
Glucose Isomerase: Functions, Structures, and Applications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Function
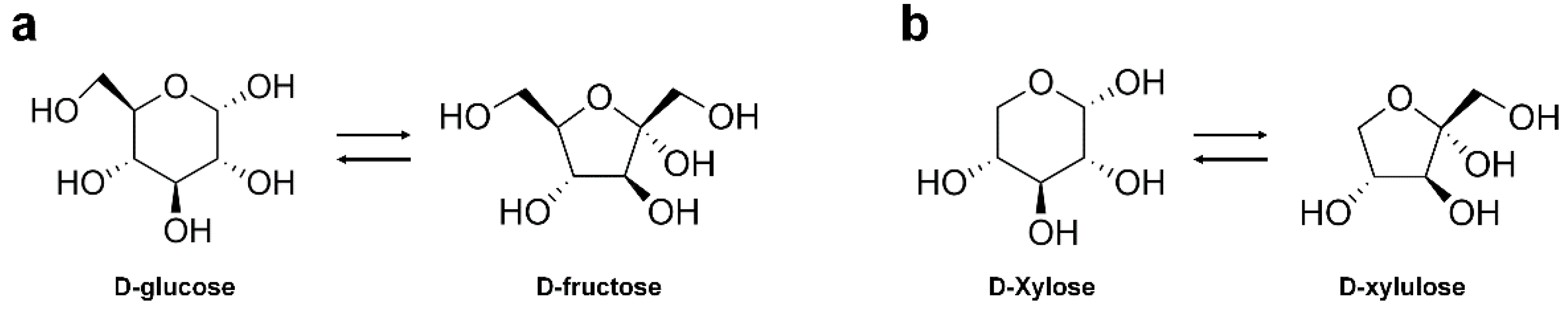
2.1. Substrate Specificity
2.2. Metal Ions
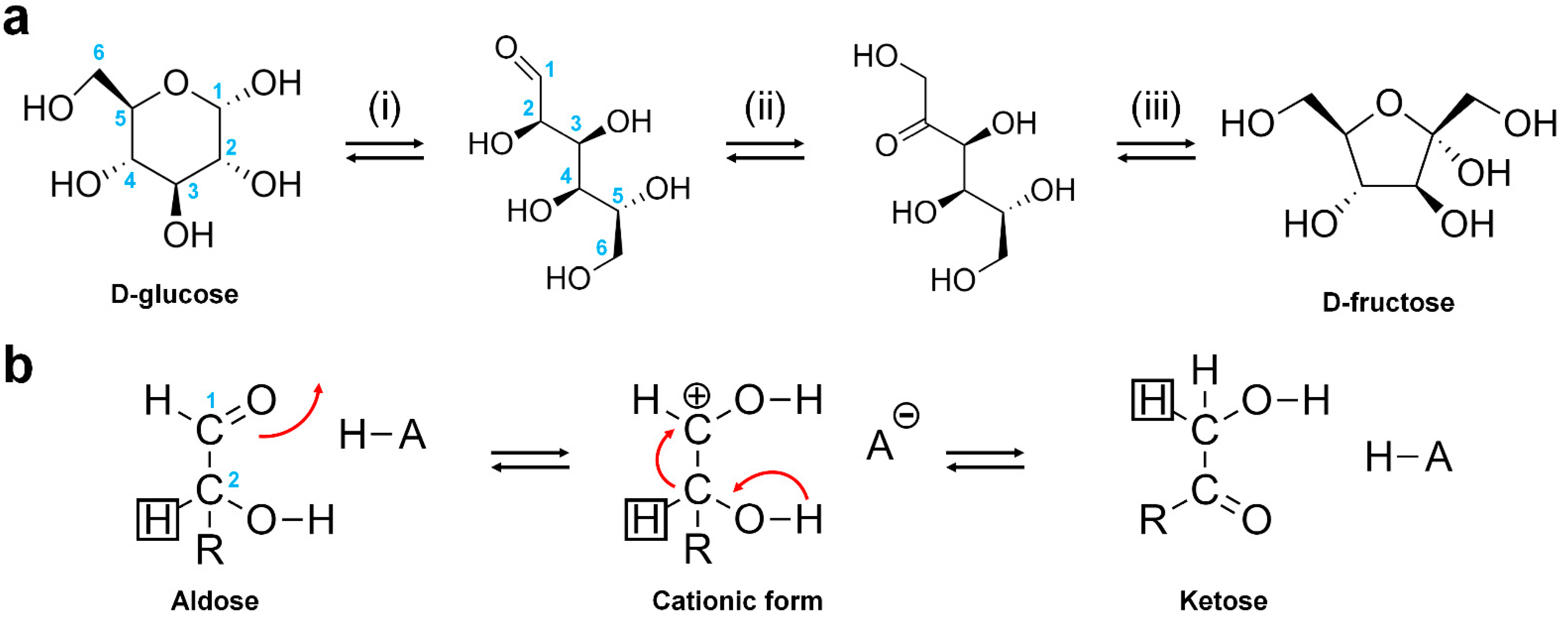
2.3. Reaction Mechanism
3. Structure
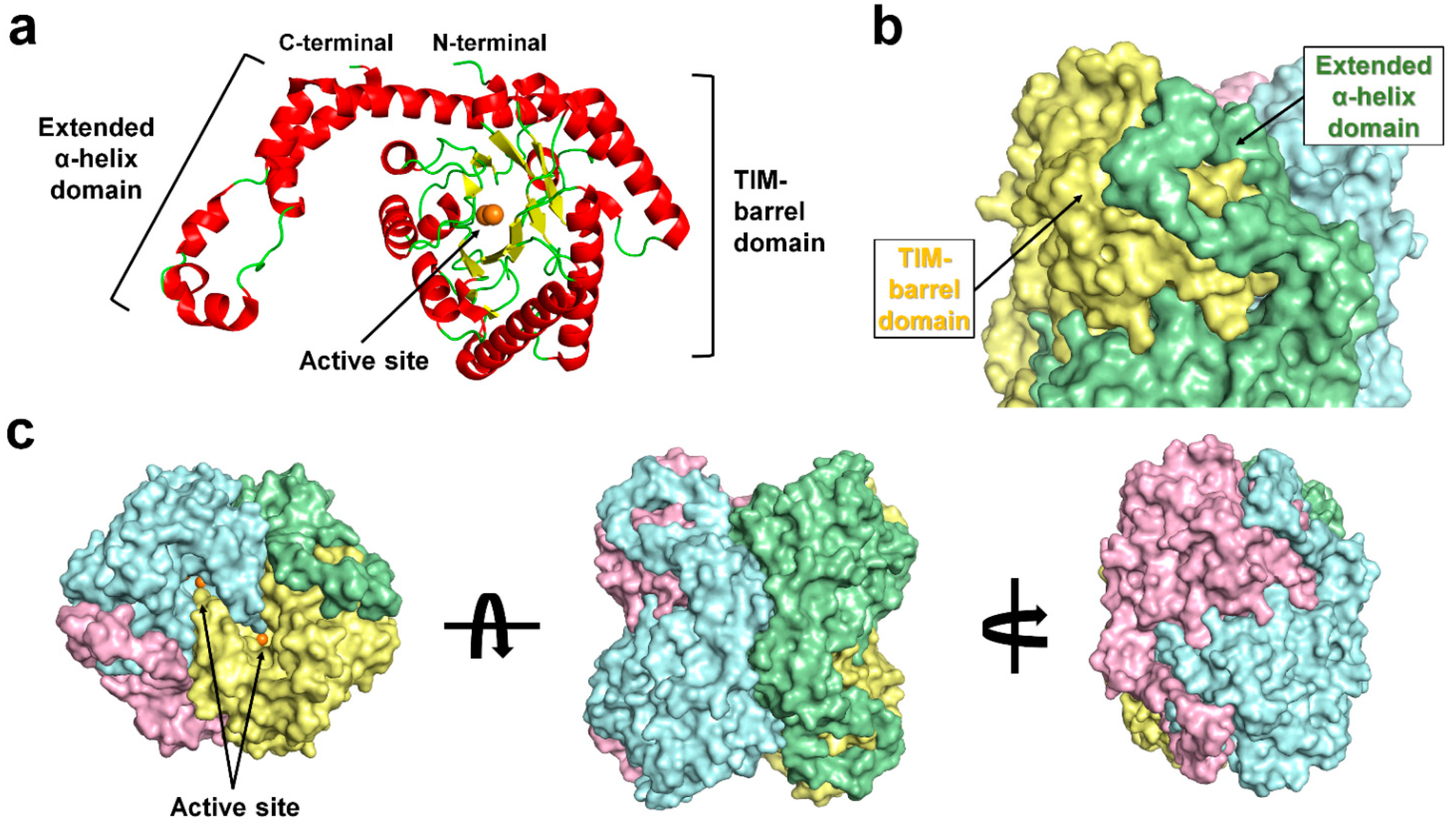
3.1. Overall Structure

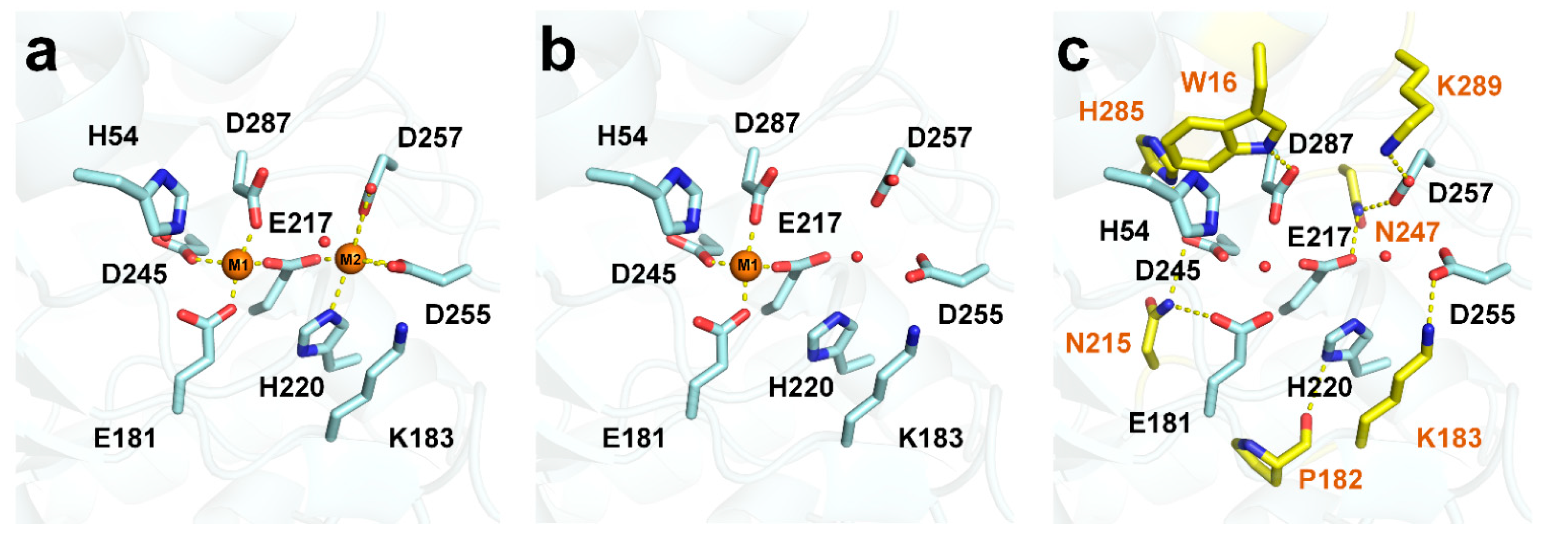
3.2. Metal-Binding State at the GI Active Site
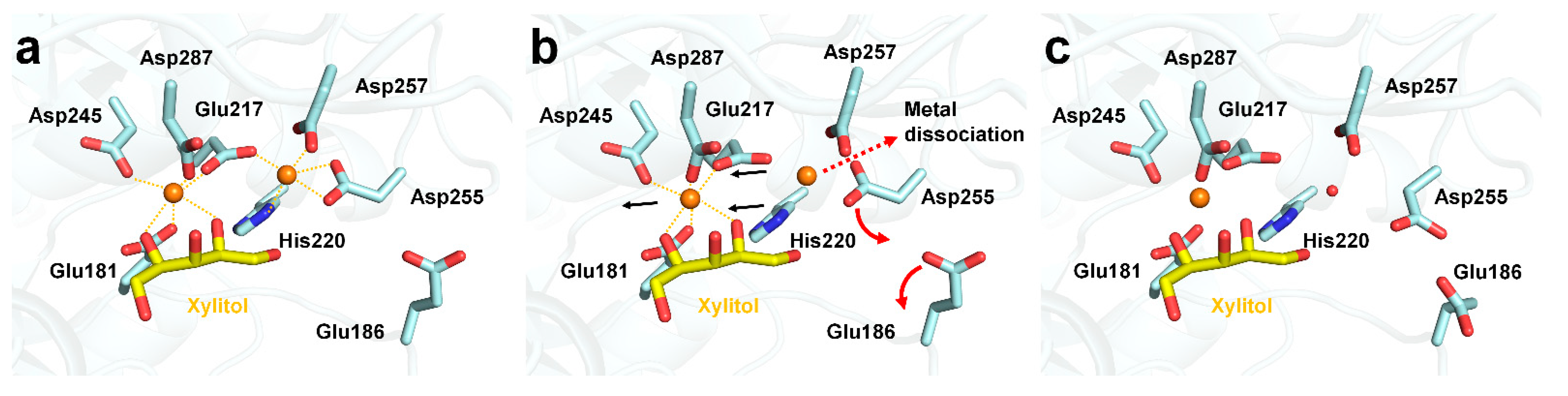
3.3. Xylitol Binding to the Active Site of GI
4. Application of GI
4.1. High-Fructose Corn Syrup (HFCS)
4.2. Ethanol Production
5. Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhosale, S.H.; Rao, M.B.; Deshpande, V.V. Molecular and industrial aspects of glucose isomerase. Microbiol. Rev. 1996, 60, 280–300. [Google Scholar] [CrossRef]
- Kilara, A.; Shahani, K.M. The use of immobilized enzymes in the food industry: A review. CRC Crit. Rev. Food Sci. Nutr. 1979, 12, 161–198. [Google Scholar] [CrossRef]
- Deshpande, V.; Rao, M. Glucose Isomerase. In Enzyme Technology; Springer: New York, NY, USA, 2006; pp. 239–252. [Google Scholar] [CrossRef]
- Singh, R.S.; Singh, T.; Pandey, A. Microbial Enzymes—An Overview. In Advances in Enzyme Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–40. [Google Scholar] [CrossRef]
- Kasumi, T.; Hayashi, K.; Tsumura, N. Purification and Enzymatic Properties of Glucose Isomerase from Streptomyces griseofuscus, S-41. Agric. Biol. Chem. 2014, 45, 619–627. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Kim, D.-Y.; Park, C.-S. Production of l-rhamnulose, a rare sugar, from l-rhamnose using commercial immobilized glucose isomerase. Biocatal. Biotransform. 2017, 36, 417–421. [Google Scholar] [CrossRef]
- Langan, P.; Sangha, A.K.; Wymore, T.; Parks, J.M.; Yang, Z.K.; Hanson, B.L.; Fisher, Z.; Mason, S.A.; Blakeley, M.P.; Forsyth, V.T.; et al. L-Arabinose binding, isomerization, and epimerization by D-xylose isomerase: X-ray/neutron crystallographic and molecular simulation study. Structure 2014, 22, 1287–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Chen, J.; Zhang, W.; Zhang, T.; Guang, C.; Mu, W. Recent research on the physiological functions, applications, and biotechnological production of D-allose. Appl. Microbiol. Biotechnol. 2018, 102, 4269–4278. [Google Scholar] [CrossRef] [PubMed]
- Fehér, C. Novel approaches for biotechnological production and application of L-arabinose. J. Carbohydr. Chem. 2018, 37, 251–284. [Google Scholar] [CrossRef]
- Menavuvu, B.T.; Poonperm, W.; Leang, K.; Noguchi, N.; Okada, H.; Morimoto, K.; Granstrom, T.B.; Takada, G.; Izumori, K. Efficient biosynthesis of D-allose from D-psicose by cross-linked recombinant L-rhamnose isomerase: Separation of product by ethanol crystallization. J. Biosci. Bioeng. 2006, 101, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.N.; Shin, K.C.; Kim, D.W.; Kim, B.J.; Park, C.S.; Yeom, S.J.; Kim, Y.S. Production of D-Allose From D-Allulose Using Commercial Immobilized Glucose Isomerase. Front. Bioeng. Biotechnol. 2021, 9, 681253. [Google Scholar] [CrossRef]
- Young, J.M.; Schray, K.J.; Mildvan, A.S. Proton magnetic relaxation studies of the interaction of D-xylose and xylitol with D-xylose isomerase. Characterization of metal-enzyme-substrate interactions. J. Biol. Chem. 1975, 250, 9021–9027. [Google Scholar] [CrossRef]
- Yoshimura, S.; Danno, G.-i.; Natake, M. Studies ond-Glucose Isomerizing Activity ofd-Xylose Grown Cells from Bacillus coagulans, Strain HN-68. Agric. Biol. Chem. 2014, 30, 1015–1023. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Rozeboom, H.J.; de Waal, P.P.; de Jong, R.M.; Dudek, H.M.; Janssen, D.B. Metal Dependence of the Xylose Isomerase from Piromyces sp. E2 Explored by Activity Profiling and Protein Crystallography. Biochemistry 2017, 56, 5991–6005. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, Y. Studies on Sugar-isomerizing Enzyme. Agric. Biol. Chem. 2014, 30, 1247–1253. [Google Scholar] [CrossRef]
- Chanitnun, K.; Pinphanichakarn, P. Glucose(xylose) isomerase production by Streptomyces sp. CH7 grown on agricultural residues. Braz. J. Microbiol. 2012, 43, 1084–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalevsky, A.Y.; Hanson, L.; Fisher, S.Z.; Mustyakimov, M.; Mason, S.A.; Forsyth, V.T.; Blakeley, M.P.; Keen, D.A.; Wagner, T.; Carrell, H.L.; et al. Metal ion roles and the movement of hydrogen during reaction catalyzed by D-xylose isomerase: A joint x-ray and neutron diffraction study. Structure 2010, 18, 688–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.Y.; Bagdasarian, M.; Meng, M.H.; Zeikus, J.G. Catalytic mechanism of xylose (glucose) isomerase from Clostridium thermosulfurogenes. Characterization of the structural gene and function of active site histidine. J. Biol. Chem. 1990, 265, 19082–19090. [Google Scholar] [CrossRef]
- Collyer, C.A.; Henrick, K.; Blow, D.M. Mechanism for aldose-ketose interconversion by D-xylose isomerase involving ring opening followed by a 1,2-hydride shift. J. Mol. Biol. 1990, 212, 211–235. [Google Scholar] [CrossRef]
- Collyer, C.A.; Blow, D.M. Observations of reaction intermediates and the mechanism of aldose-ketose interconversion by D-xylose isomerase. Proc. Natl. Acad. Sci. USA 1990, 87, 1362–1366. [Google Scholar] [CrossRef] [Green Version]
- Carrell, H.L.; Rubin, B.H.; Hurley, T.J.; Glusker, J.P. X-ray crystal structure of D-xylose isomerase at 4 Å resolution. J. Biol. Chem. 1984, 259, 3230–3236. [Google Scholar] [CrossRef]
- Taberman, H.; Bury, C.S.; van der Woerd, M.J.; Snell, E.H.; Garman, E.F. Structural knowledge or X-ray damage? A case study on xylose isomerase illustrating both. J. Synchrotron Radiat. 2019, 26, 931–944. [Google Scholar] [CrossRef]
- Lee, D.; Baek, S.; Park, J.; Lee, K.; Kim, J.; Lee, S.J.; Chung, W.K.; Lee, J.L.; Cho, Y.; Nam, K.H. Nylon mesh-based sample holder for fixed-target serial femtosecond crystallography. Sci. Rep. 2019, 9, 6971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Lee, D.; Baek, S.; Park, J.; Lee, S.J.; Park, S.; Chung, W.K.; Lee, J.L.; Cho, H.S.; Cho, Y.; et al. Viscous-medium-based crystal support in a sample holder for fixed-target serial femtosecond crystallography. J. Appl. Crystallogr. 2020, 53, 1051–1059. [Google Scholar] [CrossRef]
- Nam, K.H. Stable sample delivery in viscous media via a capillary for serial crystallography. J. Appl. Crystallogr. 2020, 53, 45–50. [Google Scholar] [CrossRef]
- Nam, K.H. Shortening injection matrix for serial crystallography. Sci. Rep. 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.H. Polysaccharide-Based Injection Matrix for Serial Crystallography. Int. J. Mol. Sci. 2020, 21, 3332. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.H. Lard Injection Matrix for Serial Crystallography. Int. J. Mol. Sci. 2020, 21, 5977. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Choi, H.; Eo, C.; Cho, Y.; Nam, K.H. Fixed-Target Serial Synchrotron Crystallography Using Nylon Mesh and Enclosed Film-Based Sample Holder. Crystals 2020, 10, 803. [Google Scholar] [CrossRef]
- Dauter, Z.; Dauter, M.; Hemker, J.; Witzel, H.; Wilson, K.S. Crystallisation and preliminary analysis of glucose isomerase from Streptomyces albus. FEBS Lett. 1989, 247, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Whitlow, M.; Howard, A.J.; Finzel, B.C.; Poulos, T.L.; Winborne, E.; Gilliland, G.L. A metal-mediated hydride shift mechanism for xylose isomerase based on the 1.6 Å Streptomyces rubiginosus structures with xylitol and D-xylose. Proteins 1991, 9, 153–173. [Google Scholar] [CrossRef]
- Carrell, H.L.; Glusker, J.P.; Burger, V.; Manfre, F.; Tritsch, D.; Biellmann, J.F. X-ray analysis of D-xylose isomerase at 1.9 A: Native enzyme in complex with substrate and with a mechanism-designed inactivator. Proc. Natl. Acad. Sci. USA 1989, 86, 4440–4444. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.E.; Kim, I.J.; Nam, K.H. Crystal structure of glucose isomerase in complex with xylitol inhibitor in one metal binding mode. Biochem. Biophys. Res. Commun. 2017, 493, 666–670. [Google Scholar] [CrossRef]
- Park, S.H.; Kwon, S.; Lee, C.W.; Kim, C.M.; Jeong, C.S.; Kim, K.J.; Hong, J.W.; Kim, H.J.; Park, H.H.; Lee, J.H. Crystal Structure and Functional Characterization of a Xylose Isomerase (PbXI) from the Psychrophilic Soil Microorganism, Paenibacillus sp. J. Microbiol. Biotechnol. 2019, 29, 244–255. [Google Scholar] [CrossRef]
- Lee, M.; Rozeboom, H.J.; Keuning, E.; de Waal, P.; Janssen, D.B. Structure-based directed evolution improves S. cerevisiae growth on xylose by influencing in vivo enzyme performance. Biotechnol. Biofuels 2020, 13, 5. [Google Scholar] [CrossRef]
- Sugahara, M.; Mizohata, E.; Nango, E.; Suzuki, M.; Tanaka, T.; Masuda, T.; Tanaka, R.; Shimamura, T.; Tanaka, Y.; Suno, C.; et al. Grease matrix as a versatile carrier of proteins for serial crystallography. Nat. Methods 2015, 12, 61–63. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H. Room-Temperature Structure of Xylitol-Bound Glucose Isomerase by Serial Crystallography: Xylitol Binding in the M1 Site Induces Release of Metal Bound in the M2 Site. Int. J. Mol. Sci. 2021, 22, 3892. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Metoz, F. ESPript: Analysis of multiple sequence alignments in PostScript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H. Crystal structure of the metal-free state of glucose isomerase reveals its minimal open configuration for metal binding. Biochem. Biophys. Res. Commun. 2021, 547, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Sudfeldt, C.; Schaffer, A.; Kagi, J.H.; Bogumil, R.; Schulz, H.P.; Wulff, S.; Witzel, H. Spectroscopic studies on the metal-ion-binding sites of Co2+-substituted D-xylose isomerase from Streptomyces rubiginosus. Eur. J. Biochem. 1990, 193, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Callens, M.; Kerstershilderson, H.; Vangrysperre, W.; Debruyne, C.K. D-Xylose Isomerase from Streptomyces violaceoruber: Structural and Catalytic Roles of Bivalent-Metal Ions. Enzym. Microb. Technol. 1988, 10, 695–700. [Google Scholar] [CrossRef]
- Smith, C.A.; Rangarajan, M.; Hartley, B.S. D-Xylose (D-glucose) isomerase from Arthrobacter strain N.R.R.L. B3728. Purification and properties. Biochem. J. 1991, 277, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenn, T.D.; Ringe, D.; Petsko, G.A. Xylose isomerase in substrate and inhibitor michaelis states: Atomic resolution studies of a metal-mediated hydride shift. Biochemistry 2004, 43, 6464–6474. [Google Scholar] [CrossRef] [PubMed]
- Volkin, D.B.; Klibanov, A.M. Mechanism of thermoinactivation of immobilized glucose isomerase. Biotechnol. Bioeng. 1989, 33, 1104–1111. [Google Scholar] [CrossRef]
- Carrell, H.L.; Hoier, H.; Glusker, J.P. Modes of binding substrates and their analogues to the enzyme D-xylose isomerase. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 113–123. [Google Scholar] [CrossRef]
- Kovalevsky, A.; Hanson, B.L.; Mason, S.A.; Forsyth, V.T.; Fisher, Z.; Mustyakimov, M.; Blakeley, M.P.; Keen, D.A.; Langan, P. Inhibition of D-xylose isomerase by polyols: Atomic details by joint X-ray/neutron crystallography. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 1201–1206. [Google Scholar] [CrossRef]
- White, J.S. Straight talk about high-fructose corn syrup: What it is and what it ain’t. Am. J. Clin. Nutr. 2008, 88, 1716S–1721S. [Google Scholar] [CrossRef]
- Sahin, A.W.; Zannini, E.; Coffey, A.; Arendt, E.K. Sugar reduction in bakery products: Current strategies and sourdough technology as a potential novel approach. Food Res. Int. 2019, 126, 108583. [Google Scholar] [CrossRef]
- Zargaraan, A.; Kamaliroosta, L.; Yaghoubi, A.S.; Mirmoghtadaie, L. Effect of Substitution of Sugar by High Fructose Corn Syrup on the Physicochemical Properties of Bakery and Dairy Products: A Review. Nutr. Food Sci. 2016, 3, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K.; Singh, S.V. Osmotic dehydration of fruits and vegetables: A review. J. Food Sci. Technol. 2014, 51, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Basciano, H.; Federico, L.; Adeli, K. Fructose, insulin resistance, and metabolic dyslipidemia. Nutr. Metab. 2005, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Bhasin, S.; Modi, H.A. Optimization of Fermentation Medium for the Production of Glucose Isomerase Using Streptomyces sp. SB-P1. Biotechnol. Res. Int. 2012, 2012, 874152. [Google Scholar] [CrossRef] [Green Version]
- Serna-Saldivar, S.O. Maize: Foods from Maize. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Zheng, W.; Huang, J.F.; Jin, L.Q.; Jia, D.X.; Zhou, H.Y.; Xu, J.M.; Liao, C.J.; Cheng, X.P.; Mao, B.X.; et al. Improvement and characterization of a hyperthermophilic glucose isomerase from Thermoanaerobacter ethanolicus and its application in production of high fructose corn syrup. J. Ind. Microbiol. Biotechnol. 2015, 42, 1091–1103. [Google Scholar] [CrossRef]
- Singh, R.S.; Chauhan, K.; Singh, R.P. Enzymatic Approaches for the Synthesis of High Fructose Syrup. In Plant Biotechnology: Recent Advancements and Developments; Springer: Singapore, 2017; pp. 189–211. [Google Scholar] [CrossRef]
- Dai, C.; Miao, T.; Hai, J.; Xiao, Y.; Li, Y.; Zhao, J.; Qiu, H.; Xu, B. A Novel Glucose Isomerase from Caldicellulosiruptor bescii with Great Potentials in the Production of High-Fructose Corn Syrup. Biomed Res. Int. 2020, 2020, 1871934. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.H.; Sjoholm, C.; Kelly, R.M. Purification and characterization of a highly thermostable glucose isomerase produced by the extremely thermophilic eubacterium, Thermotoga maritima. Biotechnol. Bioeng. 1993, 41, 878–886. [Google Scholar] [CrossRef]
- Deng, H.; Chen, S.; Wu, D.; Chen, J.; Wu, J. Heterologous expression and biochemical characterization of glucose isomerase from Thermobifida fusca. Bioprocess Biosyst. Eng. 2014, 37, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.X.; Zhou, L.; Zheng, Y.G. Properties of a novel thermostable glucose isomerase mined from Thermus oshimai and its application to preparation of high fructose corn syrup. Enzym. Microb. Technol. 2017, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vieille, C.; Hess, J.M.; Kelly, R.M.; Zeikus, J.G. xylA cloning and sequencing and biochemical characterization of xylose isomerase from Thermotoga neapolitana. Appl. Environ. Microbiol. 1995, 61, 1867–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canilha, L.; Kumar Chandel, A.; dos Santos Milessi, T.S.; Fernandes Antunes, F.A.; da Costa Freitas, W.L.; das Gracas Almeida Felipe, M.; da Silva, S.S. Bioconversion of sugarcane biomass into ethanol: An overview about composition, pretreatment methods, detoxification of hydrolysates, enzymatic saccharification, and ethanol fermentation. J. Biomed. Biotechnol. 2012, 2012, 989572. [Google Scholar] [CrossRef] [PubMed]
- Chiang, L.C.; Gong, C.S.; Chen, L.F.; Tsao, G.T. d-Xylulose Fermentation to Ethanol by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1981, 42, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.S.; Chen, L.F.; Flickinger, M.C.; Chiang, L.C.; Tsao, G.T. Production of Ethanol from d-Xylose by Using d-Xylose Isomerase and Yeasts. Appl. Environ. Microbiol. 1981, 41, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.K.; Um, Y.; Woo, H.M.; Kim, K.H.; Lee, S.M. Ethanol production from lignocellulosic hydrolysates using engineered Saccharomyces cerevisiae harboring xylose isomerase-based pathway. Bioresour. Technol. 2016, 209, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Liu, Y.; Qian, F.; Yang, J.; Jiang, Y.; Yang, S. Construction of fast xylose-fermenting yeast based on industrial ethanol-producing diploid Saccharomyces cerevisiae by rational design and adaptive evolution. BMC Biotechnol. 2013, 13, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeke, M.M.; Dietz, H.; Li, Y.; Foulquie-Moreno, M.R.; Mutturi, S.; Deprez, S.; Den Abt, T.; Bonini, B.M.; Liden, G.; Dumortier, F.; et al. Development of a D-xylose fermenting and inhibitor tolerant industrial Saccharomyces cerevisiae strain with high performance in lignocellulose hydrolysates using metabolic and evolutionary engineering. Biotechnol. Biofuels 2013, 6, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seike, T.; Kobayashi, Y.; Sahara, T.; Ohgiya, S.; Kamagata, Y.; Fujimori, K.E. Molecular evolutionary engineering of xylose isomerase to improve its catalytic activity and performance of micro-aerobic glucose/xylose co-fermentation in Saccharomyces cerevisiae. Biotechnol. Biofuels 2019, 12, 139. [Google Scholar] [CrossRef]
- Bracher, J.M.; Martinez-Rodriguez, O.A.; Dekker, W.J.C.; Verhoeven, M.D.; van Maris, A.J.A.; Pronk, J.T. Reassessment of requirements for anaerobic xylose fermentation by engineered, non-evolved Saccharomyces cerevisiae strains. FEMS Yeast Res. 2019, 19, foy104. [Google Scholar] [CrossRef]
- Tran Nguyen Hoang, P.; Ko, J.K.; Gong, G.; Um, Y.; Lee, S.M. Genomic and phenotypic characterization of a refactored xylose-utilizing Saccharomyces cerevisiae strain for lignocellulosic biofuel production. Biotechnol. Biofuels 2018, 11, 268. [Google Scholar] [CrossRef] [Green Version]
- Vilela Lde, F.; de Araujo, V.P.; Paredes Rde, S.; Bon, E.P.; Torres, F.A.; Neves, B.C.; Eleutherio, E.C. Enhanced xylose fermentation and ethanol production by engineered Saccharomyces cerevisiae strain. AMB Express 2015, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Tanino, T.; Hotta, A.; Ito, T.; Ishii, J.; Yamada, R.; Hasunuma, T.; Ogino, C.; Ohmura, N.; Ohshima, T.; Kondo, A. Construction of a xylose-metabolizing yeast by genome integration of xylose isomerase gene and investigation of the effect of xylitol on fermentation. Appl. Microbiol. Biotechnol. 2010, 88, 1215–1221. [Google Scholar] [CrossRef]
- Kuyper, M.; Harhangi, H.R.; Stave, A.K.; Winkler, A.A.; Jetten, M.S.; de Laat, W.T.; den Ridder, J.J.; Op den Camp, H.J.; van Dijken, J.P.; Pronk, J.T. High-level functional expression of a fungal xylose isomerase: The key to efficient ethanolic fermentation of xylose by Saccharomyces cerevisiae? FEMS Yeast Res. 2003, 4, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Hector, R.E.; Dien, B.S.; Cotta, M.A.; Mertens, J.A. Growth and fermentation of D-xylose by Saccharomyces cerevisiae expressing a novel D-xylose isomerase originating from the bacterium Prevotella ruminicola TC2-24. Biotechnol. Biofuels 2013, 6, 84. [Google Scholar] [CrossRef] [Green Version]
- Harcus, D.; Dignard, D.; Lepine, G.; Askew, C.; Raymond, M.; Whiteway, M.; Wu, C. Comparative xylose metabolism among the Ascomycetes C. albicans, S. stipitis and S. cerevisiae. PLoS ONE 2013, 8, e80733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aeling, K.A.; Salmon, K.A.; Laplaza, J.M.; Li, L.; Headman, J.R.; Hutagalung, A.H.; Picataggio, S. Co-fermentation of xylose and cellobiose by an engineered Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2012, 39, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Waltman, M.J.; Yang, Z.K.; Langan, P.; Graham, D.E.; Kovalevsky, A. Engineering acidic Streptomyces rubiginosus D-xylose isomerase by rational enzyme design. Protein Eng. Des. Sel. 2014, 27, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.M.; Jellison, T.; Alper, H.S. Directed evolution of xylose isomerase for improved xylose catabolism and fermentation in the yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 5708–5716. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, K.H. Glucose Isomerase: Functions, Structures, and Applications. Appl. Sci. 2022, 12, 428. https://doi.org/10.3390/app12010428
Nam KH. Glucose Isomerase: Functions, Structures, and Applications. Applied Sciences. 2022; 12(1):428. https://doi.org/10.3390/app12010428
Chicago/Turabian StyleNam, Ki Hyun. 2022. "Glucose Isomerase: Functions, Structures, and Applications" Applied Sciences 12, no. 1: 428. https://doi.org/10.3390/app12010428
APA StyleNam, K. H. (2022). Glucose Isomerase: Functions, Structures, and Applications. Applied Sciences, 12(1), 428. https://doi.org/10.3390/app12010428