
Hot Resistance of Spores from the Thermophilic Bacillus horneckiae SBP3 of Shallow Hydrothermal Vent Origin Elucidated by Spectroscopic Analyses

 ,
,  ,
, 



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Spore Preparation
2.2. FTIR Analysis
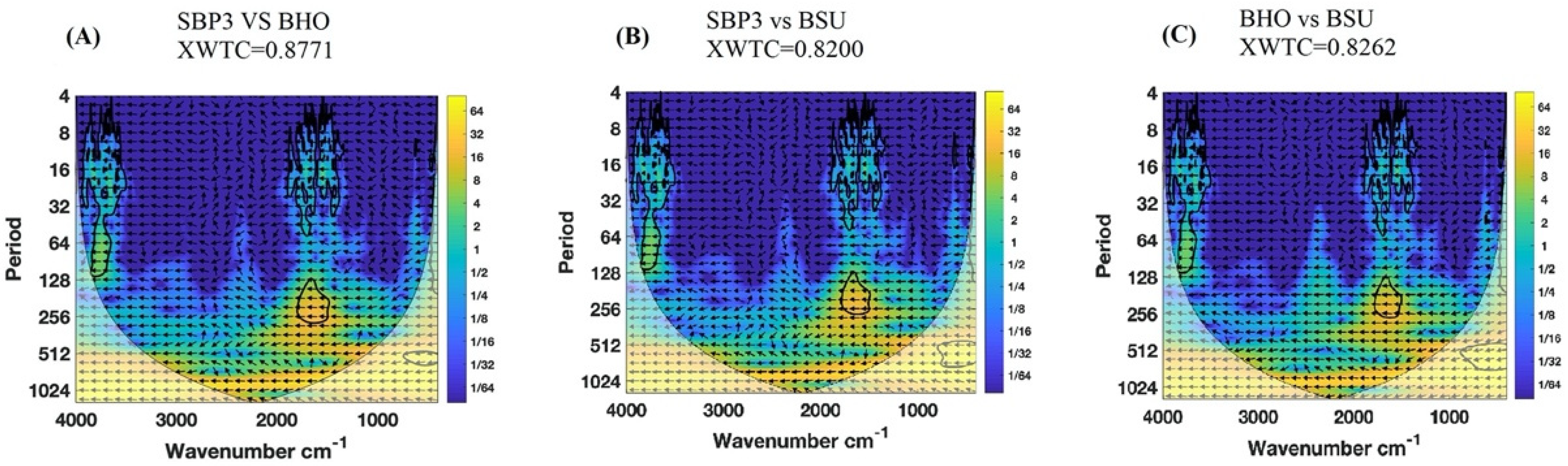
2.3. Wavelet Cross-Correlation
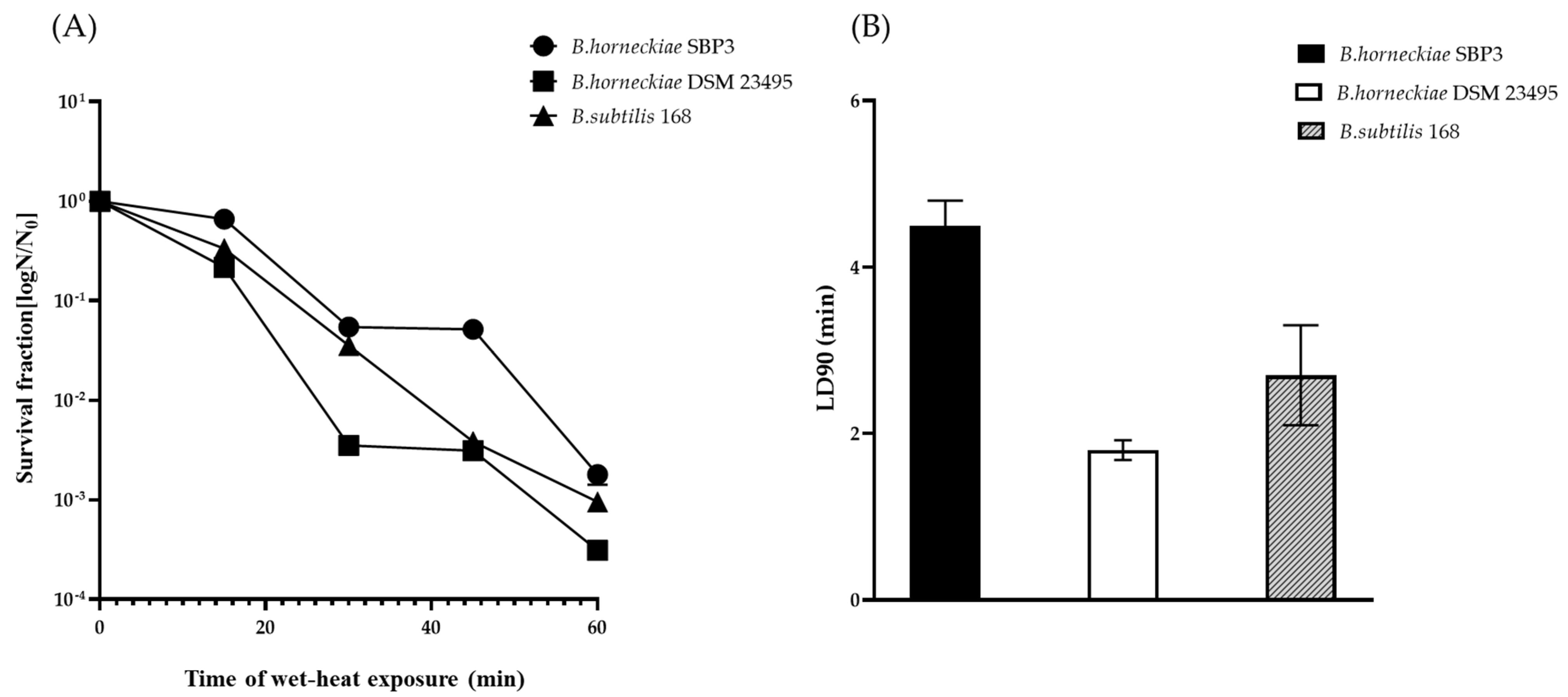
2.4. Spore Resistance to Wet Heat
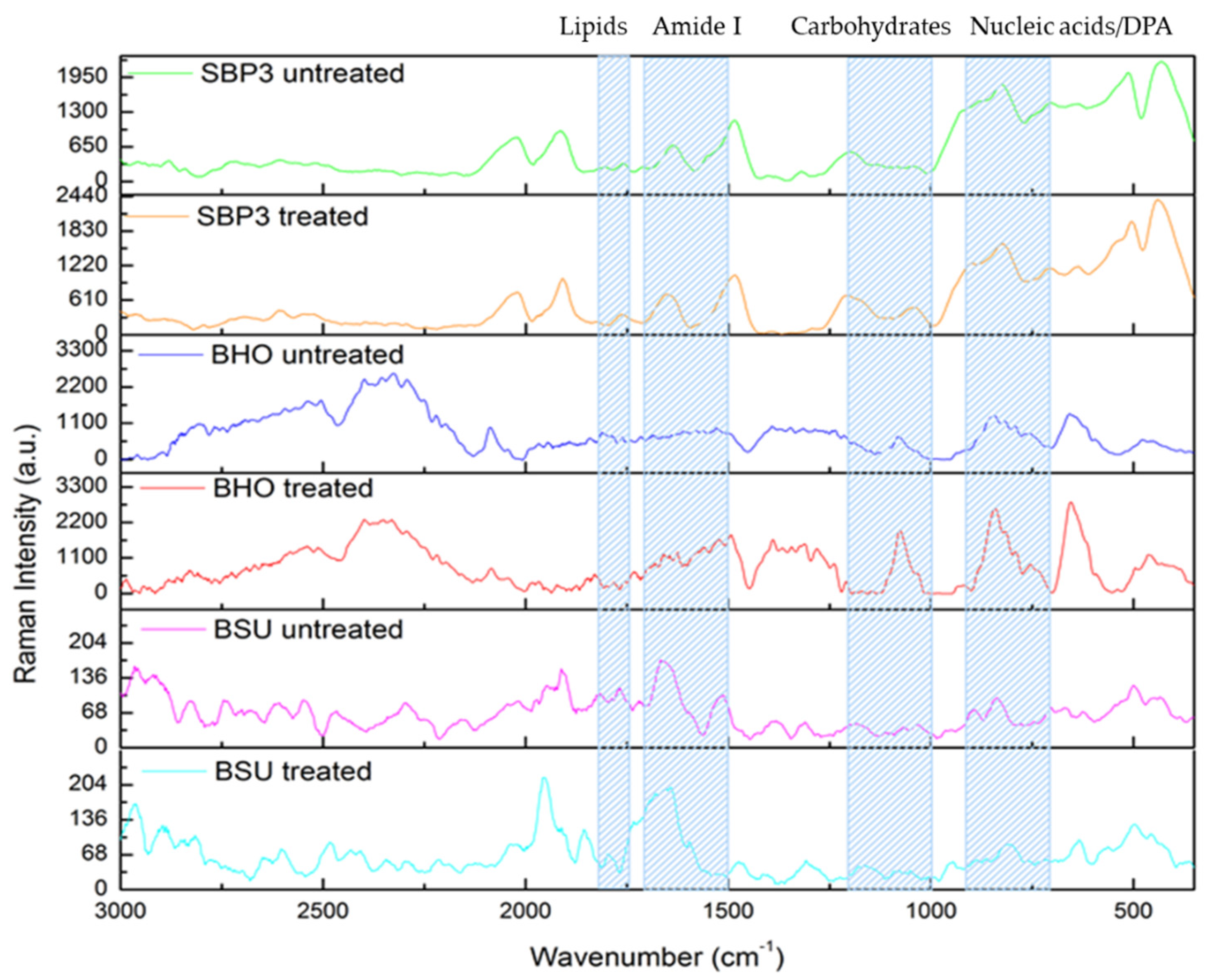
2.5. Spectroscopic Raman Analysis
2.6. Statistical Analysis
3. Results
3.1. The Morphological Spore Analysis
3.2. FTIR Analysis
3.3. Resistance to Wet Heat (98 °C)
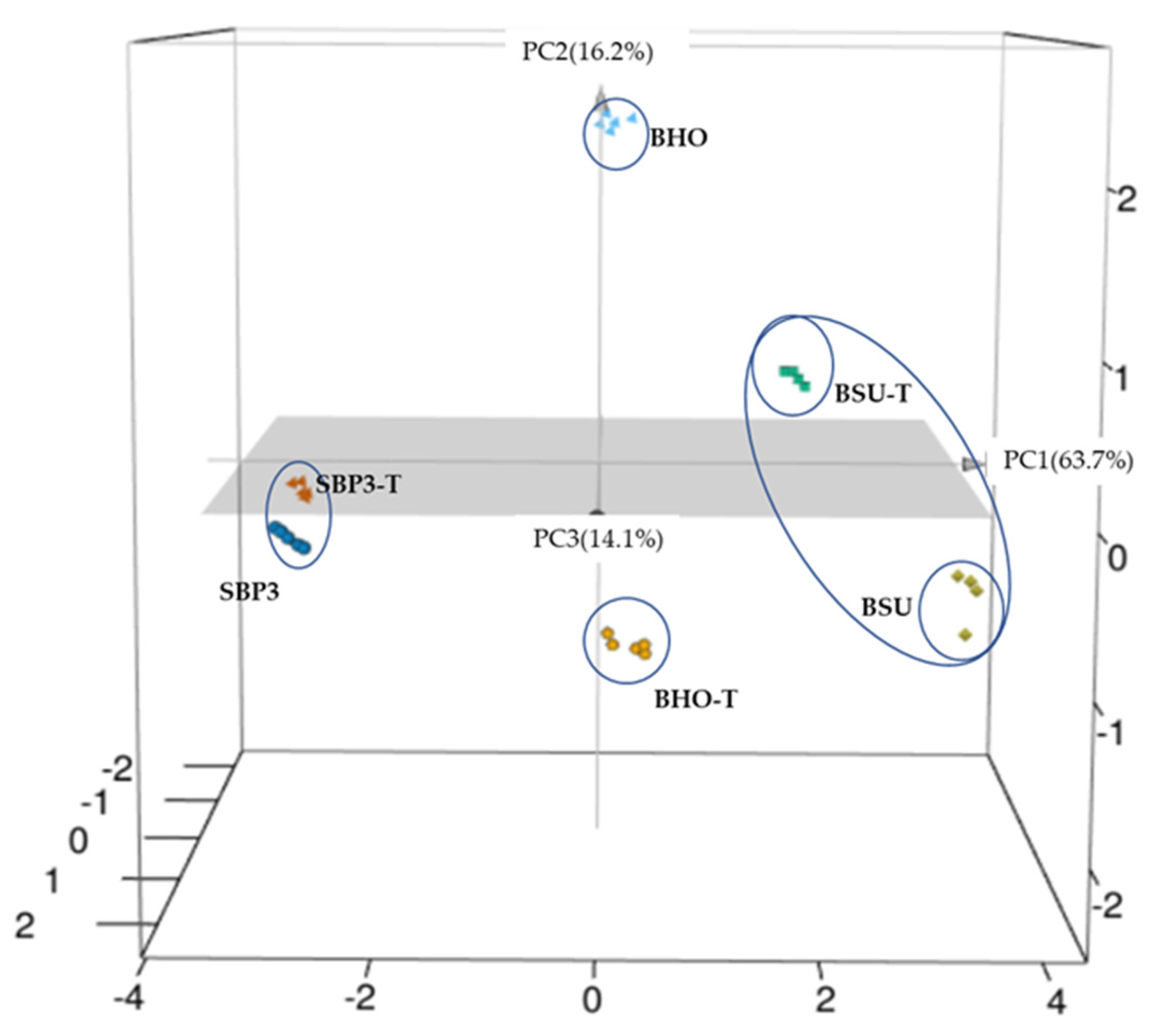
3.4. Raman Spectroscopy Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cortesão, M.; Fuchs, F.M.; Commichau, F.M.; Eichenberger, P.; Schuerger, A.C.; Nicholson, W.L.; Setlow, P.; Moeller, R. Ba-cillus subtilis Spore Resistance to Simulated Mars Surface Conditions. Front. Microbiol. 2019, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zammuto, V.; Fuchs, F.M.; Fiebrandt, M.; Stapelmann, K.; Ulrich, N.J.; Maugeri, T.L.; Pukall, R.; Gugliandolo, C.; Moeller, R. Comparing Spore Resistance ofBacillusStrains Isolated from Hydrothermal Vents and Spacecraft Assembly Facilities to Environmental Stressors and Decontamination Treatments. Astrobiology 2018, 18, 1425–1434. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Moeller, R.; Schuerger, A.C.; Reitz, G.; Nicholson, W.L. Protective Role of Spore Structural Components in Determining Bacillus subtilis Spore Resistance to Simulated Mars Surface Conditions. Appl. Environ. Microbiol. 2012, 78, 8849–8853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zammuto, V.; Rizzo, M.G.; De Plano, L.M.; Franco, D.; Guglielmino, S.; Caccamo, M.T.; Magazù, S.; Fujimori, A.; Lo Giudice, A.; Guglielmin, M.; et al. Effects of Heavy Ion Particle Irradiation on Spore Germina-tion of Bacillus spp. from Extremely Hot and Cold Environments. Life 2020, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Lentini, V.; Spanò, A.; Maugeri, T. New bacilli from shallow hydrothermal vents of Panarea Island (Italy) and their biotechnological potential. J. Appl. Microbiol. 2012, 112, 1102–1112. [Google Scholar] [CrossRef]
- Zammuto, V.; Gugliandolo, C. Resistance to high temperatures of spores from bacilli of shallow hydrothermal vent origins. AAPP Phys. Math. Nat. Sci. 2019, 97, 28. [Google Scholar] [CrossRef]
- Vaishampayan, P.; Probst, A.; Krishnamurthi, S.; Ghosh, S.; Osman, S.; McDowall, A.; Ruckmani, A.; Mayilraj, S.; Venkateswaran, K. Bacillus horneckiae sp. nov., isolated from a spacecraft-assembly clean room. Int. J. Syst. Evol. Microbiol. 2010, 60, 1031–1037. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, M.T.; Gugliandolo, C.; Zammuto, V.; Magazù, S. Thermal properties of an exopolysaccharide produced by a ma-rine thermotolerant Bacillus licheniformis by ATR-FTIR spectroscopy. Int. J. Biol. Macromol. 2019, 145, 77–83. [Google Scholar] [CrossRef]
- Maisano, G.; Majolino, D.; Migliardo, P.; Venuto, S.; Aliotta, F.; Magazú, S. Sound velocity and hydration phenomena in aqueous polymeric solutions. Mol. Phys. 1993, 78, 421–435. [Google Scholar] [CrossRef]
- Caccamo, M.T.; Magazù, S. Tagging the oligomer-to-polymer crossover on EG and PEGs by infrared and Raman spectros-copies and by wavelet cross-correlation spectral analysis. Vib. Spectr. 2016, 85, 222–227. [Google Scholar] [CrossRef]
- Cannuli, A.; Caccamo, M.T.; Castorina, G.; Colombo, F.; Magazù, S. Laser Techniques on Acoustically Levitated Droplets. EPJ Web Conf. 2018, 167, 5010. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, M.T.; Cannuli, A.; Magazù, S. Wavelet analysis of near-resonant series RLC circuit with time-dependent forcing frequency. Eur. J. Phys. 2018, 39, 045702. [Google Scholar] [CrossRef]
- Nawaz, H.; Bonnier, F.; Knief, P.; Howe, O.; Lyng, F.M.; Meade, A.D.; Byrne, H.J. Evaluation of the potential of Raman microspectroscopy for prediction of chemotherapeutic response to cisplatin in lung adenocarcinoma. Analyst 2010, 135, 3070–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Managò, S.; Valente, C.; Mirabelli, P.; Circolo, D.; Basile, F.; Corda, D.; De Luca, A.C. A reliable Raman-spectroscopy-based approach for diagnosis, classification and follow-up of B-cell acute lymphoblastic leukemia. Sci. Rep. 2016, 6, 24821. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. Available online: https://www.stats.ox.ac.uk/pub/MASS4/ (accessed on 7 May 2021).
- Vongsvivut, J.; Heraud, P.; Zhang, W.; Kralovec, J.A.; McNaughton, D.; Barrow, C.J. Rapid Determination of Protein Con-tents in Microencapsulated Fish Oil Supplements by ATR-FTIR Spectroscopy and Partial Least Square Regression (PLSR) Analysis. Food Bioprocess Tech. 2013, 7, 265–277. [Google Scholar] [CrossRef]
- Guillén, M.D.; Cabo, N. Characterization of edible oils and lard by fourier transform infrared spectroscopy. Relationships between composition and frequency of concrete bands in the fingerprint region. J. Am. Oil Chem. Soc. 1997, 74, 1281–1286. [Google Scholar] [CrossRef]
- Filip, Z.; Herrmann, S.; Kubát, J. FT-IR spectroscopic characteristics of dierently cultivated Bacillus subtilis. Microbiol. Res. 2004, 159, 257–262. [Google Scholar] [CrossRef]
- Böcker, U.; Wubshet, S.G.; Lindberg, D.; Afseth, N.K. Fourier-transform infrared spectroscopy for characterization of protein chain reductions in enzymatic reactions. Analyst 2017, 142, 2812–2818. [Google Scholar] [CrossRef]
- Serrano, V.; Liu, W.-G.; Franzen, S. An Infrared Spectroscopic Study of the Conformational Transition of Elastin-Like Poly-peptides. Biophys. J. 2007, 93, 2429–2435. [Google Scholar] [CrossRef] [Green Version]
- Perkins, D.L.; Lovell, C.R.; Bronk, B.V.; Setlow, B.; Setlow, P.; Myrick, M.L. Fourier transform infrared reflectance mi-cro-spectroscopy study of Bacillus subtilis engineered without dipicolinic acid: The contribution of calcium dipicolinate to the mid-infrared absorbance of Bacillus subtilis endospores. Appl. Spectrosc. 2005, 59, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Bizani, D.; Motta, A.S.; Morrissy, J.A.C.; Terra, R.M.S.; Souto, A.A.; Brandelli, A. Antibacterial activity of cerein 8A, a bacte-riocin-like peptide produced by Bacillus cereus. Int. Microbiol. 2005, 8, 125–131. [Google Scholar]
- Rohman, A.; Man, Y.C. Fourier transform infrared (FTIR) spectroscopy for analysis of extra virgin olive oil adulterated with palm oil. Food Res. Int. 2010, 43, 886–892. [Google Scholar] [CrossRef]
- Zelig, U.; Mordechai, S.; Shubinsky, G.; Sahu, R.K.; Huleihel, M.; Leibovitz, E.; Nathan, I.; Kapelushnik, J. Pre-screening and follow-up of childhood acute leukemia using biochemical infrared analysis of peripheral blood mononuclear cells. Biochim. Biophys. Acta 2011, 1810, 827–835. [Google Scholar] [CrossRef] [PubMed]
- De Gelder, J.; Scheldeman, P.; Leus, K.; Heyndrickx, M.; Vandenabeele, P.; Moens, L.; De Vos, P. Raman spectroscopic study of bacterial endospores. Anal. Bioanal. Chem. 2007, 389, 2143–2151. [Google Scholar] [CrossRef] [PubMed]
- Çulha, M.; Adigüzel, A.; Yazici, M.M.; Kahraman, M.; Slahin, F.; Güllüce, M. Characterization of Thermophilic Bacteria Using Surface-Enhanced Raman Scattering. Appl. Spectrosc. 2008, 62, 1226–1232. [Google Scholar] [CrossRef]
- Kong, L.; Setlow, P.; Li, Y.-Q. Analysis of the Raman spectra of Ca2+-dipicolinic acid alone and in the bacterial spore core in both aqueous and dehydrated environments. Analyst 2012, 137, 3683–3689. [Google Scholar] [CrossRef]
- Nelson, W.H.; Dasari, R.; Feld, M.; Sperry, J.F. Intensities of Calcium Dipicolinate andBacillus SubtilisSpore Raman Spectra Excited with 244 nm Light. Appl. Spectrosc. 2004, 58, 1408–1412. [Google Scholar] [CrossRef]
- Zhang, P.; Kong, L.; Setlow, P.; Li, Y.Q. Characterization of wetheat inactivation of single spores of Bacillus species by du-al-trap Raman spectroscopy and elastic light scattering. Appl. Environ. Microbiol. 2010, 76, 1796–1805. [Google Scholar] [CrossRef] [Green Version]
- Wolkers, W.F.; Oliver, A.E.; Tablin, F.; Crowe, J.H. A Fourier-transform infrared spectroscopy study of sugar glasses. Carbohydr. Res. 2004, 339, 1077–1085. [Google Scholar] [CrossRef]
- Noothalapati, H.; Sasaki, T.; Kaino, T.; Kawamukai, M.; Ando, M.; Hamaguchi, H.O.; Yamamoto, T. Label-free chemical im-aging of fungal spore walls by Raman microscopy and multivariate curve resolution. Anal. Sci. Rep. 2016, 6, 27789. [Google Scholar] [CrossRef] [Green Version]
- Rusciano, G.; Capriglione, P.; Pesce, G.; Abete, P.; Carnovale, V.; Sasso, A. Raman spectroscopy as a new tool for early de-tection of bacteria in patients with cystic fibrosis. Laser Phys. Lett. 2013, 10, 075603. [Google Scholar] [CrossRef]
- Atrih, A.; Foster, S.J. Bacterial endospores the ultimate survivors. Int. Dairy J. 2002, 12, 217–223. [Google Scholar] [CrossRef]
- Driks, A. Bacillus subtilis Spore Coat. Microbiol. Mol. Biol. Rev. 1999, 63, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.D. Bacterial spores and chemical sporicidal agents. Clin. Microbiol. Rev. 1990, 3, 99–119. [Google Scholar] [CrossRef]
- Cho, W.-I.; Chung, M.-S. Bacillus spores: A review of their properties and inactivation processing technologies. Food Sci. Biotechnol. 2020, 29, 1447–1461. [Google Scholar] [CrossRef] [PubMed]
- Coleman, W.; Setlow, P. Analysis of damage due to moist heat treatment of spores of Bacillus subtilis. J. Appl. Microbiol. 2009, 106, 1600–1607. [Google Scholar] [CrossRef]
- Belliveau, J.W.; Kwong, K.K.; Kennedy, D.N.; Baker, J.R.; Stern, C.E.; Benson, R.; Chesler, D.A.; Weisskoff, R.M.; Cohen, M.S.; Tootell, R.B.H.; et al. Magnetic resonance imaging mapping of brain function human visual cor-tex. Investig. Radiol. 1992, 27, S59. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spore resistance properties. In The Bacterial Spore: From Molecules to Systems; American Society of Microbiology: Washington, DC, USA, 2016; pp. 201–215. [Google Scholar]
- Mysliwiec, T.H.; Tierno, A.F.; Aurentz, D.J. Characterization of Bacillus subtilis sporulation and bacteriophage infection via FT-IR spectroscopy. J. Spect. 2009, 23, 456049. [Google Scholar] [CrossRef]
- Subramanian, A.; Ahn, J.; Balasubramaniam, V.M.; Rodriguez-Saona, L. Monitoring Biochemical Changes in Bacterial Spore during Thermal and Pressure-Assisted Thermal Processing using FT-IR Spectroscopy. J. Agric. Food Chem. 2007, 55, 9311–9317. [Google Scholar] [CrossRef]
- Guicheteau, J.; Argue, L.; Emge, D.; Hyre, A.; Jacobson, M.; Christesen, S. Bacillus Spore Classification via Surface-Enhanced Raman Spectroscopy and Principal Component Analysis. Appl. Spectrosc. 2008, 62, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.-C.; Cai, Y.-D. Predicting protein structural class by functional domain composition. Biochem. Biophys. Res. Commun. 2004, 321, 1007–1009. [Google Scholar] [CrossRef] [PubMed]
- Koga, Y. Thermal Adaptation of the Archaeal and Bacterial Lipid Membranes. Archaea 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.S.; Nakamura, L.K.; Cohan, F.M. Bacillus mojavensis sp. nov., distinguishable from Bacillus subtilis by sexual iso-lation, divergence in DNA sequence, and differences in fatty acid composition. Int. J. Syst. Evol. Microbiol. 1994, 44, 256–264. [Google Scholar]
- Coleman, W.H.; Chen, D.; Li, Y.-Q.; Cowan, A.E.; Setlow, P. How Moist Heat Kills Spores of Bacillus subtilis. J. Bacteriol. 2007, 189, 8458–8466. [Google Scholar] [CrossRef] [Green Version]
- Besten, H.M.D.; Wells-Bennik, M.H.; Zwietering, M.H. Natural Diversity in Heat Resistance of Bacteria and Bacterial Spores: Impact on Food Safety and Quality. Annu. Rev. Food Sci. Technol. 2018, 9, 383–410. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavenumber Values (cm−1) | Band Assignment | References |
|---|---|---|
| 3300–3200 | H-bond and OH group of alcohol, phenols and organic acid including nucleic acids and proteins amide A | [20] |
| 3100–3000 | n(C-H) of cis C=H bonds | [21] |
| ~2925 | Lipids | [21] |
| 1700–1750 | Protein and esters of muramic acid and ester fatty acid group | [22] |
| 1660–1628 | Amide I peptidic conformation | [23] |
| ~1548 | Amide II peptidic conformation | [24] |
| ~1380 | CH2 and CH3 bending from lipids, DPA amide III | [25] |
| ~1066 | Ring vibrations of carbohydrates | [21] |
| ~966 | CH conjugated trans, trans isomers | [26] |
| ~780 | Nucleic acids, sugar-phosphate vicration | [27] |
| Raman Band/Peak (cm−1) | Tentative Assignment of Functional Group | References |
|---|---|---|
| 622 | Proteins | [29] |
| 661 | CaDPA | [30] |
| 725 | DNA | [31] |
| 780 | DNA | [31] |
| 824 | CaDPA | [31] |
| 864 | C-C stretch | [29] |
| 1004 | Phenilanine | [32] |
| 1017 | CaDPA | [29] |
| 1060–1070 | Glucose-saccharide band | [33] |
| 1115 | Proteins | [29] |
| 1200–1244 | Proteins, amide III | [29] |
| 1336 | DNA | [31] |
| 1395 | CaDPA | [30] |
| 1448 | Proteins | [32] |
| 1485 | DNA | [31] |
| 1555 | CaDPA | [31] |
| 1582 | CaDPA | [31] |
| 1624 | Proteins | [34] |
| 1645–1674 | Proteins, amide I, lipids | [32] |
| 1700–1736 | Lipids | [35] |
| Bacillus horneckiae SBP3 DSM 103063 Zammuto et al. [3] | Bacillus horneckiae DSM 23495T Vayshampayan et al. [10] | Bacillus subtilis 168 Roberts et al. [48] | |
|---|---|---|---|
| Temperature range (°C) | 25–45 | 4–40 | 25–37 |
| Fatty acids | |||
| Straight chain saturated | |||
| C16:0 | 0.8 | 0.5 | 3.14 |
| Branched saturated | |||
| iso-C14:0 | 3.4 | 4 | 1.13 |
| iso-C15:0 | 63.1 | 54.3 | 29.27 |
| iso-C16:0 | 3.5 | 2.5 | |
| iso-C17:0 | 2 | 1.3 | 9.59 |
| anteiso-C15:0 | 8.8 | 19.9 | 40.19 |
| anteiso-C17:0 | 1.8 | 2.3 | 9.38 |
| Monounsaturated | |||
| C16:1 ω7 c-alchol | 7.8 | 6.8 | 0.23 |
| iso-C17:1 ω10 c | 3.7 | 2.3 | 1.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zammuto, V.; Caccamo, M.T.; Magazù, S.; Spanò, A.; Guglielmino, S.; Gugliandolo, C. Hot Resistance of Spores from the Thermophilic Bacillus horneckiae SBP3 of Shallow Hydrothermal Vent Origin Elucidated by Spectroscopic Analyses. Appl. Sci. 2021, 11, 4256. https://doi.org/10.3390/app11094256
Zammuto V, Caccamo MT, Magazù S, Spanò A, Guglielmino S, Gugliandolo C. Hot Resistance of Spores from the Thermophilic Bacillus horneckiae SBP3 of Shallow Hydrothermal Vent Origin Elucidated by Spectroscopic Analyses. Applied Sciences. 2021; 11(9):4256. https://doi.org/10.3390/app11094256
Chicago/Turabian StyleZammuto, Vincenzo, Maria Teresa Caccamo, Salvatore Magazù, Antonio Spanò, Salvatore Guglielmino, and Concetta Gugliandolo. 2021. "Hot Resistance of Spores from the Thermophilic Bacillus horneckiae SBP3 of Shallow Hydrothermal Vent Origin Elucidated by Spectroscopic Analyses" Applied Sciences 11, no. 9: 4256. https://doi.org/10.3390/app11094256
APA StyleZammuto, V., Caccamo, M. T., Magazù, S., Spanò, A., Guglielmino, S., & Gugliandolo, C. (2021). Hot Resistance of Spores from the Thermophilic Bacillus horneckiae SBP3 of Shallow Hydrothermal Vent Origin Elucidated by Spectroscopic Analyses. Applied Sciences, 11(9), 4256. https://doi.org/10.3390/app11094256