
Antioxidant Production in Dunaliella




Abstract
:1. Introduction
2. Antioxidants
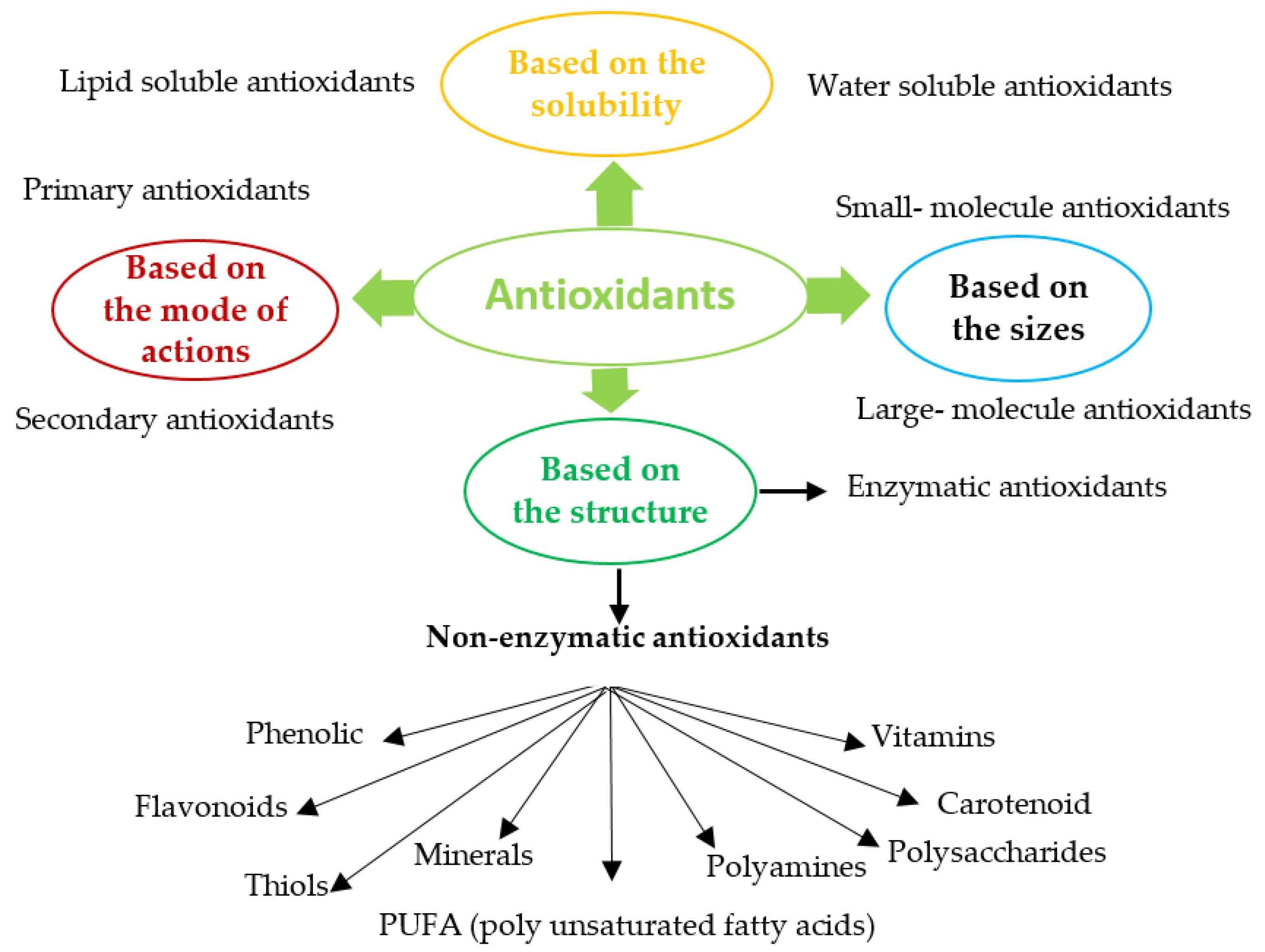
2.1. Classification of Antioxidants
2.2. Antioxidant Enzymes
2.3. Commercial Applications of Antioxidant Enzymes
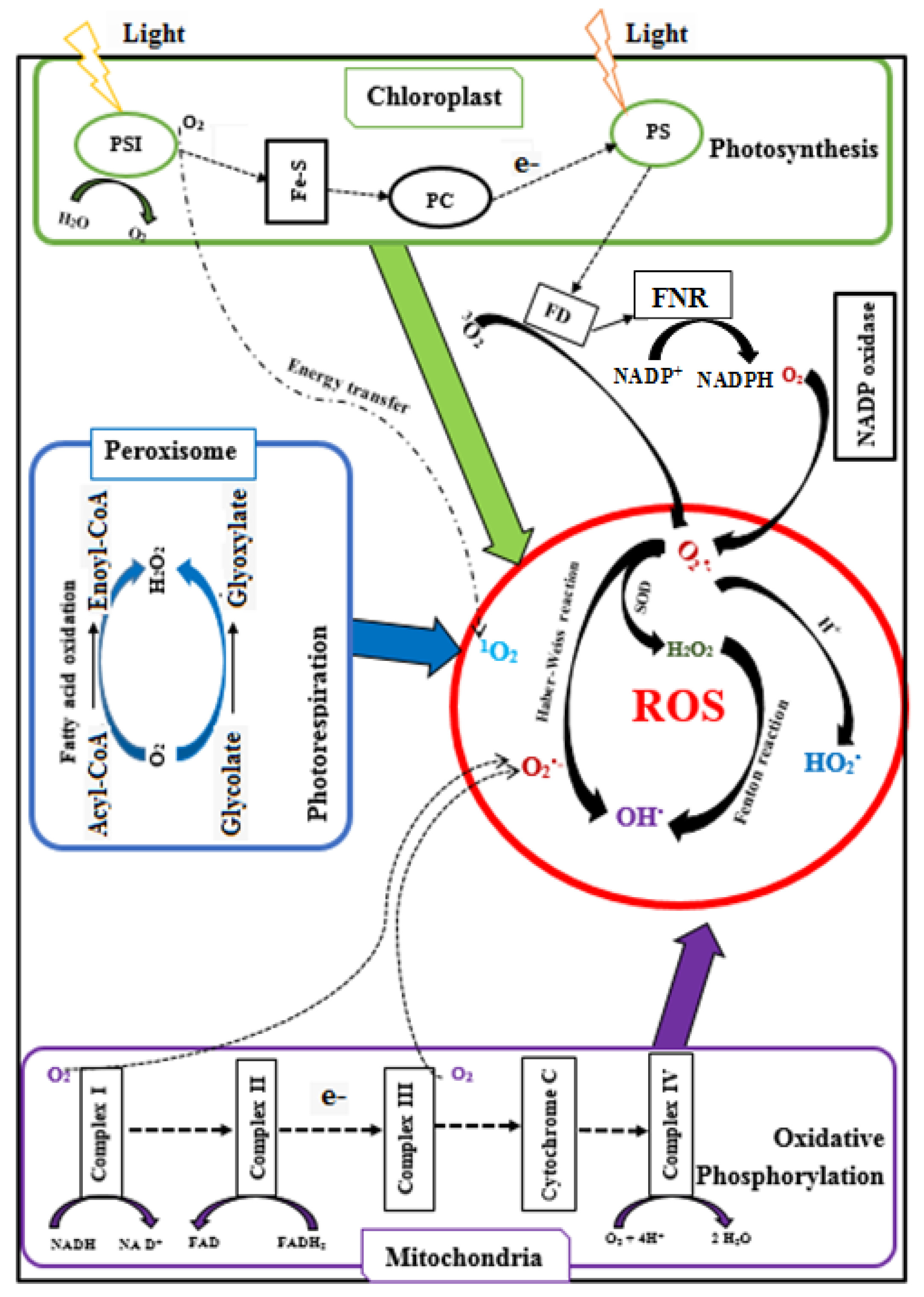
2.4. Generation and Detoxification of ROS
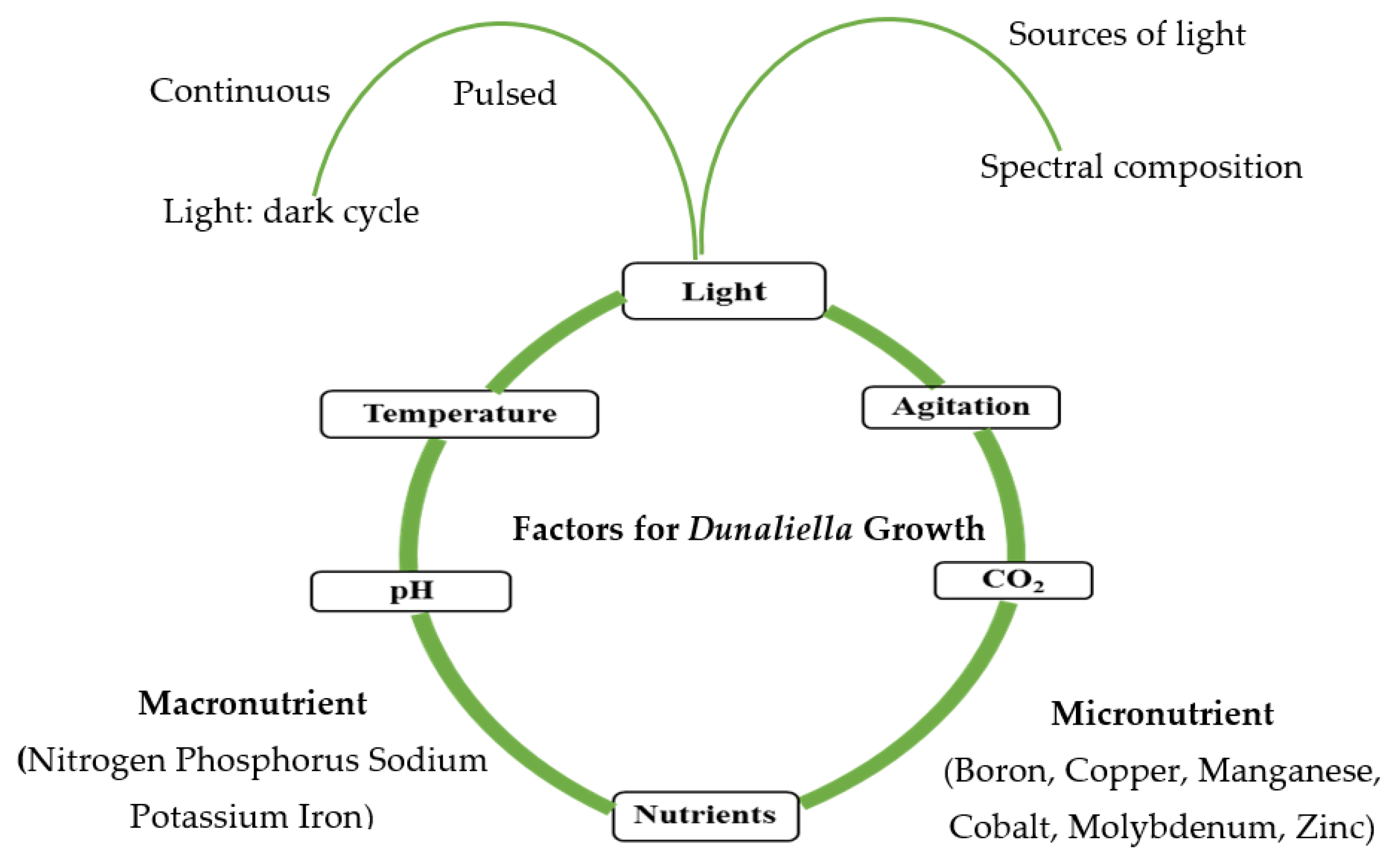
3. Cultivation Conditions
4. Tuning Antioxidant Enzyme Activity in Dunaliella
- i.
- Concentration and source nutrients (e.g., carbon, nitrogen, phosphorus, iron)
- ii.
- Concentration of trace elements (e.g., Zn2+, Cu2+)
- iii.
- Chemicals (e.g., phenol, H2O2)
- iv.
- Heavy metals (e.g., Hg2+, Cd2+)
- i.
- Irradiation level
- ii.
- Types of light (direct sunlight, artificial light sources (white, red, blue))
- iii.
- Temperature
- iv.
- pH
- v.
- Salinity
- vi.
- Size of inoculation
4.1. Salinity
4.2. Light Irradiation
4.3. Temperature
4.4. Nutrients
4.5. Metals
4.6. Chemicals
4.7. Combination of Abiotic Factors on Antioxidant Enzyme Activity
5. Culturing Systems
6. Post-Culture Downstream Processing
6.1. Harvesting
6.2. Post-Harvesting Storage Processing
7. Challenges and Future Opportunities
- i.
- ii.
- Transferring lab-scale optimised abiotic stress strategies to a commercial scale;
- iii.
- Seasonal and environmental variation in biomass production, and the control of competitors in open ponds cultivation systems;
- iv.
- Engineering developments of sustainable large-scale technologies (open raceways ponds and PBR) for Dunaliella and microalgae in general for efficient biomass and enzymatic antioxidant production;
- v.
- Developing sustainable downstream technologies (harvesting, extraction, and purification) for antioxidant enzymes production at commercial scale;
- vi.
- Establishing tools for proteomics, genomics, and metabolomics, and implement them to develop strains for the accumulation of intracellular antioxidant enzymes;
- vii.
- Stabilisation of antioxidant enzymes in food ingredients and extension of their shelf life;
- viii.
- Algae extracts can be used as taste enhances in foods; however, some strains are associated with a bitter or astringent taste;
- ix.
- Introducing a general platform to solve regulatory and labelling issues, as this varies from country to country.
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hadi, M.R.; Shariati, M.; Afsharzadeh, S. Microalgal biotechnology: Carotenoid and glycerol production by the green algae Dunaliella isolated from the Gave-Khooni salt marsh, Iran. Biotechnol. Bioprocess Eng. 2008, 13, 540–544. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, R.A.; He, M.; Aftab, R.A.; Zheng, S.; Nagi, M.; Bakri, R.; Wang, C. Bioenergy application of Dunaliella salina SA 134 grown at various salinity levels for lipid production. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gujar, A.; Cui, H.; Ji, C.; Kubar, S.; Li, R. Development, Production and Market Value of Microalgae Products. Appl. Microbiol. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; De Cooman, L. Microalgae as a Novel Source of Antioxidants for Nutritional Applications. Handb. Mar. Microalgae Biotechnol. Adv. 2015, 10, 160–180. [Google Scholar] [CrossRef]
- Marino, T.; Casella, P.; Sangiorgio, P.; Verardi, A.; Ferraro, A.; Hristoforou, E.; Molino, A.; Musmarra, D. Natural beta-carotene: A microalgae derivate for nutraceutical applications. Chem. Eng. Trans. 2020, 79, 103–108. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs. 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Raja, R.; Coelho, A.; Hemaiswarya, S.; Kumar, P.; Carvalho, I.S.; Alagarsamy, A. Applications of microalgal paste and powder as food and feed: An update using text mining tool. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 740–747. [Google Scholar] [CrossRef]
- Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.N.; Gayen, K.; Bhowmick, T.K. Downstream processing of microalgae for pigments, protein and carbohydrate in industrial application: A review. Food Bioprod. Process. 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Figueira, F.D.S.; Crizel, T.D.M.; Silva, C.R.; Salas-Mellado, M.d.l.M. Elaboration of gluten-free bread enriched with the microalgae Spirulina platensis. Braz. J. Food Technol. 2011, 14, 308–316. [Google Scholar]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella vulgaris and Spirulina maxima biomass in pasta products. Part 1: Preparation and evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Nicolau, E.; de Oliveira, R.G.; Fuentes-Grünewald, C.; Picot, L. Analysis of Scientific Research Driving Microalgae Market Opportunities in Europe. Mar. Drugs 2020, 18, 264. [Google Scholar] [CrossRef] [PubMed]
- Transparency Market Research. Microalgae—Based Products Market (Product: Spirulina, Chlorella, Astaxanthin, β-Carotene, PUFA Fatty Acid, Lutein, and Others; Food and Beverages, Personal Care, Nutraceuticals and Dietary Supplements, Pharmaceuticals, Chemicals, Fuel, Animal Feed)—Global Industry Analysis, Sixe, Share, Growth, Trends, and Forecast, 2020–2030. 2020. Available online: https://www.transparencymarketresearch.com/about-us.html (accessed on 26 April 2021).
- Maadane, A.; Merghoub, N.; Ainane, T.; El Arroussi, H.; Benhima, R.; Amzazi, S.; Bakri, Y.; Wahby, I. Antioxidant activity of some Moroccan marine microalgae: Pufa profiles, carotenoids and phenolic content. J. Biotechnol. 2015, 215, 13–19. [Google Scholar] [CrossRef]
- Gangadhar, K.N.; Pereira, H.; Rodrigues, M.J.; Custódio, L.; Barreira, L.; Malcata, F.X.; Varela, J. Microalgae-based unsaponifiable matter as source of natural antioxidants and metal chelators to enhance the value of wet Tetraselmis chuii biomass. Open Chem. 2016, 14, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Assuncao, M.F.G.; Amaral, R.; Martins, C.B.; Ferreira, J.D.; Ressurreicao, S.; Santos, S.D.; Varejão, J.M.T.B.; Santos, L.M.A. Screening microalgae as potential sources of antioxidants. J. Appl. Phycol. 2017, 29, 865–877. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangl, D.; Zedler, J.A.Z.; Rajakumar, P.D.; Martinez, E.M.R.; Riseley, A.; Włodarczyk, A.; Purton, S.; Sakuragi, Y.; Howe, C.J.; Jensen, P.E.; et al. Biotechnological exploitation of microalgae. J. Exp. Bot. 2015, 66, 6975–6990. [Google Scholar] [CrossRef] [Green Version]
- Villarruel-López, A.; Ascencio, F.; Nuño, K. Microalgae, a Potential Natural Functional Food Source—A Review. Polish J. Food Nutr. Sci. 2017, 67, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jiang, J.G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell. Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. The biotechnology of cultivating the halotolerant alga Dunaliella. Trends Biotechnol. 1990, 8, 121–126. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Shariati, M.; Hadi, M.R. Microalgal Biotechnology and Bioenergy in Dunaliella. In Progress in Molecular and Environmental Bioengineering—From Analysis and Modeling to Technology Applications; Carpi, A., Ed.; Intechopen: Rijeka, Croatia, 2011; Available online: https://www.intechopen.com/books/progress-in-molecular-and-environmental-bioengineering-from-analysis-and-modeling-to-technology-applications/microalgal-biotechnology-and-bioenergy-in-dunaliella (accessed on 26 April 2021). [CrossRef] [Green Version]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal industry in China: Challenges and prospects. J. Appl. Phycol. 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Sui, Y.; Vlaeminck, S.E. Dunaliella Microalgae for Nutritional Protein: An Undervalued Asset. Trends Biotechnol. 2020, 38, 10–12. [Google Scholar] [CrossRef]
- Liang, M.H.; Xue, L.L.; Jiang, J.G. Two-stage cultivation of Dunaliella tertiolecta with glycerol and triethylamine for lipid accumulation: A viable way to alleviate the inhibitory effect of triethylamine on biomass. Appl. Environ. Microbiol. 2019, 85, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalamurugan, G.L.; Valerie, O.; Mark, L. Valuable bioproducts obtained from microalgal biomass and their commercial applications: A review. Environ. Eng. Res. 2018, 23, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Church, J.; Hwang, J.H.; Kim, K.T.; McLean, R.; Oh, Y.K.; Nam, B.; Joo, J.C.; Lee, W.H. Effect of salt type and concentration on the growth and lipid content of Chlorella vulgaris in synthetic saline wastewater for biofuel production. Bioresour. Technol. 2017, 243, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Widowati, I.; Zainuri, M.; Kusumaningrum, H.P.; Susilowati, R.; Hardivillier, Y.; Leignel, V.; Bourgougnon, N.; Mouget, J.L. Antioxidant activity of three microalgae Dunaliella salina, Tetraselmis chuii and Isochrysis galbana clone Tahiti. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bali, Indonesia, 2017; Volume 55, p. 012067. [Google Scholar]
- Singh, R.; Kumar, M.; Mittal, A.; Kumar, P. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Raveendran, S.; Kuruvilla, A.; Rebello, S. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef]
- Hanaa, H.; El Baz, F.K.; El-Baroty, G.S. Production of Antioxidant by the Green Alga Dunaliella salina. Int. J. Agric. Biol. 2004, 6, 49–57. [Google Scholar]
- Haghjou, M.M.; Shariati, M.; Smirnoff, N. The effect of acute high light and low temperature stresses on the ascorbate-glutathione cycle and superoxide dismutase activity in two Dunaliella salina strains. Physiol. Plant. 2009, 135, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Moane, S.; Murray, P. Effect of macro- and micro-nutrient limitation on superoxide dismutase activities and carotenoid levels in microalga Dunaliella salina CCAP 19/18. Bioresour. Technol. 2013, 147, 23–28. [Google Scholar] [CrossRef]
- Al-Rashed, S.A.; Ibrahim, M.M.; El-Gaaly, G.A.; Al-Shehri, S.; Mostafa, A. Evaluation of radical scavenging system in two microalgae in response to interactive stresses of UV-B radiation and nitrogen starvation. Saudi J. Biol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Baranwal, M.; Reddy, S.M. Antioxidant and cytotoxic activity of carotenes produced by Dunaliella salina under stress. Pharm. Biol. 2016, 54, 2269–2275. [Google Scholar] [CrossRef] [Green Version]
- Mirshekari, M.; Einali, A.; Valizadeh, J. Metabolic changes and activity pattern of antioxidant enzymes induced by salicylic acid treatment in green microalga Dunaliella salina under nitrogen deficiency. J. Appl. Phycol. 2019, 31, 1709–1719. [Google Scholar] [CrossRef]
- Hosseini Tafreshi, A.; Shariati, M. Dunaliella biotechnology: Methods and applications. J. Appl. Microbiol. 2009, 107, 14–35. [Google Scholar] [CrossRef]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar]
- Krishnamurthy, P.; Wadhwani, A. Antioxidant Enzymes and Human Health. In Antioxidant Enzyme; Intech: Rijeka, Croatia, 2012; Volume 1, pp. 3–18. Available online: https://www.intechopen.com/books/antioxidant-enzyme/antioxidant-enzymes-and-human-health (accessed on 26 April 2021). [CrossRef] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K. Assay Guided Comparison for Enzymatic and Non-Enzymatic Antioxidant Activities with Special Reference to Medicinal Plants. In Antioxidant Enzyme; Intech: Rijeka, Croatia, 2012; Volume 14, pp. 381–400. Available online: https://www.intechopen.com/books/antioxidant-enzyme/assay-guided-comparison-for-enzymatic-and-non-enzymatic-antioxidant-activities-with-special-referenc (accessed on 26 April 2021). [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mtaki, K.; Kyewalyanga, M.S.; Mtolera, M.S.P. Assessment of antioxidant contents and free radical-scavenging capacity of chlorella vulgaris cultivated in low cost media. Appl. Sci. 2020, 10, 8611. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, J.; Mehandia, S.; Singh, G.; Raina, A.; Arya, S.K. Catalase enzyme: Application in bioremediation and food industry. Biocatal. Agric. Biotechnol. 2018, 16, 192–199. [Google Scholar] [CrossRef]
- Gopal, R.K.; Elumalai, S. Industrial Production of Superoxide Dismutase (SOD): A Mini Review. J. Probiotics Health 2017, 5. [Google Scholar] [CrossRef]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A Comprehensive Review on Function and Application of Plant Peroxidases. Biochem. Anal. Biochem. 2017, 6, 1–16. [Google Scholar] [CrossRef]
- Keokamnerd, T.; Acton, J.C.; Han, I.Y.; Dawson, P.L. Effect of commercial rosemary oleoresin preparations on ground chicken thigh meat quality packaged in a high-oxygen atmosphere. Poult. Sci. 2008, 87, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M. Khalil-ur-Rehman Potential applications of peroxidases. Food Chem. 2009, 115, 1177–1186. [Google Scholar] [CrossRef]
- Canter, N. Use of antioxidants in automotive lubricants. Tribol. Lubr. Technol. 2008, 64, 12. [Google Scholar]
- Jegannathan, K.R.; Nielsen, P.H. Environmental assessment of enzyme use in industrial production-a literature review. J. Clean. Prod. 2013, 42, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Godic, A.; Poljšak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell. Longev. 2014, 2014, 860479. [Google Scholar] [CrossRef]
- Allemann, I.B.; Baumann, L. Antioxidants used in skin care formulations. Ski. Ther. Lett 2008, 13, 5–9. [Google Scholar]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Bafana, A.; Dutt, S.; Kumar, S.; Ahuja, P.S. Superoxide dismutase: An industrial perspective. Crit. Rev. Biotechnol. 2011, 31, 65–76. [Google Scholar] [CrossRef]
- Beach, D. Antioxidants (Natural, and Synthetic) Market for Pharmaceuticals, Food & Beverages Sector, Feed Additives, Cosmetics Industry, and Other Applications: Global Industry Perspective, Comprehensive Analysis, and Forecast, 2014–2020, Marrket Research Store. 2016. Available online: http://www.marketresearchstore.com/report/antioxidants-market-for-pharmaceuticals-food-z38119 (accessed on 26 April 2021).
- Sasse, A.; Colindres, P.; Brewer, M.S. Effect of natural and synthetic antioxidants on the oxidative stability of cooked, frozen pork patties. J. Food Sci. 2009, 74, 1–6. [Google Scholar] [CrossRef]
- Schillaci, C.; Nepravishta, R.; Bellomaria, A. Antioxidants in food and pharmaceutical research. Albanian J. Pharm. Sci. 2014, 1, 15–25. [Google Scholar]
- Ito, N.; Hirose, M.; Fukushima, S.; Tsuda, H.; Shirai, T.; Tatematsu, M. Studies on antioxidants: Their carcinogenic and modifying effects on chemical carcinogenesis. Food Chem. Toxicol. 1986, 24, 1071–1082. [Google Scholar] [CrossRef]
- Thompson, D.C.; Trush, M.A. Studies on the mechanism of enhancement of butylated hydroxytoluene-induced mouse lung toxicity by butylated hydroxyanisole. Toxicol. Appl. Pharmacol. 1988, 96, 122–131. [Google Scholar] [CrossRef]
- Silaghi-dumitrescu, L.; Loghin, F. Evaluation of the possible endocrine disruptive effect of butylated hydroxyanisole, butylated hydroxytoluene and propyl gallate in immature female rats. Farmacia 2013, 61, 202–211. [Google Scholar]
- Thorat, I.D.; Jagtap, D.D.; Mohapatra, D.; Joshi, D.C.; Sutar, R.F.; Kapdi, S.S. Antioxidants, their properties, uses in food products and their legal implications. Int. J. Food Stud. 2013, 2, 81–104. [Google Scholar] [CrossRef]
- El Shafey, H.M.; Bahashwan, S.A.; Alghaithy, A.A.; Ghanem, S. Microbial superoxide dismutase enzyme as therapeutic agent and future gene therapy. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 435–443. [Google Scholar]
- Surai, P.; Surai, P.F. Antioxidant Systems in Poultry Biology: Superoxide Dismutase. Anim. Nutr. 2015, 1, 1–17. [Google Scholar] [CrossRef]
- Gridley, D.S.; Green, L.M.; Nelson, G.A.; Pecaut, M.J.; Slater, J.M. Therapeutic Utilities of SOD Mimetics: Cancer, Radiotherapy and SOD Mimetics. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK6099/ (accessed on 26 April 2021).
- Miłek, J.; Wójcik, M.; Verschelde, W. Thermal stability for the effective use of commercial catalase. Polish J. Chem. Technol. 2014, 16, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Morón, Ú.M.; Castilla-Cortázar, I. Protection Against Oxidative Stress and “IGF-I Deficiency Conditions”. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Available online: https://www.intechopen.com/books/antioxidant-enzyme/protection-against-oxidative-stress-and-igf-i-deficiency-conditions- (accessed on 26 April 2021). [CrossRef] [Green Version]
- Chenevix Trench, G.; Young, J.; Coggan, M.; Board, P. Glutathione S-transferase M1 and T1 polymorphisms: Susceptibility to colon cancer and age of onset. Carcinogenesis 1995, 16, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, E.; Chan, G.P. Skin-whitening and skin-condition-improving effects of topical oxidized glutathione: A double- blind and placebo-controlled clinical trial in healthy women. Clin. Cosmet. Investig. Dermatol. 2014, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Barrita, J.; Sánchez, M. Antioxidant Role of Ascorbic Acid and His Protective Effects on Chronic Diseases. Oxidative Stress Chronic Degener. Dis. A Role Antioxid. 2013, 450–484. [Google Scholar] [CrossRef] [Green Version]
- Borek, C. Antioxidants and the prevention of hormonally regulated cancer. J. Men’s Health Gend. 2005, 2, 346–352. [Google Scholar] [CrossRef]
- Constantinou, C.; Papas, A.; Constantinou, A.I. Vitamin E and cancer: An insight into the anticancer activities of vitamin E isomers and analogs. Int. J. Cancer 2008, 123, 739–752. [Google Scholar] [CrossRef]
- Petrulea, M.; Muresan, A.; Duncea, I. Oxidative Stress and Antioxidant Status in Hypo- and Hyperthyroidism. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Available online: https://www.intechopen.com/books/antioxidant-enzyme/oxidative-stress-and-antioxidant-status-in-hypo-and-hyperthyroidism (accessed on 26 April 2021). [CrossRef] [Green Version]
- Kaliora, A.C.; Dedoussis, G.V.Z.; Schmidt, H. Dietary antioxidants in preventing atherogenesis. Atherosclerosis 2006, 187, 1–17. [Google Scholar] [CrossRef]
- Sak, K. Cytotoxicity of dietary flavonoids on different human cancer types. Pharmacogn. Rev. 2014, 8, 122–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Wei, H.; He, B. Dietary flavonoids intake and the risk of coronary heart disease: A dose-response meta-analysis of 15 prospective studies. Thromb. Res. 2015, 135, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.L.; Feskens, E.J.M.; Kromhout, D.; Hertog, M.G.L.; Hollman, P.C.H.; Hertog, M.G.L.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. 2014, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.A.; Custódio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, W.S.; Mozaffarian, D.; Rimm, E.; Kris-Etherton, P.; Rudel, L.L.; Appel, L.J.; Engler, M.M.; Engler, M.B.; Sacks, F. Omega-6 fatty acids and risk for cardiovascular disease: A science advisory from the American Heart Association nutrition subcommittee of the council on nutrition, physical activity, and metabolism; council on cardiovascular nursing; and council on epidem. Circulation 2009, 119, 902–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundebye, A.-K.; Hove, H.; Måge, A.; Bohne, V.J.B.; Hamre, K. Levels of synthetic antioxidants (ethoxyquin, butylated hydroxytoluene and butylated hydroxyanisole) in fish feed and commercially farmed fish. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2010, 27, 1652–1657. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Olszowy, M.; Jóźwik-Dolęba, M. Antagonistic Antioxidant Effect in Butylated Hydroxytoluene/Butylated Hydroxyanisole Mixture. J. Food Process. Preserv. 2015, 39, 2240–2248. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef]
- Kaushik, D.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Bak, S.; Beisson, F.; Bishop, G.; Hamberger, B.; Höfer, R.; Paquette, S.; Werck-Reichhart, D. Cytochromes P450. Arab. B 2011, 9, e0144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Montalvo, A.; Sun, Y.; Diaz-Ruiz, A.; Ali, A.; Gutierrez, V.; Palacios, H.H.; Curtis, J.; Siendones, E.; Ariza, J.; Abulwerdi, G.A. Cytochrome b5 reductase and the control of lipid metabolism and healthspan. NPJ Aging Mech. Dis. 2016, 2, 16006. [Google Scholar] [CrossRef] [PubMed]
- Pruchniak, M.P.; Araźna, M.; Demkc, U. Biochemistry of oxidative stress. Adv. Exp. Med. Biol. 2016, 878, 9–19. [Google Scholar] [CrossRef]
- Barbusiński, K. Controversy Over Fenton Mechanism. Ecol. Chem. Eng. S 2009, 16, 347–358. [Google Scholar]
- Tatsumi, H.; Nakase, H.; Kano, K.; Ikeda, T. Mechanistic study of the autoxidation of reduced flavin and quinone compounds. J. Electroanal. Chem. 1998, 443, 236–242. [Google Scholar] [CrossRef]
- Vašková, J.; Vaško, L.; Kron, I. Oxidative Processes and Antioxidative Metaloenzymes. In Antioxidant Enzyme; Intechopen: Rijeka, Croatia, 2012; Volume 2, pp. 19–58. Available online: https://www.intechopen.com/books/antioxidant-enzyme/oxidative-processes-and-antioxidative-metaloenzymes (accessed on 26 April 2021). [CrossRef] [Green Version]
- Gupta, V.K.; Sharma, S.K. Plants as Natural Antioxidants. Indian J. Nat. Prod. Resour. 2006, 5, 326–334. [Google Scholar]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Marí, M.; Morales, A.; Colell, A.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial glutathione, a key survival antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2700. [Google Scholar] [CrossRef] [Green Version]
- Jajic, I.; Sarna, T.; Strzalka, K. Senescence, Stress, and Reactive Oxygen Species. Plants 2015, 4, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: A review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef] [Green Version]
- Ben-Amotz, A.; Polle, J.E.W.; Rao, D.V.S. The Alga Dunaliella; CRC Press: Boca Raton, FL, USA, 2009; ISBN 1578085454. [Google Scholar]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Valdes, J.R.; Aguilar, C.; Contreras-Esquivel, J.C.; Méndez-Zavala, A.; Montañez, J. Strategies to enhance the production of photosynthetic pigments and lipids in chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.L.; Lee, S.M.; Choi, H.J. Integration of microalgal cultivation system for wastewater remediation and sustainable biomass production. World J. Microbiol. Biotechnol. 2016, 32. [Google Scholar] [CrossRef] [PubMed]
- Kharati-Koupaei, M.; Moradshasi, A. Effects of sodium nitrate and mixotrophic culture on biomass and lipid production in hypersaline microalgae Dunaliella viridis Teod. Braz. Arch. Biol. Technol. 2016, 59, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Yeh, K.L.; Aisyah, R.; Lee, D.J.; Chang, J.S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Gibor, A. The culture of brine algae. Biol. Bull. 1956, 111, 223–229. [Google Scholar] [CrossRef]
- Gladue, R.M.; Maxey, J.E. Microalgal feeds for aquaculture. J. Appl. Phycol. 1994, 6, 131–141. [Google Scholar] [CrossRef]
- Ramos, A.A.; Polle, J.; Tran, D.; Cushman, J.C.; Jin, E.; Varela, J.C. The unicellular green alga Dunaliella salina Teod. as a model for abiotic stress tolerance: Genetic advances and future perspectives. Algae 2011, 26, 3–20. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Borowitzka, L.J. Micro-Algal Biotechnology; Cambridge University Press: Cambridge, UK, 1988; ISBN 0521323495. [Google Scholar]
- Herrero, C.; Cid, A.; Fabregas, J.; Abalde, J. Yields in biomass and chemical constituents of four commercially important marine microalgae with different culture media. Aquac. Eng. 1991, 10, 99–110. [Google Scholar] [CrossRef]
- Cifuentes, A.S.; Gonzalez, M.A.; Parra, O.O. The effect of salinity on the growth and carotenogenesis in two Chilean strains of Dunaliella salina Teodoresco. Biol. Res. 1996, 29, 227–236. [Google Scholar] [PubMed]
- Ibañez, E.; Cifuentes, A. Benefits of using algae as natural sources of functional ingredients. J. Sci. Food Agric. 2013, 93, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Tredici, M.R.; Rodolfi, L.; Biondi, N.; Bassi, N.; Sampietro, G. Techno-economic analysis of microalgal biomass production in a 1-ha Green Wall Panel (GWP®) plant. Algal Res. 2016, 19, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Singh, B. Algal biorefinery: An integrated approach for sustainable biodiesel production. Biomass Bioenergy 2019, 131, 105398. [Google Scholar] [CrossRef]
- Harvey, P.J.; Ben-Amotz, A. Towards a sustainable Dunaliella salina microalgal biorefinery for 9-cis β-carotene production. Algal Res. 2020, 50, 102002. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Paglia, G.; Magnúsdóttir, M.; Steinarsdóttir, E.A.; Gudmundsson, S.; Palsson, B.Ø.; Andrésson, Ó.S.; Brynjólfsson, S. Effects of abiotic stressors on lutein production in the green microalga Dunaliella salina. Microb. Cell Fact. 2014, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Jha, B. Antioxidant response of the microalga Dunaliella salina under salt stress. Bot. Mar. 2011, 54, 195–199. [Google Scholar] [CrossRef]
- Mishra, A.; Mandoli, A.; Jha, B. Physiological characterization and stress-induced metabolic responses of Dunaliella salina isolated from salt pan. J. Ind. Microbiol. Biotechnol. 2008, 35, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Tammam, A.A.; Fakhry, E.M.; El-sheekh, M. Effect of salt stress on antioxidant system and the metabolism of the reactive oxygen species in Dunaliella salina and Dunaliella tertiolecta. J. Biotechnol. 2011, 10, 3795–3808. [Google Scholar] [CrossRef]
- Jahnke, L.S.; White, A.L. Long-term hyposaline and hypersaline stresses produce distinct antioxidant responses in the marine alga Dunaliella tertiolecta. J. Plant Physiol. 2003, 160, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Tian, J.; Yu, J. Changes in ultrastructure and responses of antioxidant systems of algae (Dunaliella salina) during acclimation to enhanced ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2009, 97, 152–160. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, acumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [Green Version]
- Janknegt, P.J.; De Graaff, C.M.; Van De Poll, W.H.; Visser, R.J.W.; Helbling, E.W.; Buma, A.G.J. Antioxidative responses of two marine microalgae during acclimation to static and fluctuating natural uv radiation. Photochem. Photobiol. 2009, 85, 1336–1345. [Google Scholar] [CrossRef]
- White, A.L.; Jahnke, L.S. Contrasting effects of UV-A and UV-B on photosynthesis and photoprotection of beta-carotene in two Dunaliella spp. Plant Cell Physiol. 2002, 43, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Polle, J.E.W.; Melis, A.; Lee, T.K.; Jin, E. Up-regulation of photoprotection and PSII-repair gene expression by irradiance in the unicellular green alga Dunaliella salina. Mar. Biotechnol. 2006, 8, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Van De Laak, C.C.W.; Kaasenbrood, P.S.; Lorier, J.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in light-stressed Dunaliella salina. Biotechnol. Bioeng. 2010, 106, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Ras, M.; Steyer, J.P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 2013, 12, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Haghjou, M.M.; Shariati, M.; Pozveh, M.H. The effect of low light intensities on oxidative stress induced by short-term chilling in Dunaliella salina teod. Pak. J. Biol. Sci. 2006, 9, 2048–2054. [Google Scholar] [CrossRef]
- Alizadeh, G.I.; Aliev, I.I.; Magerramova, K.K.; Galandarli, I.Z.; Dibirova, G.H.; Jalilova, A.R. The response reaction of Dunaliella cells against the influence of Methylene blue and Norflurazon under the low temperature stress conditions. Eur. J. Biotechnol. Biosci. 2015, 3, 7–10. [Google Scholar]
- Lv, H.; Cui, X.; Wahid, F.; Xia, F.; Zhong, C.; Jia, S. Analysis of the physiological and molecular responses of Dunaliella salina to macronutrient deprivation. PLoS ONE 2016, 11, e0152226. [Google Scholar] [CrossRef] [PubMed]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative Stress Is a Mediator for Increased Lipid Accumulation in a Newly Isolated Dunaliella salina Strain. PLoS ONE 2014, 9, e91957. [Google Scholar] [CrossRef]
- Michalak, a Phenolic Compounds and Their Antioxidant Activity in Plants Growing under Heavy Metal Stress. Plant Cell 2006, 15, 523–530.
- Hazani, A.A.; Ibrahim, M.M.; Shehata, A.I.; El-Gaaly, G.A.; Daoud, M.; Fouad, D.; Rizwana, H.; Moubayed, N. Ecotoxicity of Ag-nanoparticles on two microalgae, Chlorella vulgaris and Dunaliella tertiolecta. Arch. Biol. Sci. 2013, 65, 1447–1457. [Google Scholar] [CrossRef]
- Zamani, N.; Rasekh, F.; Moradshahi, A.; Kholdebarin, B. Physiological responses of Dunaliella tertiolecata to Hg2+-induced oxidative stress. Iran. J. Sci. Technol. 2009, 33, 65–74. [Google Scholar]
- Tsuji, N.; Hirayanagi, N.; Okada, M.; Miyasaka, H.; Hirata, K.; Zenk, M.H.; Miyamoto, K. Enhancement of tolerance to heavy metals and oxidative stress in Dunaliella tertiolecta by Zn-induced phytochelatin synthesis. Biochem. Biophys. Res. Commun. 2002, 293, 653–659. [Google Scholar] [CrossRef]
- Nikookar, K.; Moradshahi, A.; Hosseini, L. Physiological responses of Dunaliella salina and Dunaliella tertiolecta to copper toxicity. Biomol. Eng. 2005, 22, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, L.; Zhang, W. Chemicals to enhance microalgal growth and accumulation of high-value bioproducts. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Qv, X.Y.; Jiang, J.G. Toxicity evaluation of two typical surfactants to Dunaliella bardawil, an environmentally tolerant alga. Environ. Toxicol. Chem. 2013, 32, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Mofeed, J.; Abdel-aal, E.I. Effect of Phenol on Some Antioxidant Enzymes in the Marine Microalga Dunaliella salina. J. Environ. Sci. (JOESE 5) 2015, 44, 185–196. [Google Scholar]
- Tappel, A.L. Glutathione peroxidase and hydroperoxides. Methods Enzymol. 1978, 52, 506–513. [Google Scholar]
- Chen, H.; Jiang, J.G. Toxic effects of chemical pesticides (trichlorfon and dimehypo) on Dunaliella salina. Chemosphere 2011, 84, 664–670. [Google Scholar] [CrossRef]
- Einali, A.; Valizadeh, J. Propyl gallate promotes salt stress tolerance in green microalga Dunaliella salina by reducing free radical oxidants and enhancing β-carotene production. Acta Physiol. Plant. 2015, 37, 1–11. [Google Scholar] [CrossRef]
- Nikookar, K.; Moradshahi, A.; Kharati, M. Influence of Salinity on the Growth, Pigmentation and Ascorbate Peroxidase Activity of Dunaliella Lake in Shiraz. Iran. J. Sci. Technol. 2004, 28, 117–125. [Google Scholar]
- Marcano, L.B.C.; Carruyo, I.M.; Montiel, X.M.; Morales, C.B.; De Soto, P.M. Effect of cadmium on cellular viability in two species of microalgae (Scenedesmus sp. and Dunaliella viridis). Biol. Trace Elem. Res. 2009, 130, 86–93. [Google Scholar] [CrossRef]
- Arun, N.; Vidyalaxmi, V.; Singh, D.P. Chromium (VI) induced oxidative stress in halotolerant alga Dunaliella salina and D. tertiolecta isolated from sambhar salt lake of Rajasthan (India). Cell. Mol. Biol. 2014, 60, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Tsiaka, P.; Tsarpali, V.; Ntaikou, I.; Kostopoulou, M.N.; Lyberatos, G.; Dailianis, S. Carbamazepine-mediated pro-oxidant effects on the unicellular marine algal species Dunaliella tertiolecta and the hemocytes of mussel Mytilus galloprovincialis. Ecotoxicology 2013, 22, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal carotenoids: A review of production, current markets, regulations, and future direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, E.; Acién, F.G.; Fernández, J.M.; García, J.L.; González, C.; Molina, E. Characterization of a flat plate photobioreactor for the production of microalgae. Chem. Eng. J. 2008, 138, 136–147. [Google Scholar] [CrossRef]
- García-González, M.; Moreno, J.; Manzano, J.C.; Florencio, F.J.; Guerrero, M.G. Production of Dunaliella salina biomass rich in 9-cis-??-carotene and lutein in a closed tubular photobioreactor. J. Biotechnol. 2005, 115, 81–90. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Jiang, J.G. Continuous cultivation of Dunaliella salina in photobioreactor for the production of β-carotene. Eur. Food Res. Technol. 2008, 227, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Kleinegris, D.M.M.; Janssen, M.; Brandenburg, W.A.; Wijffels, R.H. Continuous production of carotenoids from Dunaliella salina. Enzym. Microb. Technol. 2011, 48, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Ndukwe, G.I.; Mustapha, K.B.; Ayo, R.I. Constraints to large scale algae biomass production and utilization. J. Algal Biomass Util. 2012, 3, 14–32. [Google Scholar]
- Kim, J.; Yoo, G.; Lee, H.; Lim, J.; Kim, K.; Kim, C.W.; Park, M.S.; Yang, J.W. Methods of downstream processing for the production of biodiesel from microalgae. Biotechnol. Adv. 2013, 31, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J.; Heaven, S. A review of the harvesting of micro-algae for biofuel production. Rev. Environ. Sci. Biotechnol. 2013, 12, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.K.; Garg, S.; Li, Y.; Malekizadeh, A.; Schenk, P.M. Critical analysis of current Microalgae dewatering techniques. Biofuels 2013, 4, 397–407. [Google Scholar] [CrossRef]
- Japar, A.S.; Takriff, M.S.; Yasin, N.H.M. Harvesting microalgal biomass and lipid extraction for potential biofuel production: A review. J. Environ. Chem. Eng. 2017, 5, 555–563. [Google Scholar] [CrossRef]
- Roy, M.; Mohanty, K. A comprehensive review on microalgal harvesting strategies: Current status and future prospects. Algal Res. 2019, 44, 101683. [Google Scholar] [CrossRef]
- Najjar, Y.S.; Abu-Shamleh, A. Harvesting of microalgae by centrifugation for biodiesel production: A review. Algal Res. 2020, 51, 102046. [Google Scholar] [CrossRef]
- Gao, S.; Yan, R.; Chen, F. Effects of Different Storage Temperatures and Times on Germination and Antioxidant Responses of Jatropha curcas L. Seeds. J. Agric. Sci. Technol. 2015, 17, 1619–1628. [Google Scholar]
- Ahmed, F.; Li, Y.; Fanning, K.; Netzel, M.; Schenk, P.M. Effect of drying, storage temperature and air exposure on astaxanthin stability from Haematococcus pluvialis. Food Res. Int. 2015, 74, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Trung, T.S.; Huyen, N.T.K.; Minh, N.C.; Le Trang, T.T.; Han, N.T. Optimization of harvesting of microalgal Thalassiosira pseudonana biomass using chitosan prepared from shrimp shell waste. Asian J. Agric. Res. 2016, 10, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.S.; Rufino, M.S.; Moura, C.F.; Cavalcanti, F.R.; Alves, R.E.; Miranda, M.R. The influence of processing and long-term storage on the antioxidant metabolism of acerola (Malpighia emarginata) purée. Rev. Bras. Frutic. 2011, 30, 398–408. [Google Scholar] [CrossRef]
- Mediani, A.; Abas, F.; Tan, C.; Khatib, A. Effects of Different Drying Methods and Storage Time on Free Radical Scavenging Activity and Total Phenolic Content of Cosmos Caudatus. Antioxidants 2014, 3, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Roy, U.K.; Nielsen, B.V.; Milledge, J.J. Effect of post-harvest conditions on antioxidant enzyme activity in Dunaliella tertiolecta biomass. Biocatal. Agric. Biotechnol. 2020, 27, 101661. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Wosu, C.I.; Ben-Amotz, A.; Harvey, P.J. Potential of new isolates of Dunaliella salina for natural β-carotene production. Biology 2018, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.R.; Rao, P.H.; Subramanian, V.V.; Sivasubramanian, V. Enzymatic and non-enzymatic antioxidant potentials of Chlorella vulgaris grown in effluent of a confectionery industry. J. Food Sci. Technol. 2014, 51, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miȩkus, N.; Iqbal, A.; Marszałek, K.; Puchalski, C.; Świergiel, A. Green chemistry extractions of carotenoids from daucus carota L.-Supercritical carbon dioxide and enzyme-assisted methods. Molecules 2019, 24, 4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavka, A.; Jornsson, L.J. Comparison of the growth of filamentous fungi and yeasts in lignocellulose-derived media. Biocatal. Agric. Biotechnol. 2014, 3, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Gomez, J.A.; Höffner, K.; Barton, P.I. From sugars to biodiesel using microalgae and yeast. Green Chem. 2016, 18, 461–475. [Google Scholar] [CrossRef] [Green Version]
- Munir, N.; Sharif, N.; Naz, S.; Farkhanda, M. Algae: A potent antioxidant source. Sky J. Microbiol. Res. 2013, 1, 22–31. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Antioxidants | Applications | Ref | |
|---|---|---|---|
| Natural antioxidants | SOD | Added to cosmetic products to protect against skin damage | [49,52,60,63,64] |
| Protect against lipid peroxidation, heat, and cold stress in poultry production | |||
| As a therapeutic agent for treatment of inflammatory disorders | |||
| Normal cells protector during radiotherapy for cancer patients | |||
| CAT | Eliminate excessive H2O2 in the textile industry, pulp, and paper industry used for bleaching fibres and pulp, and as a bactericidal disinfectant in food processing and in the pharmaceutical industry | [51,57,65] | |
| In aesthetics (mask treatment) to increase cellular oxygenation in the upper layers of facial epidermis | |||
| Reducing the risk of diabetes mellitus | |||
| GPX | Immune system booster | [42,66] | |
| GST | Protective role against neurogenerative diseases | [66,67] | |
| Decreases the risk of tumours of the head and neck, oral, cavity and colon | |||
| Glutathione | Anti-wrinkle formation, and as a modifier of skin smoothness | [68] | |
| Vitamins | As a food preservative and bread improver, protective activity against heart diseases, reduced the risk of colorectal adenomas and prostate cancer, reduction of thyroid hormone levels | [69,70,71,72,73] | |
| Flavonoids | As cancer preventive agents, protection against type 2 diabetes Functional food additive | [74,75,76,77] | |
| Carotenoid | Anticancer agents, additive to cosmetics and multivitamin preparation Food colouring agent, pro-vitamin A in food and animal feed | [29,38,78] | |
| PUFA | Prevention of heart and inflammatory diseases | [79,80] | |
| Synthetic Antioxidants | BHA | Extending the shelf life of vegetable oil, frying oil, animal feed, cereals, chewing gum, potato flakes and cosmetic products | [81,82,83] |
| BHT | Increasing the shelf life of animal fats, chewing gum, animal feed, vegetable oils | [82,83] | |
| TBHQ | Used as preservative for enhancing storage stability of vegetable oils, margarine, fish oil, fried foods, essential oils, nuts, edible animal fats, butterfat, and packed fried foods | [82] | |
| Propyl gallate | As an antioxidant agent in foods and vegetable oil | [82] | |
| BHA = butylated hydroxyl anisole; TBHQ = tert-butylhydroquinone; BHT = butylated hydroxyl toluene | |||
| ROS | Reaction | Life Span | Function | Sources of ROS | Scavenging Antioxidants |
|---|---|---|---|---|---|
| Singlet (1O2) First excited electronic state of O2 | Chlorophyll triplet state (Chl) is generated sue to insufficient energy dissipation during photosynthesis Chl + 3O2 → 1O2 Reduction of transition metal (Fe3+) O2•− + Fe3+ → 1O2 + Fe2+ | 3 µs (appx.) 4 µs in H2O 100 µs in polar solvents | Gene up-regulation, molecular defense against photo-oxidative stress | Chloroplast | β-carotene, lycopene, tocopherol, ASc, plastoquinone, and proline |
| Superoxide radical (O2•−) One electron reduction of 3O2 | Reduction of oxygen (3O2) during electron transport during the photosynthesis process in chloroplasts or during oxidative phosphorylation in the mitochondria 3O2 + e− → O2•− 3O2 + Xanthine (Xanthine oxidase) → O2•− + uric acid 3O2 + NADPH (NADPH oxidase) → O2•− + NADP+ + H+ | 2–4 µs | Triggering the formation of more ROS which further participates in membrane lipid peroxidation | PSI in Chloroplast oxidative phosphorylation in mitochondria Peroxisomes Plasma membrane | SOD, ASc, glutathione (GSH), flavonoids, Cu |
| H2O2 Two electron reduction of 3O2 or univalent reduction of O2•− | Protonation reaction (acidic conditions) O2•− + H+ + HO2• →H2O2 + 2O2 Glycolate + O2 (Glycolate oxidase) → H2O2 + Glyoxylate Fatty acids (β−oxidation) → Acetyl coenzyme + H2O2 | 1 ms | Act as a signaling molecule (low concentration of H2O2), at high concentration of H2O2 triggers tolerance to various stress, as a regulator of physiological processes (photorespiration and photosynthesis) | Chloroplast, Mitochondria, and Peroxisome | CAT, POD (GPX and APX), peroxiredoxin, ASc, tocopherol, GSH, β-carotene, Se flavonoids, lipoic acid, |
| HO2• | Protonation of superoxide ions O2•− + H+ → HO2• | Attacks PUFA in the negatively charged membrane surface | Mitochondria, microsomes and peroxisomes | ||
| HO• (Three electron reduction of 3O2) | Fenton reaction: H2O2 + Fe2+ → HO• + HO− + Fe3+ Haber-Weiss reactions: O2•− + H2O2 → HO• + HO− + O2 | 1 ps | Attack unsaturated fatty acids in membranes | Mitochondria | ASc, GSH, flavonoids, lipoic acid, proline |
| Nitric oxide (NO•) | L-arginine + O2 Nitric oxide synthase → NO• + Citrulline | Intercellular messenger, the quencher of O2•−, defense against various protozoa, fungi, and mycobacteria | Peroxisome, cytosol | GSH | |
| Peroxynitrite (ONOO−) | NO• + O2•− → ONOO− | React with amino acids residues in enzymes causing inactivation | Peroxiredoxin, Uric acid | ||
| Lipid hydroperoxide | Oxidation of PUFA | Tissue injuries and diseases | Mitochondrial membrane PUFA |
| Characteristics | Photo-Autotrophic | Heterotrophic | Mixotrophic |
|---|---|---|---|
| Carbon assimilation process | Photosynthesis | Aerobic respiration | Photosynthesis and aerobic respiration |
| Energy sources | Light (solar or artificial) | Organic carbon (e.g., glucose, acetic acid glycerol, wastewaters) | Light and organic carbon |
| Carbon sources | Inorganic carbon (CO2, NaHCO3) | Organic carbon | Inorganic and organic |
| Light energy | Mandatory | Not required | Not essential |
| Photo-inhibition effect | High | No effect | Low |
| Growth-limiting factor | Light | Oxygen | Light and oxygen |
| Photo-oxidative damage risk | High | Very low | Protected by accumulating oxygen |
| Types of bioreactors | Photo-bioreactors and open ponds | Fermenters | Photo-bioreactors, open ponds |
| Availability of bioreactor’s vessels | Photo-bioreactor | Commercially available | Bioreactor |
| Bioreactor’s surface tovolume ratio (m2/m3) | High | Low | High |
| Sterility | Usually sanitised | Sterility required | Not required |
| Contamination risk | Low | High | Medium |
| Harvesting cost | High due to the low density of biomass | Low due to high biomass concentration | Low due to high biomass concentration |
| Bioreactor set up cost (per unit of volume) | High | Low | High |
| Bioreactor operation cost (per kg of biomass) | Low | Medium | High |
| Scale-up | Design optimisation required | Easy | Design optimisation required |
| Efficiency in wastewater treatment | Low | High | High |
| Growth rate | Low | Medium | High |
| Biomass density (g L−1) | Low | Medium | High |
| Lipids (%) | Low | High | High |
| Proteins (%) | High | Low | Medium |
| Carbohydrates (%) | Low | High | Low |
| Chlorophyll and β-carotene (%) | High | Low | Medium |
| Lutein (%) | Medium | High | High |
| Stress Conditions | Strains | Optimum Light (µmol Photon m−2 s−1) | Salinity (NaCl) (M) | Temperature (°C) | Antioxidant Enzymatic Response | References | |
|---|---|---|---|---|---|---|---|
| Salinity | 0.05–3.00 M NaCl | D. tertiolecta (UTEX999) | 150 Continuous light | 0.1–0.5 | 26 | No effect on SOD and CAT, GTR activities APX increased (2-fold) at high salinities (0.2–3 M) | [125] |
| 0.5–5.5 M NaCl | D. salina | 150 Light: dark cycle (12:12 h) | 1.5 | (25 ± 2) | CAT activity decreases with salinity SOD increased (0.5–2 M) and then decrease APX decreases (0.5–2 M) and then constant | [122] | |
| 0.05–4.0 M NaCl | D. salina and D. tertiolecta | 150 Light: dark cycle (12:12 h) | 1.25 | (25 ± 2) | CAT and SOD activities increase at hypo saline condition in D. tertiolecta APX activity in D. tertiolecta higher at hypo saline and lower at hyper saline condition GPX activity decreased in D. tertiolecta at higher saline condition (>1.25 M) | [124] | |
| 1–4.0 M NaCl | D. salina | 54 (Continuous) | 2 | 22 | APX increase (171%) at 4 M compared to 2 M | [153] | |
| Light | Outdoor with natural UVR | D. tertiolecta | 250 Light: dark cycle (12:12 h) | - | 20 | SOD activity slightly changed in short and unaltered for long term exposure but no change in APX and GTX activity | [130] |
| UV-B for 6 days | D. salina | 60–80 Light: dark cycle (12:12 h) | - | 20 | CAT activity increased on 3rd day SOD activity increased on 4th day POD activity increased on 3rd day | [128] | |
| UV-B and UV-C for 24 h | D. bardawil and D. salina | 150 UV-A | - | 26 | APX increased by UV -A | [131] | |
| UV-B for 4 h | D. salina | 920 | - | 25 | CAT, SOD and POD activity increased | [34] | |
| Nutrients | Depletion of N, P, S, NP, NS, NPS | D. salina | 60 (Continuous) | - | 30 | SOD and CAT activity increased | [137] |
| KNO3 (0.05, 0.5, 5 mM) | D. salina | 150 Continuous light | - | 25 | CAT, SOD and APX activity decreased with increased nitrogen concentration | [138] | |
| Light and nutrients | Light intensity and depletion of Mn, Zn, Fe, N | D. salina | 70 Light: dark cycle (16:8 h) | 20 | Mn-SOD overexpressed | [36] | |
| UV-B 2 h day−1 for 3 days and deprivation of nitrogen | D. salina | 85 Continuous light | 25±5 | POD and APX activity increased | [37] | ||
| Light and temperature | Low temperature and low light for 24 h | D. salina (Gh-U) | 100 Light: dark (16:8 h) | 28 ± 0.5 | SOD activity increased APX activity increased at low light level GR, MDHAR increased at low temperature | [135] | |
| Low temperature and medium light for 24 h | D. salina (IR-1 and Gh-U) | 100 Light: dark (16:8 h) | 28 ± 0.5 | POD activity increased, GTX increased in strain Gh-U but not in IR-1 at low temperature GR, DHAR activity increased in Gh-U than IR-1 | [35] | ||
| Metals | Cu2+ (1–20 µM) for 24–72 h | D. salina and D. tertiolecta | 54 Continuous light | 22 | APX activity increased | [143] | |
| Hg2+ (0–40 µM) for 48 h and 28 days | D. tertiolecta | 54 (Continuous light) | (22 ± 2) | APX activity increased | [141] | ||
| Cd2+ (0–20 µg L−1) for 24, 48,72, and 96 h | D. viridis | 117 (Light: dark) (12:12 h) | (28 ± 2) | Pigment production decreased | [154] | ||
| Cr6+ (40 ppm) for 24 h | D. salina and D. tertiolecta | 46 White fluorescent light | 25 | SOD activity decreased Chlorophyll ‘a’ content decreased with increase in the exposure time | [155] | ||
| Silver nanoparticles (10, 50, 100 and 200 mg L−1 Ag-NPs) for 8 days | (D. tertiolecta) | 100 Continuous light | 25 | CAT activity increased until 6 days then decreased, SOD activity increased up to 4 days, POD activity increased until 6 days | [140] | ||
| Chemicals | Carbamazepine (CBZ) (0–200 mg L−1) 24, 48,72, and 96 h | D. tertiolecta (CCAP19/6B) | 58 Continuous | (24 ± 1) | Increased carotenoid | [156] | |
| SDBS and CTAC for 48 h | D. bardawil | 144 (Light: dark) (14:10 h) | 26 | CAT and SOD activity increased | [148] | ||
| Trichlorfon and dimehypo | D. salina | 54 (Light: dark) (14:10 h) | 26 | CAT activity increased | [151] | ||
| 2-chlorophenol for 24, 48, and 96 h | D. salina | 100 (Continuous daylight) | (25 ± 1) °C | SOD, CAT, GTX activity increased APX activity decreased | [149] | ||
| Chemical and salinity | (1, 2, 3 M NaCl) and PG (propyl gallate) for 48 h | D. salina (UTEX 200) | 70 Light: dark (16:8 h) | 25 | Maximum CAT activity at 2 M NaCl and no change in SOD activity at 1 and 3 M NaCl Minor increase in APX activity at 3 M NaCl | [152] | |
| Chemical and temperature | 10 °C and 5 °C, Methylene blue and norflurazon | D. salina (IPASS D-294) | 73.6 White fluorescent light | 27 | CAT activity increased Chlorophyll and carotenoid content decreased with temperature | [136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, U.K.; Nielsen, B.V.; Milledge, J.J. Antioxidant Production in Dunaliella. Appl. Sci. 2021, 11, 3959. https://doi.org/10.3390/app11093959
Roy UK, Nielsen BV, Milledge JJ. Antioxidant Production in Dunaliella. Applied Sciences. 2021; 11(9):3959. https://doi.org/10.3390/app11093959
Chicago/Turabian StyleRoy, Uttam Kumer, Birthe Vejby Nielsen, and John James Milledge. 2021. "Antioxidant Production in Dunaliella" Applied Sciences 11, no. 9: 3959. https://doi.org/10.3390/app11093959
APA StyleRoy, U. K., Nielsen, B. V., & Milledge, J. J. (2021). Antioxidant Production in Dunaliella. Applied Sciences, 11(9), 3959. https://doi.org/10.3390/app11093959