
Lactoplantibacillus plantarum KG Lysates Inhibit the Internalization of Staphylococcus aureus by Human Keratinocytes through the Induction of Human Beta-Defensin 3


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. Bacterial Lysate Preparation
2.3. Cell Viability Assay
2.4. Real-Time PCR
2.5. Western Blot Analysis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Preparation of S. aureus
2.8. Internalization Assay
2.9. Preparation of Conditioned Media (CM)
2.10. Statistical Analysis
3. Results
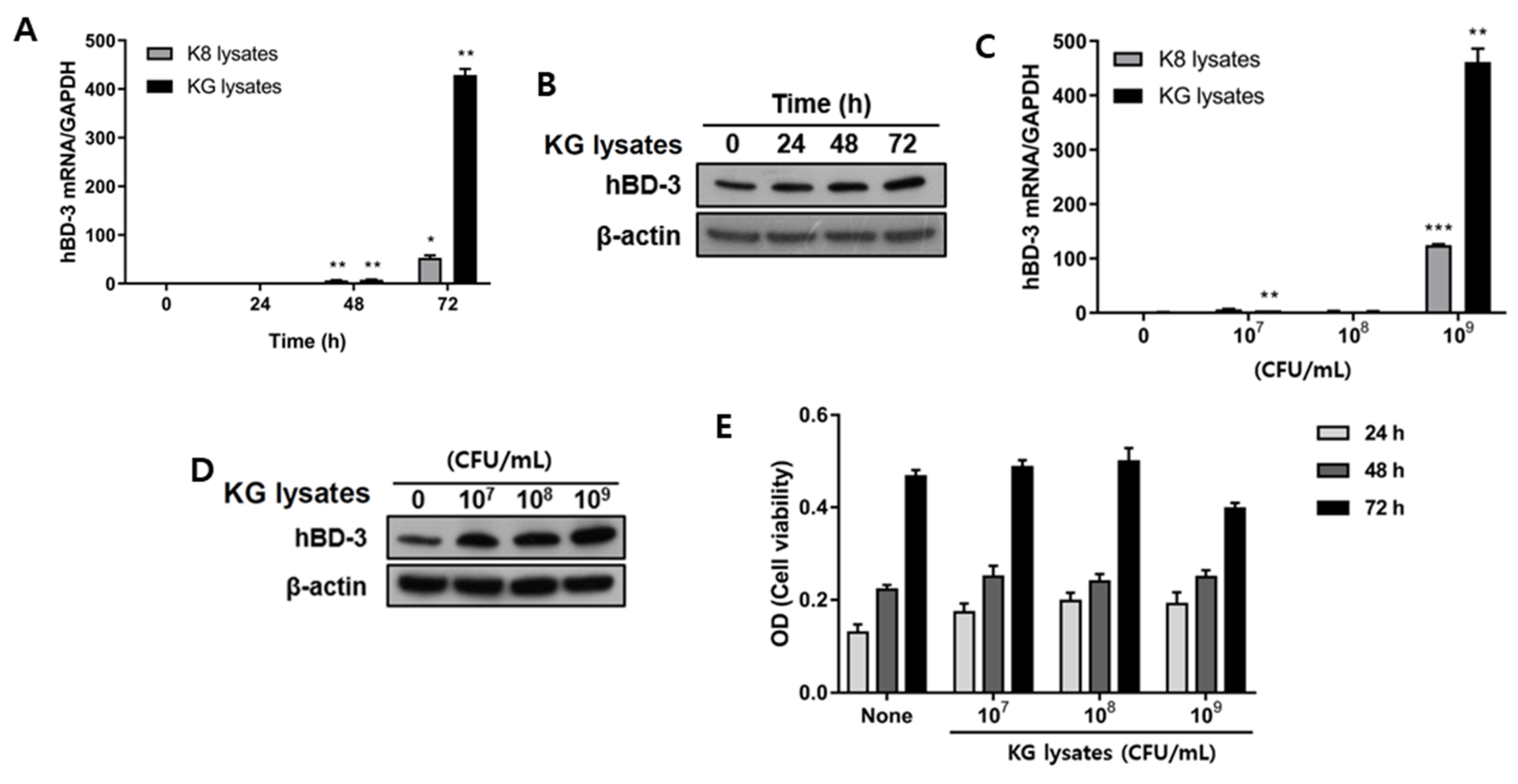
3.1. L. plantarum KG Lysates Significantly Increased hBD3 Production in HaCaT Cells
3.2. L. plantarum KG Lysates Inhibited S. aureus Internalization into HaCaT Cells via hBD3 Induction
3.3. JAK-STAT Signaling Pathway Was Involved in hBD3 Induction in HaCaT Cells
3.4. L. plantarum KG Lysates Increased EGF and EGFR Expression in HaCaT Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoover, D.M.; Wu, Z.; Tucker, K.; Lu, W.; Lubkowski, J. Antimicrobial characterization of human beta-defensin 3 derivatives. Antimicrob. Agents Chemother. 2003, 47, 2804–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewary, P.; Li, Z.; de la Rosa, G.; Chen, Q.; Oppenheim, J.; Yang, D. Human Beta Defensin 3 (HBD3) induces migration and activation of antigen presenting cells and acts as an immune enhancer. FASEB J. 2008, 22, 673. [Google Scholar] [CrossRef]
- Bayer, A.; Lammel, J.; Tohidnezhad, M.; Lippross, S.; Behrendt, P.; Klüter, T.; Pufe, T.; Cremer, J.; Jahr, H.; Rademacher, F.; et al. The Antimicrobial Peptide Human Beta-Defensin-3 Is Induced by Platelet-Released Growth Factors in Primary Keratinocytes. Mediat. Inflamm. 2017, 2017, 6157491. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, O.E.; Cowland, J.B.; Theilgaard-Mönch, K.; Liu, L.; Ganz, T.; Borregaard, N. Wound healing and expression of antimicrobial peptides/polypeptides in human keratinocytes, a consequence of common growth factors. J. Immunol. 2003, 170, 5583–5589. [Google Scholar] [CrossRef] [Green Version]
- Kiehne, K.; Fincke, A.; Brunke, G.; Lange, T.; Fölsch, U.R.; Herzig, K.H. Antimicrobial peptides in chronic anal fistula epithelium. Scand. J. Gastroenterol. 2007, 42, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, O.E.; Thapa, D.R.; Rosenthal, A.; Liu, L.; Roberts, A.A.; Ganz, T. Differential regulation of beta-defensin expression in human skin by microbial stimuli. J. Immunol. 2005, 174, 4870–4879. [Google Scholar] [CrossRef] [Green Version]
- Wehkamp, J.; Harder, J.; Wehkamp, K.; Wehkamp-von Meissner, B.; Schlee, M.; Enders, C.; Sonnenborn, U.; Nuding, S.; Bengmark, S.; Fellermann, K.; et al. NF-kappaB- and AP-1-mediated induction of human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle 1917: A novel effect of a probiotic bacterium. Infect. Immun. 2004, 72, 5750–5758. [Google Scholar] [CrossRef] [Green Version]
- Paolillo, R.; Romano Carratelli, C.; Sorrentino, S.; Mazzola, N.; Rizzo, A. Immunomodulatory effects of Lactobacillus plantarum on human colon cancer cells. Int. Immunopharmacol. 2009, 9, 1265–1271. [Google Scholar] [CrossRef]
- Kobatake, E.; Kabuki, T. S-Layer Protein of Lactobacillus helveticus SBT2171 Promotes Human β-Defensin 2 Expression via TLR2-JNK Signaling. Front. Microbiol. 2019, 10, 2414. [Google Scholar] [CrossRef]
- Liu, A.Y.; Destoumieux, D.; Wong, A.V.; Park, C.H.; Valore, E.V.; Liu, L.; Ganz, T. Human beta-defensin-2 production in keratinocytes is regulated by interleukin-1, bacteria, and the state of differentiation. J. invest. Dermatol. 2002, 118, 275–281. [Google Scholar] [CrossRef]
- Lee, D.E.; Huh, C.S.; Ra, J.; Choi, I.D.; Jeong, J.W.; Kim, S.H.; Ryu, J.H.; Seo, Y.K.; Koh, J.S.; Lee, J.H.; et al. Clinical Evidence of Effects of Lactobacillus plantarum HY7714 on Skin Aging: A Randomized, Double Blind, Placebo-Controlled Study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Kim, S.A.; Park, S.D.; Kim, H.J.; Kim, J.S.; Bae, C.H.; Kim, J.Y.; Nam, W.; Lee, J.L.; Sim, J.H. Regulatory effects of Lactobacillus plantarum HY7714 on skin health by improving intestinal condition. PLoS ONE 2020, 15, e0231268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.H.; Chou, C.H.; Chiang, Y.J.; Lin, C.G.; Lee, C.H. Regulatory effects of Lactobacillus plantarum-GMNL6 on human skin health by improving skin microbiome. Int. J. Med. Sci. 2021, 18, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.; Jung, B.J.; Kim, N.R.; Park, J.E.; Chung, D.K. Lipoteichoic acid isolated from Lactobacillus plantarum suppresses LPS-mediated atherosclerotic plaque inflammation. Mol. Cells 2013, 35, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.F.; Kim, H.; Kim, H.R.; Gim, M.G.; Chung, D.K. Different immune regulatory potential of Lactobacillus plantarum and Lactobacillus sakei isolated from Kimchi. J. Microbiol. Biotechnol. 2014, 24, 1629–1635. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, H.R.; Kim, N.R.; Jeong, B.J.; Lee, J.S.; Jang, S.; Chung, D.K. Oral administration of Lactobacillus plantarum lysates attenuates the development of atopic dermatitis lesions in mouse models. J. Microbiol. 2015, 53, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Hong, Y.F.; Jeon, B.; Jung, B.J.; Chung, D.K.; Kim, H. Differential Cytokine Regulatory Effect of Three Lactobacillus Strains Isolated from Fermented Foods. J. Microbiol. Biotechnol. 2016, 26, 1517–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.E.; Kim, H.; Chung, D.K. Lipoteichoic Acid Isolated from Lactobacillus plantarum Maintains Inflammatory Homeostasis through Regulation of Th1- and Th2-Induced Cytokines. J. Microbiol. Biotechnol. 2019, 29, 151–159. [Google Scholar] [CrossRef]
- Gambichler, T.; Skrygan, M.; Tomi, N.S.; Othlinghaus, N.; Brockmeyer, N.H.; Altmeyer, P.; Kreuter, A. Differential mRNA expression of antimicrobial peptides and proteins in atopic dermatitis as compared to psoriasis vulgaris and healthy skin. Int. Arch. Allergy Immunol. 2008, 147, 17–24. [Google Scholar] [CrossRef]
- Wang, J.N.; Li, M. The Immune Function of Keratinocytes in Anti-Pathogen Infection in the Skin. Int. J. Dermatol. Venereol. 2020, 3, 231–238. [Google Scholar] [CrossRef]
- Schittek, B.; Paulmann, M.; Senyürek, I.; Steffen, H. The role of antimicrobial peptides in human skin and in skin infectious diseases. Infect. Disord. Drug Targets 2008, 8, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuyi, Y.; Feng, W.; Jing, T.; Hongzhang, H.; Haiyan, W.; Pingping, M.; Liwu, Z.; Zwahlen, R.A.; Hongyu, Y. Human beta-defensin-3 (hBD-3) upregulated by LPS via epidermal growth factor receptor (EGFR) signaling pathways to enhance lymphatic invasion of oral squamous cell carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2011, 112, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Pedersen, M.W.; Woetmann, A.; Villingshøj, M.; Stockhausen, M.T.; Odum, N.; Poulsen, H.S. EGFR induces expression of IRF-1 via STAT1 and STAT3 activation leading to growth arrest of human cancer cells. Int. J. Cancer 2008, 122, 342–349. [Google Scholar] [CrossRef]
- Lulli, D.; Carbone, M.; Pastore, S. Epidermal growth factor receptor inhibitors trigger a type I interferon response in human skin. Oncotarget 2016, 7, 47777–47793. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Zhang, Y.B.; Gui, J.F.; Lemon, S.M.; Yamane, D. Interferon regulatory factor 1 (IRF1) and anti-pathogen innate immune responses. PLoS Pathog. 2021, 17, e1009220. [Google Scholar] [CrossRef]
- Jang, K.S.; Baik, J.E.; Han, S.H.; Chung, D.K.; Kim, B.G. Multi-spectrometric analyses of lipoteichoic acids isolated from Lactobacillus plantarum. Biochem. Biophys. Res. Commun. 2011, 407, 823–830. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, K.-O.; Yu, C.; Kim, H.; Chung, D.-K. Lactoplantibacillus plantarum KG Lysates Inhibit the Internalization of Staphylococcus aureus by Human Keratinocytes through the Induction of Human Beta-Defensin 3. Appl. Sci. 2022, 12, 12504. https://doi.org/10.3390/app122312504
Jang K-O, Yu C, Kim H, Chung D-K. Lactoplantibacillus plantarum KG Lysates Inhibit the Internalization of Staphylococcus aureus by Human Keratinocytes through the Induction of Human Beta-Defensin 3. Applied Sciences. 2022; 12(23):12504. https://doi.org/10.3390/app122312504
Chicago/Turabian StyleJang, Kyoung-Ok, Chaeyeon Yu, Hangeun Kim, and Dae-Kyun Chung. 2022. "Lactoplantibacillus plantarum KG Lysates Inhibit the Internalization of Staphylococcus aureus by Human Keratinocytes through the Induction of Human Beta-Defensin 3" Applied Sciences 12, no. 23: 12504. https://doi.org/10.3390/app122312504
APA StyleJang, K. -O., Yu, C., Kim, H., & Chung, D. -K. (2022). Lactoplantibacillus plantarum KG Lysates Inhibit the Internalization of Staphylococcus aureus by Human Keratinocytes through the Induction of Human Beta-Defensin 3. Applied Sciences, 12(23), 12504. https://doi.org/10.3390/app122312504