. 2013, 16, 1031–1048. [Google Scholar]
- Nakamoto, M.; Kunimura, K.; Suzuki, J.I.; Kodera, Y. Antimicrobial properties of hydrophobic compounds in garlic: Allicin, vinyldithiin, ajoene and diallyl polysulfides (Review). Exp. Ther. Med. 2020, 19, 1550–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokić, S.A.-O.; Gagić, T.A.-O.; Knez, Ž.; Šubarić, D.; Škerget, M. Separation of Active Compounds from Food by-Product (Cocoa Shell) Using Subcritical Water Extraction. Molecules 2018, 23, 1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillier, N.K.; Frost, E.H.; Shutler, D. Fate of dermally applied miticides fluvalinate and amitraz within honey bee (hymenoptera: Apidae) bodies. J. Econ. Entomol. 2013, 106, 558–565. [Google Scholar] [CrossRef]
- Turcatto, A.P.; Lourenço, A.P.; De Jong, D. Propolis consumption ramps up the immune response in honey bees infected with bacteria. Apidologie 2018, 49, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Naithani, R.; Huma, L.C.; Holland, L.E.; Shukla, D.; McCormick, D.L.; Mehta, R.G.; Moriarty, R.M. Antiviral activity of phytochemicals: A comprehensive review. Mini-Rev. Med. Chem. 2008, 8, 1106–1133. [Google Scholar] [CrossRef] [PubMed]
- Bernklau, E.; Bjostad, L.; Hogeboom, A.; Carlisle, A.; Arathi, H.S. Dietary phytochemicals, honey bee longevity and pathogen tolerance. Insects 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Geldert, C.; Abdo, Z.; Stewart, J.E.; Arathi, H.S. Dietary supplementation with phytochemicals improves diversity and abundance of honey bee gut microbiota. J. Appl. Microbiol. 2021, 130, 1705–1720. [Google Scholar] [CrossRef]
- Palmer-Young, E.C.; Sadd, B.M.; Irwin, R.E.; Adler, L.S. Synergistic effects of floral phytochemicals against a bumble bee parasite. Ecol. Evol. 2017, 7, 1836–1849. [Google Scholar] [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LoCascio, G.M.; Aguirre, L.; Irwin, R.E.; Adler, L.S. Pollen from multiple sunflower cultivars and species reduces a common bumblebee gut pathogen. R. Soc. Open Sci. 2019, 6, 190279. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Chen, Y.P.; Di Prisco, G.; Pettis, J.; Williams, V. Bee cups: Single-use cages for honey bee experiments. J. Apic. Res. 2009, 48, 300–302. [Google Scholar] [CrossRef]
- Evans, J.D.; Schwarz, R.S.; Chen, Y.P.; Budge, G.; Cornman, R.S.; De La Rua, P.; De Miranda, J.R.; Foret, S.; Foster, L.; Gauthier, L.; et al. Standard methods for molecular research in Apis mellifera. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Nwokeoji, A.O.; Kilby, P.M.; Portwood, D.E.; Dickman, M.J. RNASwift: A rapid, versatile RNA extraction method free from phenol and chloroform. Anal Biochem. 2016, 512, 36–46. [Google Scholar] [CrossRef] [PubMed]
- De la Luz Cádiz-Gurrea, M.; Fernández de las Nieves, I.; Aguilera Saez, L.M.; Fernández-Arroyo, S.; Legeai-Mallet, L.; Bouaziz, M.; Segura-Carretero, A. Bioactive Compounds from Theobroma cacao: Effect of Isolation and Safety Evaluation. Plant Foods Hum. Nutr. 2019, 74, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.K.T.; Nguyen, H.A.-O. Effects of Maturity at Harvest and Fermentation Conditions on Bioactive Compounds of Cocoa Beans. Plant Foods Hum. Nutr. 2019, 74, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Kim, B.Y.; Baik, M.Y. Physicochemical properties and antioxidant capacity of raw, roasted and puffed cacao beans. Food Chem. 2016, 194, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Salvador, I.; Massarioli, A.P.; Silva, A.P.; Malaguetta, H.; Melo, P.S.; Alencar, S.A.-O. Can we conserve trans-resveratrol content and antioxidant activity during industrial production of chocolate? J. Sci. Food Agric. 2019, 99, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Haghmorad, D.; Mahmoudi, M.B.; Salehipour, Z.; Jalayer, Z.; Momtazi Brojeni, A.A.; Rastin, M.; Kokhaei, P.; Mahmoudi, M. Hesperidin ameliorates immunological outcome and reduces neuroinflammation in the mouse model of multiple sclerosis. J. Neuroimmunol. 2017, 302, 23–33. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and human health: A comprehensive review. Phytother. Res. 2018, 32, 1675–1687. [Google Scholar] [CrossRef]
- Zingg, J.M. Vitamin E: An overview of major research directions. Mol. Asp. Med. 2007, 28, 400–422. [Google Scholar] [CrossRef]
- Kodad, O.; Socias, I.C.R.; Alonso, J.M. Genotypic and Environmental Effects on Tocopherol Content in Almond. Antioxidants 2018, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Rupasinghe, H.P.V.; Boulter-Bitzer, J.; Ahn, T.; Odumeru, J.A. Vanillin inhibits pathogenic and spoilage microorganisms in vitro and aerobic microbial growth in fresh-cut apples. Food Res. Int. 2006, 39, 575–580. [Google Scholar] [CrossRef]
- Zheng, H.; Nishida, A.; Kwong, W.K.; Koch, H.; Engel, P.; Steele, M.I.; Moran, N.A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. mBio 2016, 7, e01326-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gul, K.; Tak, A.; Singh, A.K.; Singh, P.; Yousuf, B.; Wani, A.A. Chemistry, encapsulation, and health benefits of β-carotene—A review. Cogent Food Agric. 2015, 1, 1018696. [Google Scholar] [CrossRef]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Predictive markers of honey bee colony collapse. PLoS ONE 2012, 7, e32151. [Google Scholar] [CrossRef] [PubMed]
- Javeri, I.; Chand, N. Chapter 31-Curcumin. In Nutraceuticals; Gupta, R.C., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 435–445. [Google Scholar]
- Nagpal, M.; Sood, S. Role of curcumin in systemic and oral health: An overview. J. Nat. Sci. Biol. Med. 2013, 4, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.H.; Loo, C.Y.; Bebawy, M.; Luk, F.; Mason, R.S.; Rohanizadeh, R. Curcumin and its derivatives: Their application in neuropharmacology and neuroscience in the 21st century. Curr. Neuropharmacol. 2013, 11, 338–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, Y. Physiological and ecological functions and biosynthesis of health-promoting compounds in fruit and vegetables. In Improving the Health-Promoting Properties of Fruit and Vegetable Products; Tomas Barberan, F.A., Gil, M.I., Eds.; Woodhead Publishing Ltd.: Cambridge, UK, 2008; pp. 201–247. [Google Scholar]
- Das, A.B.; Goud, V.V.; Das, C. 9-Phenolic Compounds as Functional Ingredients in Beverages. In Value-Added Ingredients and Enrichments of Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 285–323. [Google Scholar]
- Duke, J.A. Handbook of Phytochemical Constituents of Grass, Herbs and Other Economic Plants; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name (Scientific Name) | Level of Purity | Source | Conc. (ppm) |
|---|---|---|---|
| Astragalus (Astragalus propinquus) | Natural | Vitamin Shoppe#VS-1637/1776483 | 100, 1000 |
| Berberine chloride | 98% | Sigma#B3251 | 100 |
| Beta-carotene | 95% | Sigma#C4582 | 2000 |
| Cacao raw (Theobroma cacao L) | Natural | Whole raw cocoa beans (Peru) | 100 |
| Cacao roasted (Theobroma cacao L) | Natural | Whole roasted cocoa beans (Peru) | 100 |
| Calendula (Calendula officinalis) | Natural | Vitamin Shoppe # BB-1034/1083588 | 100, 1000 |
| Cat’s Claw (Uncaria tomentosa) | Natural | Vitamin Shoppe #BB-1346/1098914 | 100, 1000 |
| Carvacrol | 98% | Sigma#282197 | 100 |
| Chrysin | 97% | Sigma#C80105 | 100 |
| Cinnamic acid | 99% | Sigma#W228826 | 100 |
| Curcumin (Curcuma longa) | 75% | Sigma#8.20354 | 100 |
| Decanoic acid | 98% | Sigma #C1875 | 100 |
| Echinacea (Echinacea) | Natural | Vitamin Shoppe #VS-1625/1776038 | 100, 1000 |
| Elderberry (Sambucus) | Natural | Vitamin Shoppe #BB-1181/1681436 | 100, 1000 |
| Garlic oil (Allium sativum) | Natural | Vitamin Shoppe | 100, 1000, 10,000 |
| Ginger (Zingiber officinale) | Natural | Sigma#W252204 | 100, 1000 |
| Hesperidin | 80% | Sigma#H5254 | 100 |
| Licorice (Glycyrrhiza glabra) | Natural | Vitamin Shoppe #BB-1101/1087241 | 100, 1000 |
| Limonene | 97% | Sigma#183164 | 100 |
| Olive leaf (Olea europaea) | Natural | Vitamin Shoppe | 100, 1000 |
| Oil of oregano (Origanum vulgare) | Natural | Vitamin Shoppe #VS-1782/1484956 | 100, 1000 |
| Thymol | 99% | Sigma#16254 | 10, 100 |
| Tocopherol | FCG | Sigma#W530066 | 100 |
| Tyrosine low, high | 98% | Sigma#T3754 | 10, 100 |
| Vanillin (Vanilla planifolia) | 99% | Sigma#V1104 | 100 |
| Compound | Honey Bee Immune Gene | Virus/Microbe |
|---|---|---|
| Beta-carotene 2000 ppm | Abaecin NS (p = 0.3702) Hymenoptaecin NS (p = 0.8662) Apidaecin NS (p = 0.8347) PGRP-LC NS (p = 0.1352) Vitellogenin NS (p = 0.1391) Eater NS (p = 0.5425) MRJP1 NS (p = 0.7023) CytochromeP450 NS (p = 0.1735) Defensin1 NS (p = 0.8176) Endochitinase NS (p = 0.5428) Glucosidase NS (p = 0.6669) Trehalase NS (p = 0.2995) Peritrophin NS (p = 0.5006) | DWV NS (p = 0.6862) VDV NS (p = 0.6531) S. alvi lower (mean control = 2.6885 +- 0.5713 (SE) n = 10; mean beta-carotene = 0.4533 +− 0.7376 (SE), n = 6, (p = 0.0311) |
| Cacao (raw) 100 ppm | DWV increased (B control vs. A cacao raw, p = 0.0307) VDV decrease (A control vs. B cacao raw, p = 0.0013) | |
| Cacao (roasted) 100 ppm | DWV NS (B control vs. AB cacao roasted, p = 0.7970) VDV NS (A control vs. A cacao roasted, p = 0.9769) | |
| Carvacrol 100 ppm | Eater NS (p = 0.3931) Hymenoptaecin lower (A control vs. AB Carvacrol p = 0.1797) Vitellogenin NS (p = 0.9502) | DWV NS (p = 0.8588) |
| Cat’s Claw Jill2 1000 ppm | DWV NS (p = 0.5532) | |
| Chrysin 100 ppm | Eater lower (AB control vs. A Chrysin, p = 0.2149) Hymenoptaecin NS (p = 0.5896) Vitellogenin NS (p = 0.9854) | DWV NS (p = 0.9571) Trypanosome NS (p = 0.1276) VDV lower (A control vs. AB chrysin p = 0.1592) Gilliamella lower (A control vs. AB chrysin p = 0.6296) |
| Cinnamic acid 100 ppm | Eater NS, p = 0.8382 Hymenoptaecin lower (A control vs. B cinnamic acid, p = 0.0392) Vitellogenin NS (p = 0.1909) | DWV NS, p = 0.8911 |
| Curcumin 100 ppm | Eater lower (AB control vs. C curcumin, p = 0.0086) Hymenoptaecin NS (p = 0.9952) Vitellogenin NS (p = 0.7768) | DWV NS (p = 0.9001) Trypanosome NS (p = 0.1776) VDV lower (A control vs. AB curcumin p = 0.2573) Gilliamella lower (A control vs. BC curcumin p = 0.0268) |
| Elderberry Jill2 1000 ppm | DWV NS (p = 0.5940) | |
| Ginger 100 ppm | Eater NS p = 0.7334 Hymenoptaecin lower (A control vs. AB ginger p = 0.4487) Vitellogenin NS (p = 0.2233) | DWV NS (p = 0.7434) |
| Garlic oil 1000 (A&B) or 10,000 ppm (C) | DWV NS (p = 0.2780) (Supplemental File experiment A) DWV NS (p = 0.6484) (B) DWV NS (p = 0.4288) (C) | |
| Hesperidin 100 ppm | Eater lower (AB control vs. A hesperidin, p = 0.1260) Hymenoptaecin lower (A vs. B P = 0.0475) Vitellogenin NS (p = 0.6422) | DWV NS (p = 0.8099) Trypanosome NS (p = 0.3059) VDV lower (A control vs. B hesperidin p = 0.0208) Gilliamella lower (A control vs. ABC hesperidin, p = 0.1104) |
| Limonene 100 ppm | DWV NS (B control vs. AB limonene, p = 0.1844) VDV NS (A control vs. A limonene, p = 0.4293) | |
| Olive leaf 1000 ppm | DWV NS (p = 0.2295)-(B) | |
| Oregano oil 1000 ppm | DWV NS (p = 0.3750)-(A) DWV NS (p = 0.5759)-(B) | |
| Tocopherol 100 ppm | Eater lower (AB control vs. BC tocopherol, p = 0.0899) Hymenoptaecin NS (p = 0.3320) Vitellogenin NS (p = 0.5248) | DWV NS (p = 0.7300) Trypanosome NS (p = 0.4107) VDV lower (A control vs. AB tocopherol p = 0.2414) Gilliamella lower (A control vs. C tocopherol, p = 0.0019) |
| Tyrosine (high) 100 ppm | DWV NS (B control vs. AB tyrosine (high), p = 0.3345) VDV NS (A control vs. A tyrosine (high), p = 0.7490) | |
| Tyrosine (low) 10 ppm | DWV NS (B control vs. AB tyrosine (low), p = 0.4598) VDV NS (A control vs. tyrosine (low), p = 0.8813) | |
| Vanillin 100 ppm | Eater lower (AB control vs. A vanillin, p = 0.3416) Hymenoptaecin NS (p = 0.4622) Vitellogenin NS (p = 0.7257) | DWV NS (p = 0.7720) Trypanosome NS (p = 0.1548) VDV lower (A control vs. AB vanillin p = 0.2301) Gilliamella lower (A control vs. ABC vanillin, p = 0.0706) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boncristiani, D.L.; Tauber, J.P.; Palmer-Young, E.C.; Cao, L.; Collins, W.; Grubbs, K.; Lopez, J.A.; Meinhardt, L.W.; Nguyen, V.; Oh, S.; et al. Impacts of Diverse Natural Products on Honey Bee Viral Loads and Health. Appl. Sci. 2021, 11, 10732. https://doi.org/10.3390/app112210732
Boncristiani DL, Tauber JP, Palmer-Young EC, Cao L, Collins W, Grubbs K, Lopez JA, Meinhardt LW, Nguyen V, Oh S, et al. Impacts of Diverse Natural Products on Honey Bee Viral Loads and Health. Applied Sciences. 2021; 11(22):10732. https://doi.org/10.3390/app112210732
Chicago/Turabian StyleBoncristiani, Dawn L., James P. Tauber, Evan C. Palmer-Young, Lianfei Cao, William Collins, Kyle Grubbs, Jillian A. Lopez, Lyndel W. Meinhardt, Vy Nguyen, Sungtaek Oh, and et al. 2021. "Impacts of Diverse Natural Products on Honey Bee Viral Loads and Health" Applied Sciences 11, no. 22: 10732. https://doi.org/10.3390/app112210732
APA StyleBoncristiani, D. L., Tauber, J. P., Palmer-Young, E. C., Cao, L., Collins, W., Grubbs, K., Lopez, J. A., Meinhardt, L. W., Nguyen, V., Oh, S., Peterson, R. J., Zamora, H., Chen, Y., & Evans, J. D. (2021). Impacts of Diverse Natural Products on Honey Bee Viral Loads and Health. Applied Sciences, 11(22), 10732. https://doi.org/10.3390/app112210732