1. Background
Cardiovascular disease (CVD) is one of the leading causes of morbidity and mortality worldwide. It accounts for approximately 17.9 million deaths each year [
1]. Despite guideline-based treatment, patients with CVD (due to patients’ individual variability, environmental factors etc.) continue to experience a variety of cardiovascular events. Unfortunately, the current pandemic situation may only aggravate the incidence of CVD [
2]. Infection with the SARS-CoV-2 virus may result in endothelial dysfunction, organ hypoxia and thrombosis [
3,
4,
5].
Anticoagulation therapy is used in CVD treatment, and, nowadays, it may lead to a better prognosis in severe COVID-19 patients [
6]. Anticoagulants are usually prescribed to prevent various thromboembolic complications (such as pulmonary thrombosis and deep vein thrombosis after heart valve surgery). A new generation of oral anticoagulants, direct oral anticoagulants (DOACs), which include dabigatran, rivaroxaban, apixaban and edoxaban, has been introduced into practice, and they have already proved themselves as relatively effective, safe and convenient drugs for the prevention of thromboembolic complications [
7,
8,
9]. Rivaroxaban is an oral direct inhibitor of activated coagulation factor X (FXa), which can be administered in a simple, fixed-dose regimen, with drug dosage based on the patient’s body mass, age and sex [
7]. Rivaroxaban is a fast-acting (from the first dose) target-specific anticoagulant that inhibits both free and bound activated serine proteases, such as FXa. FXa is common to both internal and external coagulation activation pathways, so rivaroxaban can effectively impede the conversion of prothrombin to thrombin in the prothrombinase complex and inhibit further coagulation processes [
10].
After the administration of rivaroxaban, about 90% of it binds to plasma proteins (mainly albumin) and may circulate unchanged in the bloodstream. Rivaroxaban is eliminated from the body by two routes: unchanged or metabolized by the liver. Approximately one-third (35%) of the administered dose is excreted in the urine as unchanged active substance. The remaining two-thirds of the drug dose is metabolized by cytochrome P450 (CYP) family enzymes (CYP3A4/5, CYP2J2) or by other enzymes from CYP-independent pathways [
11,
12]. CYP3A4 and CYP2J2 are responsible for about one-third of drug elimination (18% and 14%, respectively). In addition to this oxidative biotransformation of rivaroxaban by CYP enzymes, non-CYP-related hydrolysis of amide bonds occurs, resulting in 14% of metabolism of rivaroxaban [
13]. It is known that the concentrations of rivaroxaban and its metabolites may vary depending on the activity of the cytochrome P450 enzymes [
11]; however, it is not clear how the therapeutic effect of rivaroxaban depends on the activity of these enzymes during specific conditions (such as inflammation).
Rivaroxaban shows a pleiotropic effect. Recent research data suggest that FXa possesses a function in the process of coagulation; additionally, it may have a proinflammatory effect [
14]. It was assumed that the inhibition of FXa with rivaroxaban may induce anti-inflammatory responses and protect the vascular endothelium from inflammation. A recent study showed that treatment with rivaroxaban is associated with a reduction in a key inflammation and coagulation biomarkers, resulting in decreased levels of D-dimer, the thrombin–antithrombin III complex (TAT), high-sensitivity C-reactive protein (hs-CRP) and high-sensitivity interleukin-6 (hs-IL-6) [
15]. Due to its anti-inflammatory effect, rivaroxaban may also have a potential pleiotropic role in patients infected with SARS-CoV-2 [
14].
It was mentioned previously that rivaroxaban is used as a fixed-dose drug, which means it does not require dosage adjustment. However, it is well known that patients have interindividual variabilities; thus, a more precise approach may be required to overcome possible variabilities in therapeutic drug effects [
16]. The study conducted by Miklič et al., 2019, proved that rivaroxaban concentrations in blood plasma differ from patient to patient [
17]. Interindividual variations in concentrations of rivaroxaban are a result of the metabolism of hepatic CYP 450 enzymes [
13]. In turn, the function of CYP 450 enzymes involved in the hepatic metabolism of exogenous compounds strongly depends on genetic variations, transcriptional regulation, inflammation and other factors [
11,
18,
19,
20,
21].
It is worth noting that alterations of CYP 450 enzyme activity might depend on environmental factors, such as drugs and food [
22,
23,
24]. Therefore, treatment with DOACs carries the risk of bleeding or a decreased therapeutic effect. Rivaroxaban co-administration with inhibitors of CYP 450 enzymes should be avoided, as they may lead to an increased plasma concentration and risk of bleeding. The opposite effect is observed if strong inducers of CYP 450 enzymes, such as rifampicin, carbamazepine and phenobarbital, are administered [
22,
23]. Under fasting conditions, the bioavailability of rivaroxaban was determined to be decreased. Thus, it might increase the risk of thrombosis [
25]. Dietary supplements and herbs can interact with anticoagulants. Patients taking rivaroxaban are at particular risk of interaction with dietary supplements, as there are no current governmental regulations or voluntary programs that address dietary supplement interactions with prescription drugs [
26]. It was determined that a strong CYP 450 enzyme inducer, St. John’s wort, decreases plasma concentrations of all available DOACs [
24,
27]. Thus, all of the above-mentioned factors emphasize the need for a clinically significant impact on CYP 450 enzyme identification.
The transcriptional regulation of gene expression is one of the major biological processes that coordinate and alter cellular activity after various intra- and extra-cellular signals, such as drug molecules. The expression of CYP 450 is controlled by transcription factors such as CAR (constitutive androstane receptor), PXR (pregnane X receptor), RXR (retinoid X receptor), PPARα (peroxisome proliferator-activated receptor) and HNF4α [
19,
20]. During inflammation, the gene transcription of major drug-metabolizing enzymes belonging to P450 is repressed through IL-6 and IL-1β action [
21]. Proinflammatory cytokines may cause a phenomenon better known as transient phenoconversion with the downregulation of drug metabolism. During inflammation, extensive metabolizers may become phenotypically poor metabolizers [
28].
It is known that enzyme CYP4F2 is activated during processes of inflammation and thrombosis [
29]. In humans, CYP4F2 oxidizes naturally occurring arachidonic acid to 20-hydroxyeicosatetraenoic acid (20-HETE). Additionally, CYP4F2 participates in the resolution of inflammation by reducing the level of leukotriene B4 [
30]. Previously published studies showed that CYP4F2 has an impact on warfarin dosage during treatment with the anticoagulant warfarin [
31]. Our recent works added new data—enzyme CYP4F2 has an impact on the antiplatelet effect in patients with stable and unstable cardiovascular disease and diabetes treated with antiplatelets ticagrelor and clopidogrel [
32,
33]. The study, which included 667 patients with acute coronary syndromes and treated with ticagrelor or clopidogrel dual antiplatelet therapy after stent implantation, showed higher CYP4F2 activity in patients without diabetes and in patients receiving ticagrelor [
32]. Another study showed that the concentration of CYP4F2 was lower in patients with angina pectoris compared to healthy subjects [
33]. These studies carried out on the antiplatelets ticagrelor and clopidogrel and on the anticoagulant warfarin [
31] showed a significant impact of CYP4F2 enzyme activity on drug effects. Therefore, there was a possibility that direct oral anticoagulants might have acted on the CYP4F2 enzyme. The endothelial lining of blood vessels is a barrier whose damage is directly involved in thrombosis and inflammation; thus, we decided to evaluate the activity of a novel anticoagulant, rivaroxaban, on the CYP4F2 and transcription factors in HUVEC cells.
2. Methods
2.1. Cell Culture Cultivation
The commercial cell line of human umbilical vein endothelial cells (HUVECs) obtained from Gibco (Gibco by Life Technologies, Carlsbad, CA, USA) was used for the study. HUVECs were cultured in Medium 200 (Gibco by Life Technologies, Carlsbad, CA, USA) supplemented with Large Vessel Endothelial Supplement (LVES 50X, Gibco by Life Technologies, Carlsbad, CA, USA) and antibiotic penicillin/streptomycin solution (Gibco by Life Technologies, Carlsbad, CA, USA). Cells were grown in 6-well cell culture plates (0.3 × 106 cells/well, Nunclon Delta Surface, Thermo Fisher Scientific, Waltham, MA, USA) and incubated at 37 °C, 5% CO2, under maintained humidity conditions. The enzymatic detachment of HUVECs was carried out using TrypLe Express reagent (Gibco by Life Technologies, Carlsbad, CA, USA). By performing the trypan blue exclusion test, cell concentration and viability were estimated with Countess II FL automatic cell counter (Invitrogen, Waltham, MA, USA). Cell growth was assessed using the EVOS XL Core cell imaging system (Invitrogen, Waltham, MA, USA). All experiments were conducted with HUVECs in passages 3–6.
2.2. Treatment with Rivaroxaban
Rivaroxaban stock solution was prepared using a 20 mg Xarelto tablet (Bayer, Leverkusen, Germany) dissolved in sodium chloride 0.9 mg/mL solution and albumin at 37 °C. Albumin solution was prepared by dissolving 100 mg of bovine serum albumin (BSA) in sodium chloride. The drug and albumin solutions were mixed, as it is known that serum albumin binding for rivaroxaban is high (92–95%). HUVECs were treated with 0.1–10 μM rivaroxaban solution and incubated for 24 h.
2.3. Treatment with Dexamethasone
Glucocorticoid dexamethasone (1 mg/mL) was used to induce transcription factors HNF4α, PXR and CAR in HUVECs. HUVECs were treated with 1 μM rivaroxaban solution and incubated for 24 h.
2.4. Detection of Rivaroxaban and Its Metabolites Using UPLC-ESI-MS Method
HUVEC growth medium was collected after 24 h of treatment with rivaroxaban, and 1.5 mL of the initial medium was concentrated under vacuum at 65 °C until the volume reached 300 μL. The medium was transferred to auto-sampler vials with inserts.
Chromatographic separation of rivaroxaban and its metabolites was performed as follows: The mobile phase consisted of the following solvents (eluents): 0.1% formic acid (A) and acetonitrile (B) in a ratio of 75A/25B. The flow rate was 0.4 mL/min. Ultra-performance liquid chromatography analysis was carried out using an Acquity H-class UPLC system (Waters, Milford, MA, USA). Precolumn ACQUITY UPLC BEH C18 VanGuard (1.7 µm, 2.1 mm × 5 mm) and column ACQUITY UPLC BEH C18 (1.7 µm, 2.1 mm × 100 mm) were used for the study. Triple quadrupole mass spectrometer (Xevo TQD, Waters, Milford, MA, USA) equipped with an electrospray ionization source was used. The mass spectrometer was operated in positive ion mode.
2.5. Detection of a New Compound Using LC-QTOF-MS Method
The medium of the treated HUVEC cells was also analyzed using liquid chromatography quadrupole time-of-flight mass spectrometry (LC-QTOF-MS). The medium was concentrated and transferred to auto-sampler vials with inserts.
Chromatographic analyses were performed using ACQUITY UPLC BEH C18 column (1.7 µm, 2.1 mm × 100 mm). The mobile phase was composed of 0.1% formic acid (A) and acetonitrile (B) in a ratio of 75A/25B. The flow rate was 0.4 mL/min. For QTOF-MS operated in positive ion mode, the parameters were set as follows: capillary and nozzle voltage were 4000 V and 500 V, respectively; drying gas temperature was 200 °C; and gas flow rate was 10 L/min. The mass spectrometer scan range was 80–1200 m/z.
2.6. Transfection of HUVECs with hsa-miR-24-3p
In total, 3.0 × 105 cells were seeded into 6-well plates 18–24 h prior to transfection. Growth medium was replaced with antibiotic-free medium before transfection. Synthetic miRNA mimics hsa-miR-24-3p, positive control (hsa-miR-1-3p) and negative control miRNA (Ambion by Life Technologies, Waltham, MA, USA) were transfected into cells following the Invitrogen by Life Technologies transfection protocol.
Forward transfection was performed using Lipofectamine RNAiMAX transfection reagent (Invitrogen, Waltham, MA, USA). Opti-MEM medium (Gibco by Life Technologies, Carlsbad, CA, USA) was used to prepare mixtures of Lipofectamine and miRNAs (hsa-miR-24-3p, positive and negative control mimics). miRNAs were diluted so that the final concentration in the well was 120 nM. After 24 h of transfection with miRNA mimics, cells were harvested for further quantitative PCR analysis and enzyme-linked immunosorbent assay.
2.7. RNA Isolation
After transfection or treatment with rivaroxaban and dexamethasone, the medium was removed, and HUVEC cells were trypsinized, harvested and washed with cold PBS before RNA extraction. Total RNA from the cell suspension was extracted using mirVana miRNA Isolation Kit (Invitrogen, Waltham, MA, USA) according to the manufacturer’s recommendations. The principle of the kit is based on the organic extraction and further immobilization of total RNA on glass fiber columns. Cells’ suspension samples for total RNA extraction must contain between 102 and 107 cells. RNA concentration and purity were determined using NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) by measuring the absorbance of UV light at 260 nm (A260) and absorbance ratios (A260/A280; A260/A230).
2.8. cDNA Synthesis
Reverse transcription (RT) was performed using High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA). Prepared samples were transferred to Biometra Tprofessional (Biometra, Burladingen, Germany) thermocycler, where the reaction was carried out according to the following program: 10 min at 25 °C, 2 h at 37 °C and 5 min at 85 °C.
2.9. qPCR Analysis
Quantitative polymerase chain reaction (qPCR) was used to evaluate the expression of genes after exposure to miRNA molecules and rivaroxaban (dexamethasone also). Gene expression experiments were carried out on an ABI 7900HT Fast Real-Time PCR System (Applied Biosystems, Waltham, MA, USA) in MicroAmp Optical 96-well plates (Applied Biosystems, Waltham, MA, USA) using Maxima SYBR Green/ROX qPCR Master Mix 2× (Thermo Fisher Scientific, Waltham, MA, USA). Custom-made primers (Thermo Fisher Scientific, USA) were used to amplify the cDNA, specifically amplifying fragments of
CYP4F2,
HNF4α,
PXR,
CAR and
TFRC genes (sequences of the primers shown in
Table 1). Thermal cycling program parameters used in the qPCR are presented in
Table 2. Relative changes in the expression of genes were evaluated using the 2
−∆∆Ct method. The expression level of transferrin receptor (
TFRC) was used as an endogenous control for data normalization.
2.10. Detection of CYP4F2 Protein
Cell lysates were prepared after detached HUVECs were resuspended in cold PBS (4 °C) and centrifuged at 1500× g for 5 min. Then, cells were resuspended in 300 μL of lysis buffer and kept on ice for 10 min. After incubation, the tubes were centrifuged at 10,000× g (4 °C) for 10 min, and the collected supernatants were stored at −80 °C until further analysis. CYP4F2 enzyme concentration in lysates was determined using a commercial Enzyme-linked Immunosorbent Assay Kit for Cytochrome P450 4F2 (CYP4F2) (Cloud-Clone Corp., Houston, TX, USA). The analysis was performed according to the ELISA kit SEL399Hu protocol provided by the manufacturer. ELISA plate for optical densities was analyzed using StatFax 4200 microplate reader (Awaraness Technologies, Palm City, FL, USA) with a primary wavelength of 450 nm; calculations were performed using a standard calibration curve.
5. Discussion
The main tasks in our study were the following. (i) To analyze the metabolism of rivaroxaban in HUVECs in vitro. To accomplish this task, an analysis of rivaroxaban and its metabolites in cell culture media was performed using both UPLC-ESI-MS and UPLC-QTOF-MS methods. (ii) To determine the impact of rivaroxaban and the hsa-miR-24-3p mimic on the expression of CYP4F2 and its potential transcription factors, HNF4α, PXR and CAR, and CYP4F2 protein levels in endothelial cell cultures.
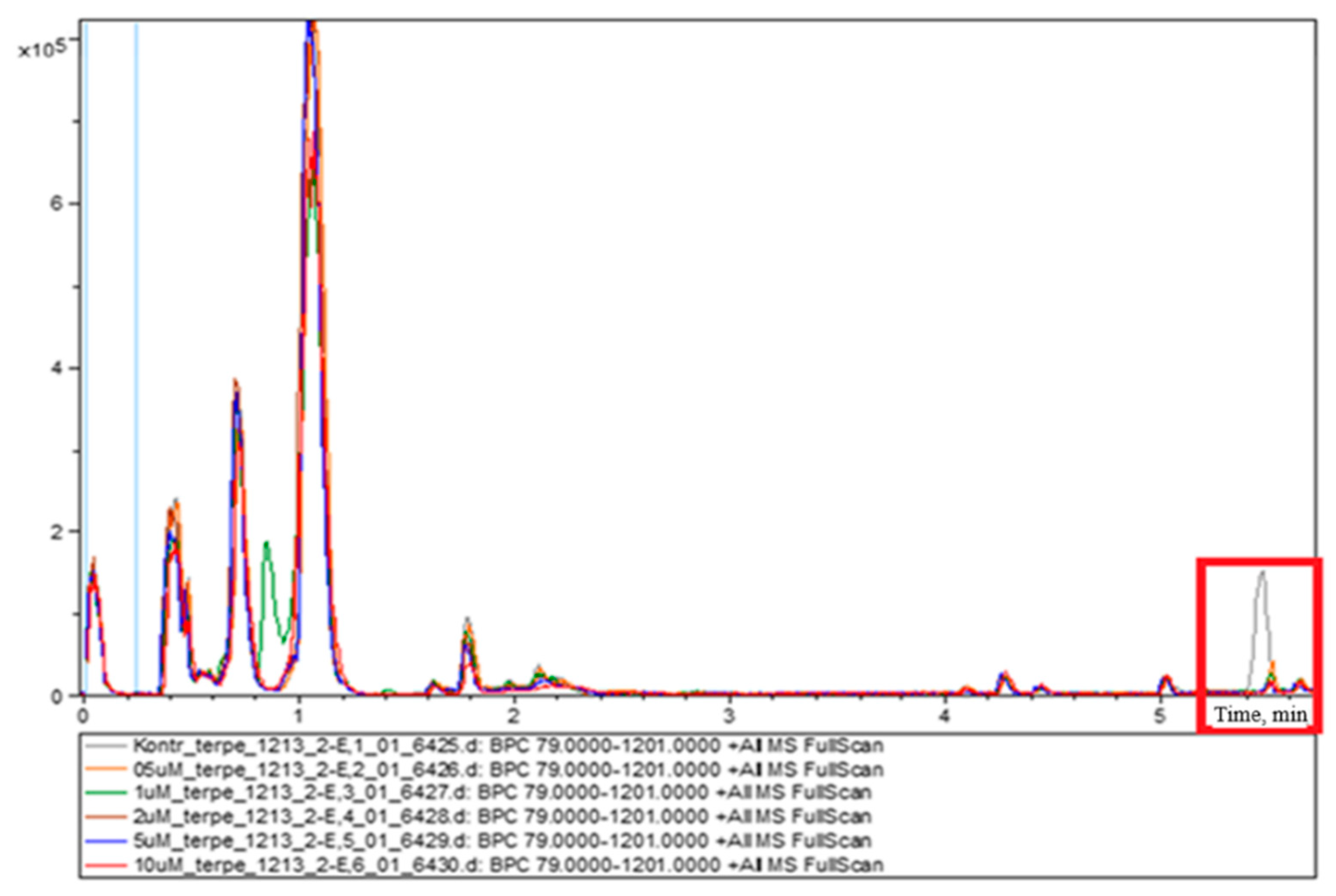
The commercial human umbilical vein endothelial cell culture, HUVEC, was chosen as a model system, as it is convenient for studying various aspects of endothelial function, including angiogenesis, oxidative stress, hypoxia and inflammatory mechanisms under normal and pathological conditions associated with CVD pathogenesis. Rivaroxaban was detected in all HUVECs studied and treated with different concentrations: 0.1, 0.2, 0.5, 1, 2, 5 and 10 μM. The selection of rivaroxaban concentrations was based on the studies of Mueck et al. and Sennesael et al. for the detection of rivaroxaban’s peak concentration in plasma 2–4 h after 20 mg tablet administration. The concentrations presented in ng/mL were converted to molar concentrations (μM) [
12,
34]. According to the study of Çelebier et al., 2013, the retention time of rivaroxaban during the performance of reversed-phase liquid chromatography was 2.37 min [
35]. A similar retention time (2.34 min) was determined in our study. Based on the
m/z of metabolites presented in other studies, the following metabolites were identified in the cell media of HUVECs from our study: M-1, M-2, M-5, M-8, M-10, M-11 and M-18 [
36]. Rivaroxaban and its metabolites are usually analyzed in liver cells [
35,
36]. It is worth noting that rivaroxaban is also metabolized in HUVECs, as the results of this study revealed.
The HUVEC culture medium samples treated with different concentrations of rivaroxaban were analyzed using the LC-QTOF-MS system. The analysis revealed a peak at 5.4–5.5 min, which decreased with an increase in the concentration of rivaroxaban. No data were found about the newly identified substance associated with rivaroxaban. The molecular formula of the new compound is C
22H
43NO. The latter compound, identified as erucamide (13-docosenamide), is known to act on the central nervous system, and it may promote angiogenesis; it also has anti-cancer and antimicrobial properties [
37]. According to the study of Tamilmani et al., this compound might be produced by the host–bacteria interaction when glucose is present [
38]. We hypothesize that rivaroxaban may also influence the metabolism of HUVEC cells and may alter it. It has been determined that erucamide production is lower in samples in the presence of higher rather than lower rivaroxaban concentrations. Future research is required for a better understanding of the correlation between erucamide production and rivaroxaban concentrations.
This study also evaluated changes in the mRNA level and the concentrations of the cytochrome P450 family enzyme CYP4F2, known as ω-hydroxylase, in HUVECs induced by treatment with different concentrations of rivaroxaban. Álvarez et al., who studied the effect of rivaroxaban on HUVECs, had determined that rivaroxaban, in a concentration-dependent manner, increases cell viability, growth and migration via the urokinase plasminogen activator by inducing its expression [
39]. The team also determined that rivaroxaban neutralizes the FXa-induced expression of inflammatory genes and platelet adhesion in HUVECs. These results demonstrate the ability of rivaroxaban to improve endothelial function in CVD. It is known that rivaroxaban exhibits anti-inflammatory properties. The exposure of HUVECs to rivaroxaban resulted in a significant concentration-dependent downregulation of inflammatory genes
VCAM-1,
ICAM-1 and
IL-8 [
40]. Another study investigating the effects of oxidized cholesterol on HUVECs showed that rivaroxaban may reduce elevated ROS levels in cells [
41]. ROS is involved in inflammatory processes and induces the development of atherosclerosis. The results of our study demonstrated the potential antioxidant properties and benefits of rivaroxaban in the prevention of atherosclerosis. The amount of CYP4F2 enzyme is found to be optimal 24 h after the onset of inflammation [
42,
43,
44]. In our study, after treatment with different doses of rivaroxaban (0.1, 0.2, 0.5, 1, 2, 5 and 10 μM), the expression of the CYP4F2 gene in HUVECs was downregulated by 1.44-, 1.71-, 1.89-, 1.93-, 2.05-, 2.4- and 2.72-fold in a concentration-dependent manner compared to control cells. The downregulation of
CYP4F2 expression decreases the amount of gene transcript and the rate of ω-hydroxylase synthesis of its encoded protein. ω-hydroxylase is one of the major enzymes in the CYP family involved in the metabolism of arachidonic acid to form biologically active eicosanoids, such as 20-HETE. CYP4F2 also metabolizes leukotriene B4 (LTB4), which is formed/synthesized during the development of inflammation [
42,
43,
44]. No other studies have been performed to evaluate the effect of rivaroxaban on the enzyme ω-hydroxylase in endothelial cells.
The effect of hsa-miR-24-3p on target genes was further evaluated in this study. A potential hsa-miR-24-3p gene target,
HNF4α, was selected for this study after a detailed analysis of information found on miRNA target databases (TargetScan v.7.2, miRTarBase 7.0) and PubMed. Takagi et al. were the first to describe the regulation of
HNF4α by miR-24 in vitro [
45]. The potential recognition elements of miR-24 were identified in
HNF4α coding and the 3′ UTR region. The study showed that miR-24 also reduced the expression of various gene targets, including cytochrome P450 7A1 and 8B1 isoforms, by inhibiting
HNF4α expression. The fact that miR-24-3p acts as an effector molecule of
HNF4α and indirectly contributes to the regulation of other genes has also been confirmed by the Salloum-Asfar study [
46]. After the analysis of the previously mentioned miRNA databases, it was determined that the 3′ UTR region of
CYP4F2 lacks hsa-miR-24-3p binding sites. However, according to the ENCODE Transcription Factor Targets database,
HNF4α can act as a
CYP4F2 transcription factor too.
HNF4α binding sites were identified in the promoter of
CYP4F2, but no functional studies have been performed yet [
47]. These results raised the hypothesis that hsa-miR-24-3p may indirectly affect
CYP4F2 expression by acting through the transcription factor
HNF4α. PXR and CAR are regulated by
HNF4α [
48,
49]. In turn/additionally, we hypothesized that
CYP4F2 expression in extra-hepatic cells might also be controlled by PXR or CAR transcription factors.
Detailed studies of miR-24-3p function in humans have not been conducted, but its association with CVD is well known. It has been shown that miR-24-3p inhibits angiogenesis in the myocardial infarction zone. Decreased survival, proliferation and migration of cells and angiogenic properties have been observed after the transfection of endothelial cells with this miRNA [
50,
51]. Increased miR-24 expression inhibits the expression of endothelial inflammatory factor
TNF-α, which contributes to the regulation of inflammation via the NF-κB signaling pathway. It is worth noting that patients with atherosclerosis had significantly lower miR-24 expression in their blood samples compared to healthy patients [
50].
According to our previously published data, miR-24-3p expression in blood plasma was 4.4-fold higher in patients with stable angina than in healthy controls [
33]. In the current study, transfection with the hsa-miR-24-3p mimic after 24 h of incubation resulted in a 1.7-fold decrease in
CYP4F2 expression in HUVECs compared to the negative control. However, after transfection with the hsa-miR-24-3p mimic, the expression of
HNF4α, PXR and
CAR was not detected, suggesting that these transcription factors are not involved in the regulation of CYP genes in endothelial cells and that other regulatory mechanisms might possibly be involved.
The evaluation of glucocorticoid-induced
HNF4α transcriptional activity was performed by applying treatment with 1 μM dexamethasone to HUVECs.
HNF4α, PXR and
CAR expression could not be determined. However, treatment with dexamethasone resulted in a change in
CYP4F2 expression; dexamethasone downregulated the expression of
CYP4F2 1.96-fold. Dexamethasone was identified as a classical inducer of cytochrome P450 subfamily enzymes (CYP1A, CYP2B, CYP3A and CYP4A), but there are no data available on how dexamethasone acts on the CYP4F subfamily or CYP4F2 gene [
52].
An analysis of CYP4F2 protein quantity was performed by ELISA to determine the effect of both the new-generation anticoagulant rivaroxaban and hsa-miR-24-3p at the protein level. Transfection with hsa-miR-24-3p in HUVECs resulted in a 6.4-fold decrease in CYP4F2 protein levels in cells. However, exposure to rivaroxaban at different concentrations did not significantly alter CYP4F2 protein levels in HUVECs. The expression of the altered gene at the mRNA level may change the amount of the corresponding protein in the cell, as gene expression at the mRNA and protein levels often varies disproportionately. The same mRNA molecule can be used for the synthesis of several hundred to several hundred thousand protein molecules, and various post-translational mechanisms contribute to this [
53].
The assessment of rivaroxaban metabolism in vitro using HUVEC culture as a model system could be seen as a limitation of the experiment. Most similar studies conducted previously cultivated cell cultures using two-dimensional methods that do not always mimic the environment of cells in vivo [
54]. The metabolic profiling of rivaroxaban in HUVECs may differ from live organisms or three-dimensional cultures that have the advantage of cell-to-cell interactions.


 ,
, 



{kind=link}
{kind=link}