
Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Bacteria from Saline Soils
2.2. In Vitro Screening of Isolates for Plant Growth-Promoting (PGP) Activities
2.2.1. Phosphate Solubilization
2.2.2. Quantitative Estimation of Indole Acetic Acid
2.2.3. Siderophore Production
2.2.4. Production of Enzymes
2.2.5. Hydrogen Cyanide (HCN) Production
2.2.6. NH3 Production
2.2.7. Nitrogen Fixation
2.2.8. Antifungal Assay
2.3. Selection of Tolerant PGPB for In Vivo Plant Growth Promotion
2.4. Molecular Characterization of Salt-Tolerant PGPR
2.5. In Vitro and In Vivo Plant Growth Promotion by the Selected Bacterial Strains
2.6. In Vitro Assays
2.6.1. Plate Grown Arabidopsis Thaliana Seedlings Inoculation
2.6.2. Wheat Seed Germination in the Presence of PGPB under Salt Stress
2.7. In Vivo Assays
2.7.1. Wheat Inoculation
2.7.2. Photosynthetic Pigments and Biochemical Markers
Photosynthetic Pigments Concentration Estimation
Total Soluble Sugars Content
Proline Content Estimation
Protein Content
Lipid Peroxidation
2.8. Antioxidant Enzyme Assays
2.8.1. Guaiacol Peroxidase Activity
2.8.2. Ascorbate Peroxidase Activity
2.8.3. Catalase Enzyme Assay
2.8.4. Superoxide Dismutase Enzyme Assay
2.9. Statistical Analysis
3. Results
3.1. Isolation of Bacteria
3.2. Plant Growth-Promoting Traits of Isolates
3.3. Selection of Tolerant PGPB for In Vivo Plant Growth Promotion
3.4. Molecular Characterization of Salt-Tolerant PGPR
3.5. In Vitro Assays
3.5.1. Arabidopsis Thaliana Inoculation
3.5.2. Seed Germination of Wheat in the Presence of PGPR under Salt Stress
3.6. In Vivo Assays
Effects of Bacterial Inoculation on Wheat Plants Growth and Development in Different Soils
3.7. Effects on Photosynthetic Pigments and Biochemical Markers
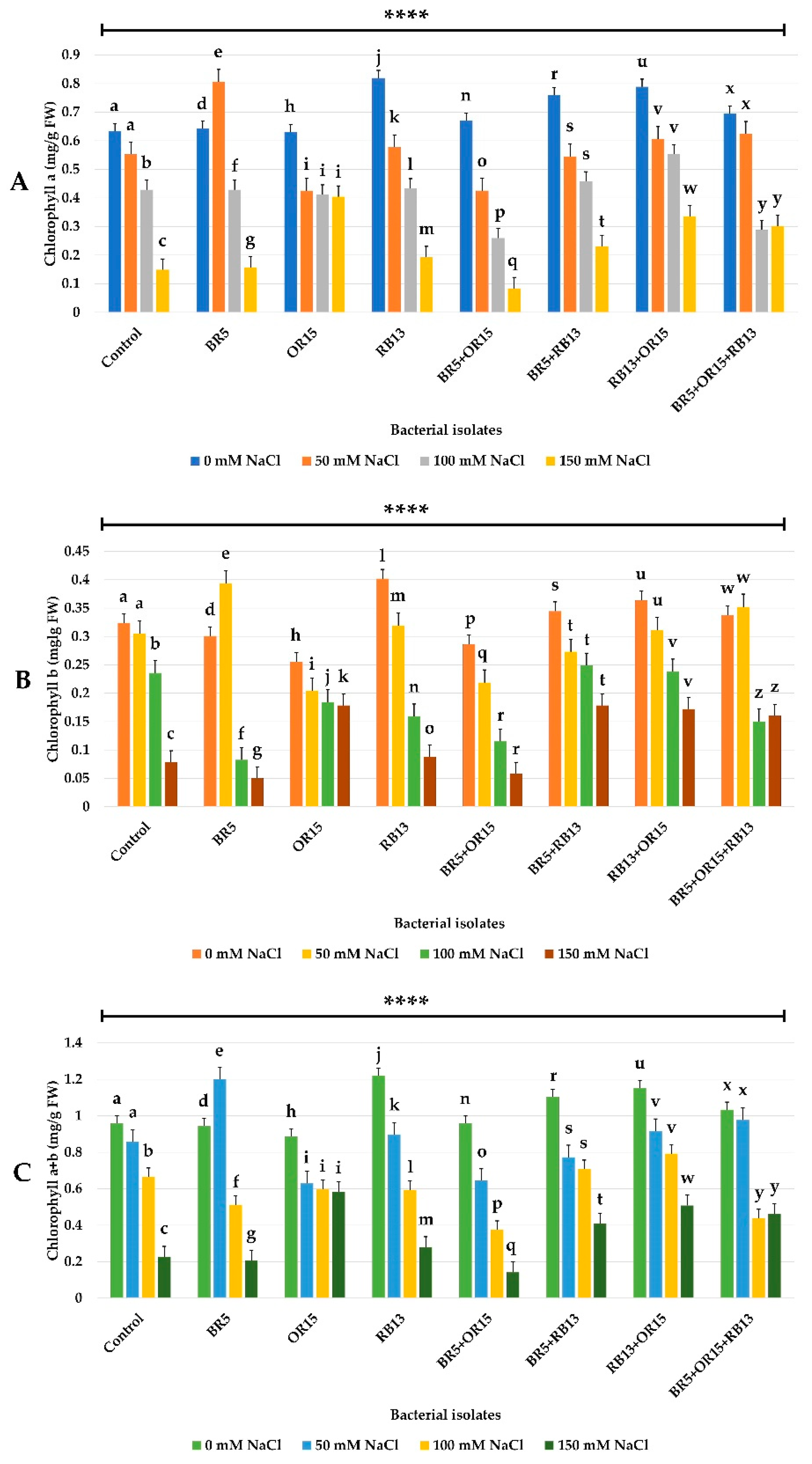
3.7.1. Chlorophyll Content
3.7.2. Total Soluble Sugars Content
3.7.3. Protein Content
3.7.4. Proline Content Estimation
3.7.5. Lipid Peroxidation
3.7.6. Effects on Antioxidant Enzymes Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rekik, I.; Chaabane, Z.; Missaoui, A.; Chenari Bouket, A.; Luptakova, L.; Elleuch, A.; Belbahri, L. Effects of untreated and treated wastewater at the morphological, physiological and biochemical levels on seed germination and development of sorghum (Sorghum bicolor (L.) Moench), alfalfa (Medicago sativa L.) and fescue (Festuca arundinacea Schreb.). J. Hazard. Mater. 2017, 326, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Frikha-Dammak, D.; Zarai, Z.; Najah, S.; Abdennabi, R.; Belbahri, L.; Rateb, M.E.; Mejdoub, H.; Maalej, S. Antagonistic properties of some halophilic thermoactinomycetes isolated from superficial sediment of a solar saltern and production of cyclic antimicrobial peptides by the novel isolate Paludifilum halophilum. BioMed Res. Int. 2017, 2017, 1205258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherif-Silini, H.; Thissera, B.; Chenari Bouket, A.; Saadaoui, N.; Silini, A.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Yahiaoui, B.; et al. Durum wheat stress tolerance induced by endophyte Pantoea agglomerans with genes contributing to plant functions and secondary metabolite arsenal. Int. J. Mol. Sci. 2019, 20, 3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slama, H.B.; Cherif-Silini, H.; Chenari Bouket, A.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.N.; Luptakova, L.; Triki, M.A.; Vallat, A.; et al. Screening for Fusarium antagonistic bacteria from contrasting niches designated the endophyte Bacillus halotolerants as plant warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Pegler, J.L.; Grof, C.P.L.; Eamens, A.L. Profiling of the differential abundance of drought and salt stress-responsive microRNAs across grass crop and genetic model plant species. Agronomy 2018, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Lara, E.; Belbahri, L. SSU rRNA reveals major trends in oomycete evolution. Fungal Divers. 2011, 49, 93–100. [Google Scholar] [CrossRef]
- Luchi, N.; Ghelardini, L.; Belbahri, L.; Quartier, M.; Santini, A. Rapid detection of Ceratocystis platani inoculum by quantitative Real-Time PCR assay. Appl. Environ. Microbiol. 2013, 79, 5394–5404. [Google Scholar] [CrossRef] [Green Version]
- Bellabarba, A.; Fagorzi, C.; di Cenzo, G.C.; Pini, F.; Viti, C.; Checcucci, A. Deciphering the symbiotic plant microbiome: Translating the most recent discoveries on rhizobia for the improvement of agricultural practices in metal-contaminated and high saline lands. Agronomy 2019, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Belbahri, L.; Chenari Bouket, A.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative genomics of Bacillus amyloliquefaciens strains reveals a core genome with traits for habitat adaptation and a secondary metabolites rich accessory genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef]
- Mefteh, F.B.; Chenari Bouket, A.; Daoud, A.; Luptakova, L.; Alenezi, F.N.; Gharsallah, N.; Belbahri, L. Metagenomic insights and genomic analysis of phosphogypsum and its associated plant endophytic microbiomes reveals valuable actors for waste bioremediation. Microorganisms 2019, 7, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mefteh, F.B.; Frikha, F.; Daoud, A.; Chenari Bouket, A.; Luptakova, L.; Alenezi, F.N.; Al-Anzi, B.S.; Oszako, T.; Gharsallah, N.; Belbahri, L. Response surface methodology optimization of an acidic protease produced by Penicillium bilaiae isolate TDPEF30, a newly recovered endophytic fungus from healthy roots of date palm trees (Phoenix dactylifera L.). Microorganisms 2019, 7, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheffi, M.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A.; Belbahri, L. Olea europaea L. root endophyte Bacillus velezensis OEE1 counteracts oomycete and fungal harmful pathogens and harbours a large repertoire of secreted and volatile metabolites and beneficial functional genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhouib, H.; Zouari, I.; Abdallah, D.B.; Belbahri, L.; Taktak, W.; Triki, M.A.; Tounsi, S. Potential of a novel endophytic Bacillus velezensis in tomato growth promotion and protection against Verticillium wilt disease. Biol. Control 2019, 139, 104092. [Google Scholar] [CrossRef]
- Abdul-Baki, B.A.A.; Anderson, J.D. Relationship between decorboxylation of glutamic acid and vigor in soyabean seed. Crop Sci. 1973, 13, 227–234. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Phenol-sulfuric total sugar. Anal Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Carrillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: accumulation of proline and glycine betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Etesami, H.; Beattie, G.A. Mining halophytes for plant growth-promoting halotolerant bacteria to enhance the salinity tolerance of non-halophytic crops. Front. Microbiol. 2018, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Durán, R.E.; Méndez, V.; Rodríguez-Castro, L.; Barra-Sanhueza, B.; Salvà-Serra, F.; Moore, E.R.B.; Castro-Nallar, E.; Seeger, M. Genomic and physiological traits of the marine bacterium Alcaligenes aquatilis QD168 isolated from Quintero Bay, Central Chile, reveal a robust adaptive response to environmental stressors. Front. Microbiol. 2019, 10, 528. [Google Scholar] [CrossRef] [Green Version]
- Mapelli, F.; Marasco, R.; Rolli, R.E.; Barbato, M.; Cherif, H.; Guesmi, A.; Ouzari, I.; Daffonchio, D.; Borin, S. Potential for plant growth promotion of rhizobacteria associated with Salicornia growing in Tunisian hypersaline soils. Biomed. Res. Int. 2013, 248078. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Tsukanova, K.A.; Chebotar, V.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Zhou, N.; Zhao, S.; Tian, C.Y. Effect of halotolerant rhizobacteria isolated from halophytes on the growth of sugar beet (Beta vulgaris L.) under salt stress. FEMS Microbiol. Lett. 2017, 364, 11. [Google Scholar] [CrossRef]
- Abbas, G.; Saqib, M.; Akhtar, J. Interactive effects of salinity and iron deficiency on different rice genotypes. J. Plant Nutr. Soil Sci. 2015, 178, 306–311. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR). A review. Cogent Food Agric. 2016, 2, 1–19. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Berg, G.; Lindström, K.; Räsänen, L.A. Alleviation of salt stress of symbiotic Galega officinalis L. (Goat’s Rue) by co-inoculation of rhizobium with root colonizing Pseudomonas. Plant Soil 2011, 369, 453–465. [Google Scholar] [CrossRef]
- Banerjee, S.; Palit, R.; Sengupta, C.; Standing, D. Stress induced phosphate solubilization by ‘Arthrobacter’ sp. and ‘Bacillus’ sp. isolated from tomato rhizosphere. Aust. J. Crop Sci. 2010, 4, 378. [Google Scholar]
- Marasco, R.; Rolli, E.; Vigani, G.; Borin, S.; Sorlini, C.; Ouzari, H.; Zocchi, G.; Daffonchio, D. Are drought-resistance promoting bacteria cross-compatible with different plant models? Plant Signal Behav. 2013, 8, e26741. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Subramanian, S.; Smith, D.L. Plant endophytes promote growth and alleviate salt stress in Arabidopsis thaliana. Sci. Rep. 2020, 10, 12740. [Google Scholar] [CrossRef] [PubMed]
- Poupin, M.J.; Timmermann, T.; Vega, A.; Zuñiga, A.; González, B. Effects of the plant growth-promoting bacterium Burkholderia phytofirmans PsJN throughout the life cycle of Arabidopsis thaliana. PLoS ONE 2013, 8, e69435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantelin, S.; Desbrosses, G.; Larcher, M.; Tranbarger, T.J.; Cleyet-Marel, J.C.; Touraine, B. Nitrate-dependent control of root architecture and N nutrition are altered by a plant growth-promoting Phyllobacterium sp. Planta 2006, 223, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wei, W.; Zhu, W.; Su, L.; Xiong, Z.; Zhou, M.; Zheng, Y.; Zhou, D.X. Histone deacetylase AtSRT1 regulates metabolic flux and stress response in Arabidopsis. Mol. Plant 2017, 10, 1510–1522. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indole acetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Farah, A.; Iqbal, A.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar]
- Silini, A.; Silini-Cherif, H.; Ghoul, M. Effect of Azotobacter vinelandii and compatible solutes on germination wheat seeds and root concentrations of sodium and potassium under salt stress. Pakistan J. Biol. Sci. 2012, 15, 132–140. [Google Scholar] [CrossRef]
- Delshadi, S.; Ebrahimi, M.; Shirmohammadi, E. Influence of plant-growth-promoting bacteria on germination, growth and nutrients’ uptake of Onobrychis sativa L. under drought stress. J. Plant Interact. 2017, 12, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Zafar-ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC deaminase producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with biochar improve wheat productivity under drought stress. Agronomy 2019, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Baset Mia, M.A.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured musa plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- Mahmood, S.; Daur, I.; Al-Solaimani, S.G.; Ahmad, S.; Madkour, M.H.; Yasir, M.; Ali, Z. Plant growth promoting rhizobacteria and silicon synergistically enhance salinity tolerance of mung bean. Front. Plant. Sci. 2016, 7, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panwar, M.; Tewari, R.; Gulati, A.; Nayyar, H. Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol. Plant. 2016, 38, 278. [Google Scholar] [CrossRef]
- Rabhi, N.E.H.; Silini, A.; Cherif-Silini, H.; Yahiaoui, B.; Lekired, A.; Robineau, M.; Esmaeel, Q.; Jacquard, C.; Vaillant-Gaveau, N.; Clément, C.; et al. Pseudomonas knackmussii MLR6, arhizospheric strain isolated from halophyte, enhances salt tolerance in Arabidopsis thaliana. J. Appl. Microbiol. 2018, 125, 1836–1851. [Google Scholar] [CrossRef] [PubMed]
- Soliman, W.S.; Fujimori, M.; Tase, K.; Sugiyama, S. Heat tolerance and suppression of oxidative stress: Comparative analysis of 25 cultivars of the C3 grass Lolium perenne. Environ. Exp. Bot. 2012, 78, 10–17. [Google Scholar] [CrossRef]
- Kang, S.M.; Khan, A.; Waqas, M.; You, Y.H.; Kim, J.H.; Kim, J.G.; Hamayun, M.; Lee, L.J. Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Interact. 2014, 9, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Al-Erwy, A.S.; Al-Toukhy, A.; Bafeel, S.O. Effect of chemical, organic and bio fertilizers on photosynthetic pigments, carbohydrates and minerals of wheat (Triticum aestivum L.) irrigated with sea water. Int. J. Adv. Res. Biol. Sci. 2016, 3, 296–310. [Google Scholar]
- Afridi, M.S.; Amna, S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; Javed, M.T.; et al. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef]
- Simkin, A.J.; Lopez-Calcagno, P.E.; Raines, C.A. Feeding the world: improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 2019, 70, 1119–1140. [Google Scholar] [CrossRef] [Green Version]
- Tewari, T.N.; Singh, B.B. Sodicity induced changes in chlorophyll, nitrate, nitrite reductase, nucleic acids, proline and yield components of lentil. Plant Soil 1991, 136, 225–230. [Google Scholar] [CrossRef]
- Zhu, S.N.; Feng, P.Z.; Feng, J.; Xu, J.; Wang, Z.M.; Xu, J.L.; Yuan, Z.H. The roles of starch and lipid in Chlorella sp. during cell recovery from nitrogen starvation. Bioresour. Technol. 2018, 247, 58–65. [Google Scholar] [CrossRef]
- Perez-Montaño, F.; Alias-Villegas, C.; Bellogin, R.A.; del Cerro, P.; Espuny, M.R.; Jimenez-Guerrero, I.; Lopez-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: from microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talaat, N.B.; Shawky, B.T. Modulation of the ROS scavenging system in salt-stressed wheat plants inoculated with arbuscular mycorrhizal fungi. J. Plant Nutr. Soil Sci. 2014, 177, 199–207. [Google Scholar] [CrossRef]
- García, J.E.; Maroniche, G.; Creus, C.; Suárez-Rodríguez, R.; Ramirez-Trujillo, J.A.; Groppa, M.D. In vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growth of maize under drought stress. Microbiol. Res. 2017, 202, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.M.; Lee, I.J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- Han, Q.Q.; Lü, X.P.; Bai, J.P.; Qiao, Y.; Paré, P.W.; Wang, S.M.; Zhang, J.L.; Wu, Y.N.; Pang, X.P.; Xu, W.P.; et al. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front. Plant Sci. 2014, 5, 525. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR inoculation on growth and antioxidants status of wheat under saline conditions. Plant Biol. 2012, 4, 605–611. [Google Scholar] [CrossRef]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.S.P. Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front. Microbiol. 2017, 8, 1945. [Google Scholar] [CrossRef]
- Kumar, A.; Dubey, A. Rhizosphere Microbiome: Engineering bacterial competitiveness for enhancing crop production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria Sampling Site | Site | Halophytes | Electrical Conductivity (mS/cm) | pH |
|---|---|---|---|---|
| Bazer Sakhra Sebkha | Bazer Sakhra Chott (36.0505803° N, 5.6805708° E) | Suaeda mollis | 30.34 ± 0.525 | 8.055 ± 0.015 |
| Oum El Bouaghi Sebkha | Tinsilt Chott (35.8852354° N, 6.4414962° E) | Suaeda mollis | 22.95 ± 0.145 | 7.99 ± 0.14 |
| Baniou Sebkha | El Hodna Chott (35.479223° N, 4.366587° E) | Salsola tetrandra | 25.56 ± 0.465 | 7.995 ± 0.014 |
| Location Site | Electrical Conductivity (mS/cm) | pH | |
|---|---|---|---|
| Soil 1 (S1) | Setif (fertile soil) (36.195889° N, 5.364722° E) | 0.48 ± 0.034 | 8.075 ± 0.055 |
| Soil 2 (S2) | Bazer Sakhra Sebkha (periphery of the sebkha) (36.071722° N, 5.675417° E) | 3.81 ± 0.16 | 7.735 ± 0.064 |
| Soil 3 (S3) | Bazer Sakhra Sebkha (Mechtat Zaabib: away from Sebkha) (36.073579° N, 5.667538° E) | 2.8 ± 0.045 | 7.885 ± 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Appl. Sci. 2021, 11, 1034. https://doi.org/10.3390/app11031034
Kerbab S, Silini A, Chenari Bouket A, Cherif-Silini H, Eshelli M, El Houda Rabhi N, Belbahri L. Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Applied Sciences. 2021; 11(3):1034. https://doi.org/10.3390/app11031034
Chicago/Turabian StyleKerbab, Souhila, Allaoua Silini, Ali Chenari Bouket, Hafsa Cherif-Silini, Manal Eshelli, Nour El Houda Rabhi, and Lassaad Belbahri. 2021. "Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria" Applied Sciences 11, no. 3: 1034. https://doi.org/10.3390/app11031034
APA StyleKerbab, S., Silini, A., Chenari Bouket, A., Cherif-Silini, H., Eshelli, M., El Houda Rabhi, N., & Belbahri, L. (2021). Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Applied Sciences, 11(3), 1034. https://doi.org/10.3390/app11031034