
Bacterial Cellulose Production by Komagateibacter xylinus with the Use of Enzyme-Degraded Oligo- and Polysaccharides as the Substrates

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
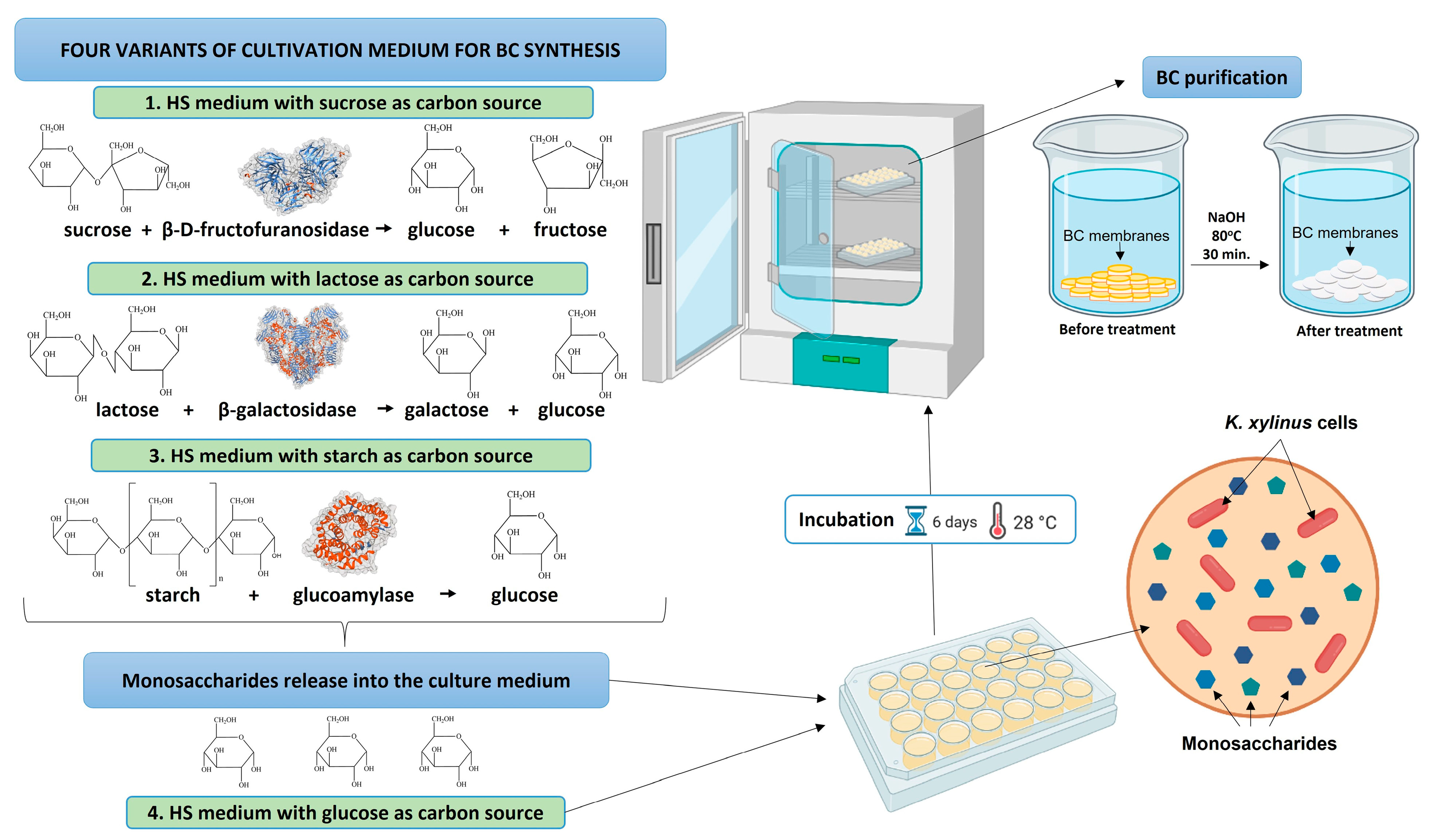
2.2. Preparation of the Culture Growth Media
2.3. Bacterial Cellulose Synthesis and Purification
2.4. Analysis of the Parameters of the Pre- and Post-Culture
2.4.1. pH Determination
2.4.2. Glucose Concentration Determination
2.4.3. Enzymes Activity Determination
2.5. Bacterial Cellulose Synthesis Efficiency and Yield Determination
2.6. Moisture Content Ratio and Swelling Ratio
2.7. BC Structural Properties Analysis
2.7.1. Attenuated Total Reflectance in Fourier Transformation InfraRed Spectroscopy (ATR-FTIR) Analysis
2.7.2. X-ray Diffraction Analysis (XRD)
2.7.3. Scanning Electron Microscope Analysis (SEM)
2.8. Statistical Analysis
3. Results and Discussion
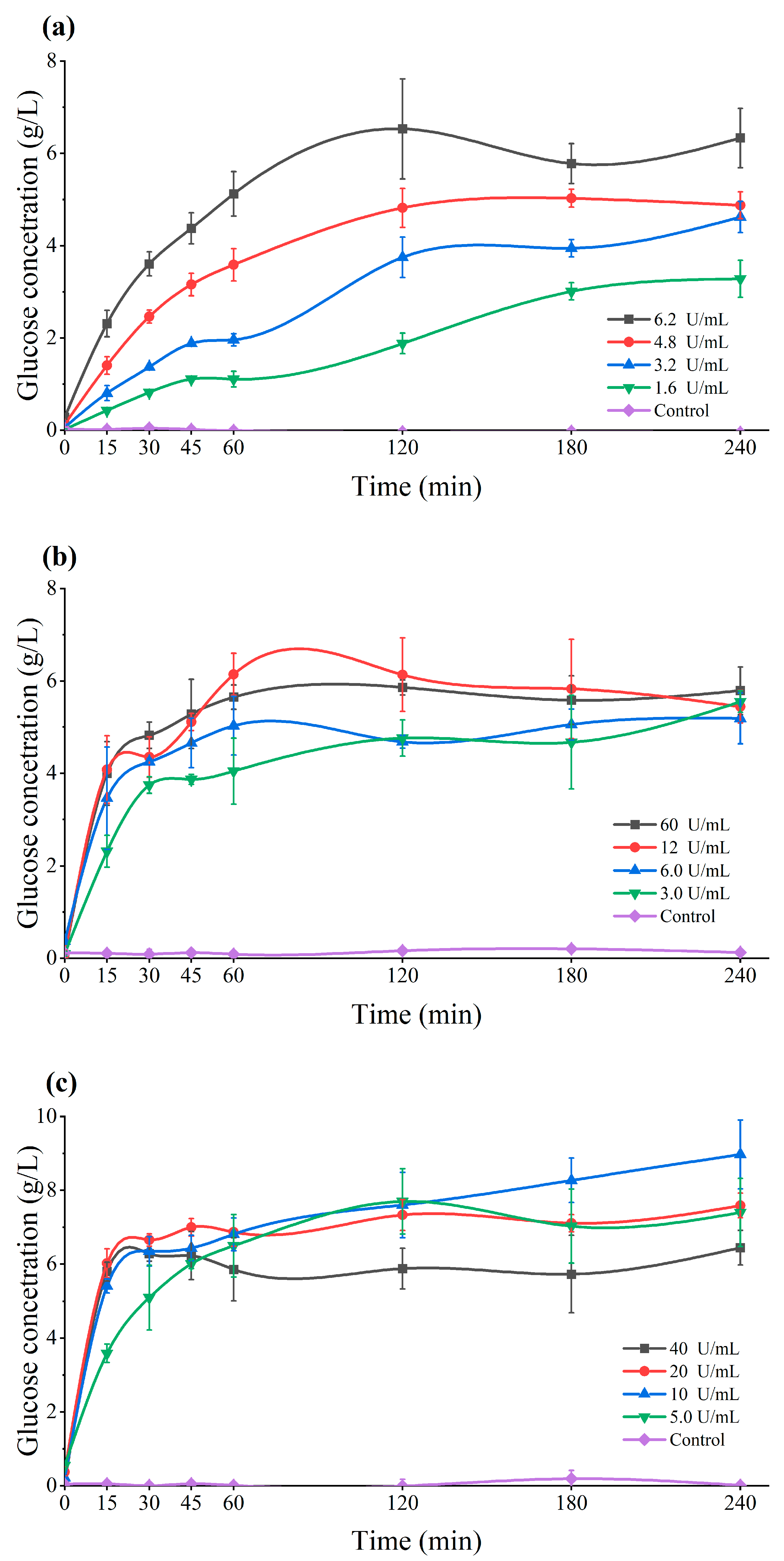
3.1. The Influence of Cultivation Media on Substrate Hydrolysis and Enzymes Stability
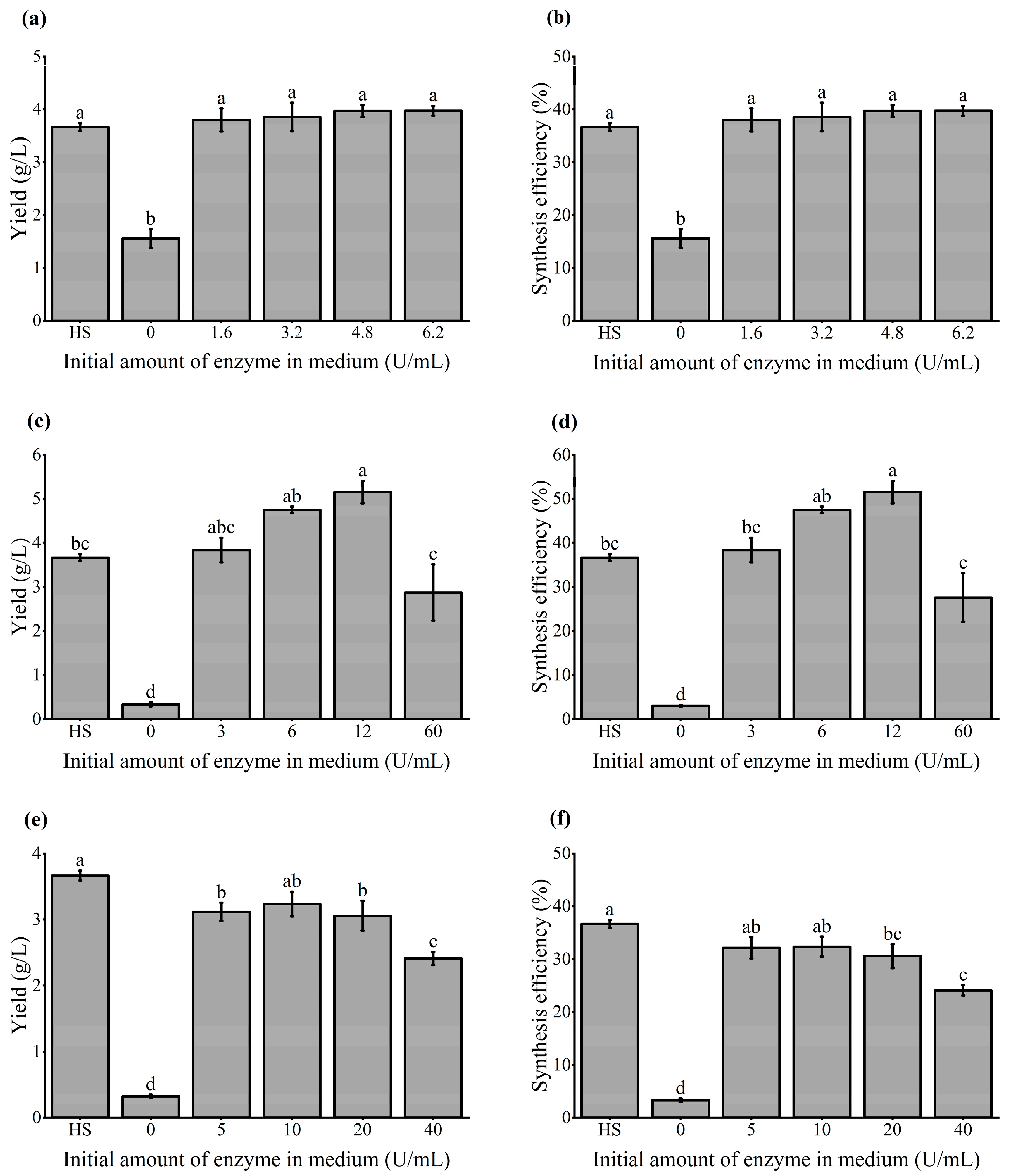
3.2. Yield and Synthesis Efficiency of BC
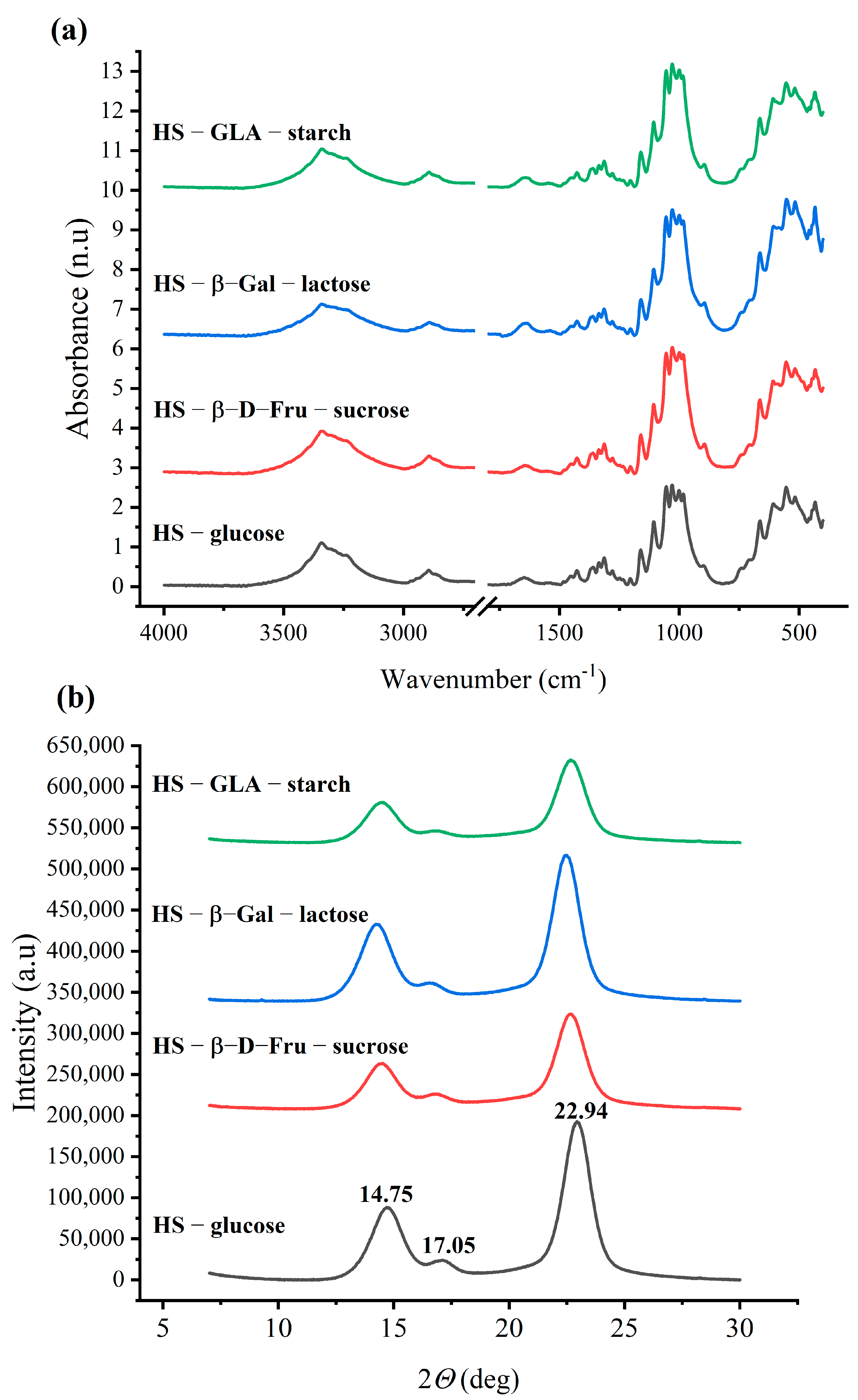
3.3. Attenuated Total Reflectance in Fourier Transformation InfraRed Spectroscopy (ATR-FTIR) and X-ray Diffraction (XRD) Analysis of Bacterial Cellulose
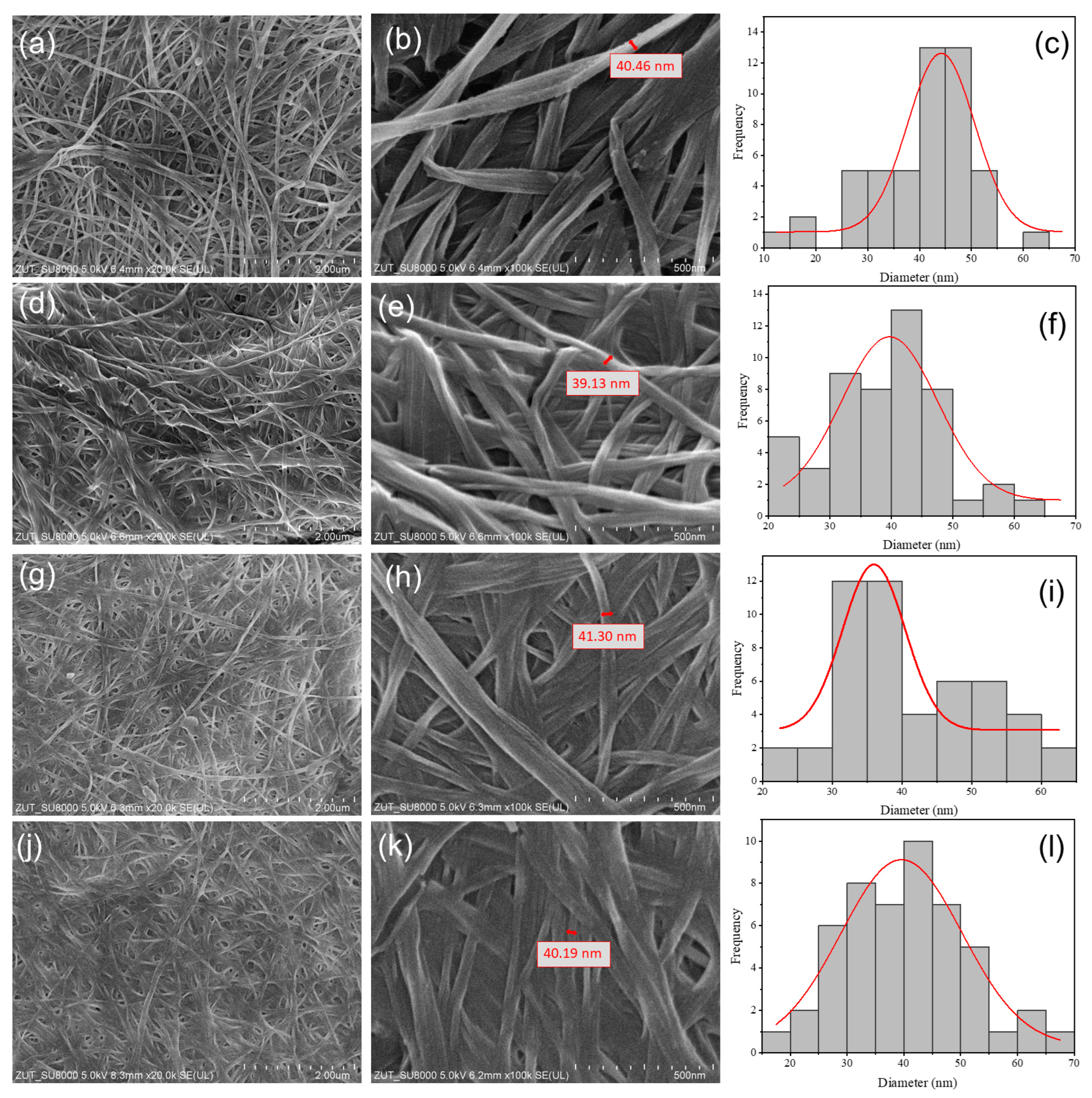
3.4. Scanning Electron Microscope (SEM) Analysis
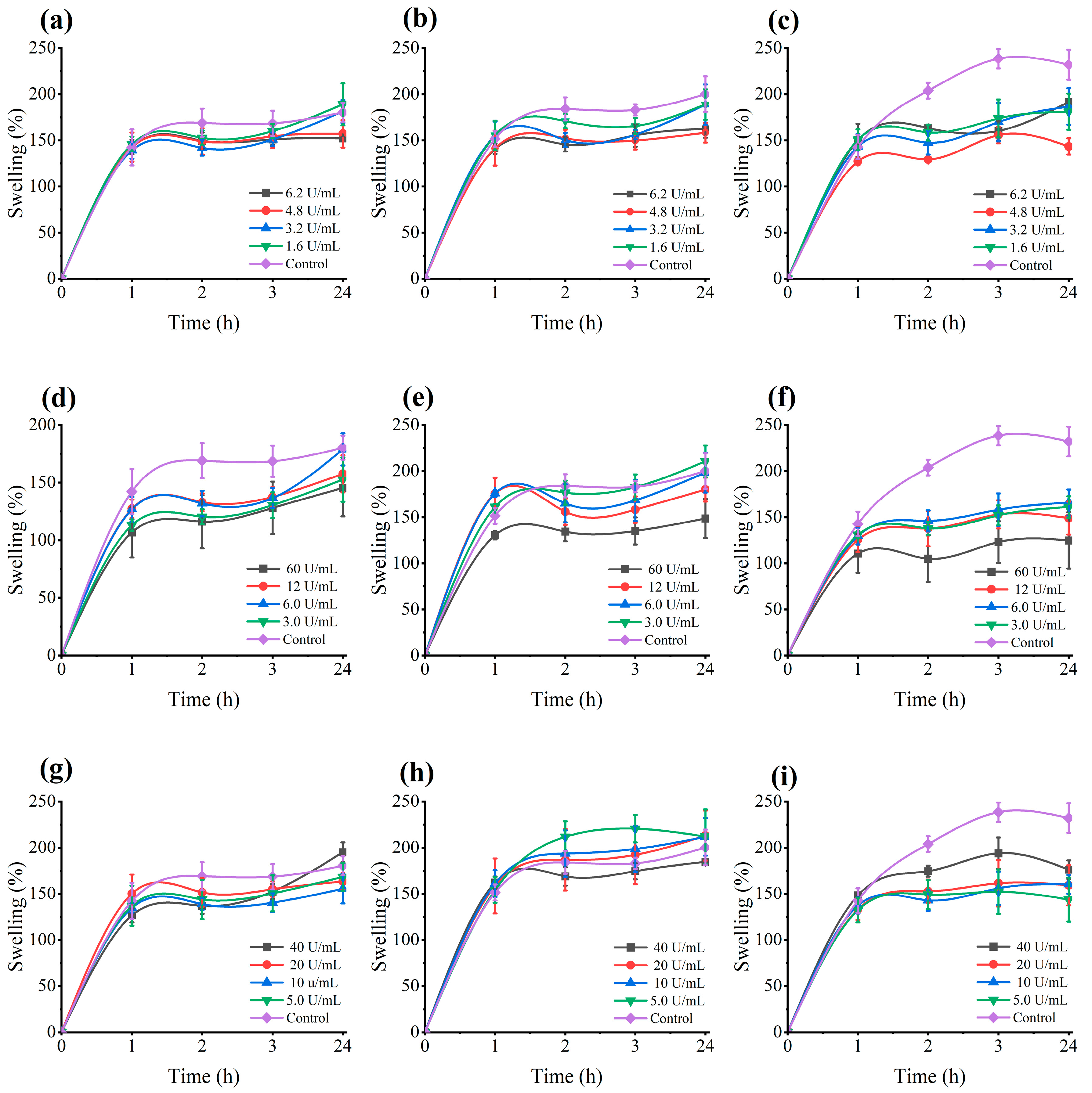
3.5. Moisture Content and Swelling Ratios
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, W.; Du, H.; Zhang, M.; Liu, K.; Liu, H.; Xie, H.; Zhang, X.; Si, C. Bacterial cellulose-based composite scaffolds for biomedical applications: A review. ACS Sustain. Chem. Eng. 2020, 8, 7536–7562. [Google Scholar] [CrossRef]
- Ahmed, J.; Gultekinoglu, M.; Edirisinghe, M. Bacterial cellulose micro-nano fibres for wound healing applications. Biotechnol. Adv. 2020, 41, 107549. [Google Scholar] [CrossRef] [PubMed]
- Bodea, I.M.; Cătunescu, G.M.; Stroe, T.F.; Dîrlea, S.A.; Beteg, F.I. Applications of bacterial-synthesized cellulose in veterinary medicine—A review. Acta Vet. Brno 2019, 88, 451–471. [Google Scholar] [CrossRef] [Green Version]
- Andriani, D.; Apriyana, A.Y.; Karina, M. The optimization of bacterial cellulose production and its applications: A review. Cellulose 2020, 27, 6747–6766. [Google Scholar] [CrossRef]
- Moniri, M.; Moghaddam, A.B.; Azizi, S.; Rahim, R.A.; Bin Ariff, A.; Saad, W.Z.; Navaderi, M.; Mohamad, R. Production and status of bacterial cellulose in biomedical engineering. Nanomaterials 2017, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Mohammadkazemi, F.; Azin, M.; Ashori, A. Production of bacterial cellulose using different carbon sources and culture media. Carbohydr. Polym. 2015, 117, 518–523. [Google Scholar] [CrossRef]
- Zahan, K.A.; Pa’e, N.; Muhamad, I.I. Monitoring the effect of pH on bacterial cellulose production and Acetobacter xylinum 0416 growth in a rotary discs reactor. Arab. J. Sci. Eng. 2015, 40, 1881–1885. [Google Scholar] [CrossRef]
- Sani, A.; Dahman, Y. Improvements in the production of bacterial synthesized biocellulose nanofibres using different culture methods. J. Chem. Technol. Biotechnol. 2009, 85, 151–165. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Żywicka, A.; Drozd, R.; Kordas, M.; Rakoczy, R. Effect of Gluconacetobacter xylinus cultivation conditions on the selected properties of bacterial cellulose. Pol. J. Chem. Technol. 2016, 18, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, D.; Flanagan, B.M.; Dykes, G.A.; Gidley, M.J. Influence of different carbon sources on bacterial cellulose production by Gluconacetobater xylinus strain ATCC 53524. J. Appl. Microbiol. 2008, 107, 576–583. [Google Scholar] [CrossRef]
- Rajaeifar, M.A.; Hemayati, S.S.; Tabatabaei, M.; Aghbashlo, M.; Mahmoudi, S.B. A review on beet sugar industry with a focus on implementation of waste-to-energy strategy for power supply. Renew. Sustain. Energy Rev. 2019, 103, 423–442. [Google Scholar] [CrossRef]
- Meghana, M.; Shastri, Y. Sustainable valorization of sugar industry waste: Status, opportunities, and challenges. Bioresour. Technol. 2020, 303, 122929. [Google Scholar] [CrossRef]
- Sagar, N.A.; Pareek, S.; Sharma, S.; Yahia, E.M.; Lobo, M.G. Fruit and vegetable waste: Bioactive compounds, their extraction, and possible utilization. Compr. Rev. Food Sci. Food Saf. 2018, 17, 512–531. [Google Scholar] [CrossRef] [Green Version]
- Esparza, I.; Jiménez-Moreno, N.; Bimbela, F.; Ancín-Azpilicueta, C.; Gandía, L.M. Fruit and vegetable waste management: Conventional and emerging approaches. J. Environ. Manag. 2020, 265, 110510. [Google Scholar] [CrossRef]
- Sinha, S.; Srivastava, A.; Mehrotra, T.; Singh, R. A review on the dairy industry waste: Water characterisctic, its impact on environment and treatment possibilieties. Issues Environ. Sci. Technol. 2019, 73–84. [Google Scholar]
- Kulshrestha, S.; Tyagi, P.; Sindhi, V.; Yadavilli, K.S. Invertase and its applications—A brief review. J. Pharm. Res. 2013, 7, 792–797. [Google Scholar] [CrossRef]
- Albanese, J.A.F.; Ruiz, M.P. Solid Waste as a Renewable Resource; Apple Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Ruan, Y.-L.; Jin, Y.; Yang, Y.-J.; Li, G.-J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef]
- Feijoo, S.; González-García, S.; Lema, J.M.; Moreira, M.T. Life cycle assessment of β-galactosidase enzyme production. J. Clean. Prod. 2017, 165, 204–212. [Google Scholar] [CrossRef]
- Payasi, A.; Mishra, N.N.; Chaves, A.L.S.; Singh, R. Biochemistry of fruit softening: An overview. Physiol. Mol. Biol. Plants 2009, 15, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Lam, W.C.; Pleissner, D.; Lin, C.S.K. Production of fungal glucoamylase for glucose production from food waste. In Solid Waste as a Renewable Resource; Albanese, J.A.F., Ruiz, M.P., Eds.; Apple Academic Press: Cambridge, MA, USA, 2015; pp. 81–96. [Google Scholar]
- Shigechi, H.; Koh, J.; Fujita, Y.; Matsumoto, T.; Bito, Y.; Ueda, M.; Satoh, E.; Fukuda, H.; Kondo, A. Direct production of ethanol from raw corn starch via fermentation by use of a novel surface-engineered yeast strain codisplaying glucoamylase and alpha-amylase. Appl. Environ. Microbiol. 2004, 70, 5037–5040. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.; Wen, L.; Wang, Y.; Li, L. Research progress of glucoamylase with industrial potential. J. Food Biochem. 2022, 46, e14099. [Google Scholar] [CrossRef] [PubMed]
- Drozd, R.; Rakoczy, R.; Konopacki, M.; Frąckowiak, A.; Fijałkowski, K. Evaluation of usefulness of 2DCorr technique in assessing physicochemical properties of bacterial cellulose. Carbohydr. Polym. 2017, 161, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Pelton, R. Carboxymethyl cellulose: Polyvinylamine complex hydrogel swelling. Macromolecules 2007, 40, 1624–1630. [Google Scholar] [CrossRef]
- Kwak, M.H.; Kim, J.E.; Go, J.; Koh, E.K.; Song, S.H.; Son, H.J.; Kim, H.S.; Yun, Y.H.; Jung, Y.J.; Hwang, D.Y. Bacterial cellulose membrane produced by Acetobacter sp. A10 for burn wound dressing applications. Carbohydr. Polym. 2015, 122, 387–398. [Google Scholar] [CrossRef]
- Park, S.; Baker, J.O.; Himmel, M.E.; Parilla, P.A.; Johnson, D.K. Cellulose crystallinity index: Measurement techniques and their impact on interpreting cellulase performance. Biotechnol. Biofuels 2010, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-S.; Han, Y.-H.; Chen, J.-L.; Zhang, D.-C.; Shi, X.-X.; Ye, Y.-X.; Chen, D.-L.; Li, M. Insights into bacterial cellulose biosynthesis from different carbon sources and the associated biochemical transformation pathways in Komagataeibacter sp. W1. Polymers 2018, 10, 963. [Google Scholar] [CrossRef] [Green Version]
- Molina-Ramírez, C.; Castro, M.; Osorio, M.; Torres-Taborda, M.; Gómez, B.; Zuluaga, R.; Gómez, C.; Gañán, P.; Rojas, O.J.; Castro, C. Effect of different carbon sources on bacterial nanocellulose production and structure using the low pH resistant strain Komagataeibacter medellinensis. Materials 2017, 10, 639. [Google Scholar] [CrossRef] [Green Version]
- Keshk, S.M.A.S.; Sameshima, K. Evaluation of different carbon sources for bacterial cellulose production. Afr. J. Biotechnol. 2005, 4, 478–482. [Google Scholar]
- Ruka, D.R.; Simon, G.P.; Dean, K.M. Altering the growth conditions of Gluconacetobacter xylinus to maximize the yield of bacterial cellulose. Carbohydr. Polym. 2012, 89, 613–622. [Google Scholar] [CrossRef]
- Klein, J.; Rosenberg, M.; Markoš, J.; Dolgoš, O.; Krošlák, M.; Krištofíková, L. Biotransformation of glucose to gluconic acid by Aspergillus niger—Study of mass transfer in an airlift bioreactor. Biochem. Eng. J. 2002, 10, 197–205. [Google Scholar] [CrossRef]
- El-Saied, H.; Basta, A.H.; Gobran, R.H. Research progress in friendly environmental technology for the production of cellulose products (bacterial cellulose and its application). Polym. Plast. Technol. Eng. 2004, 43, 797–820. [Google Scholar] [CrossRef]
- Talley, K.; Alexov, E. On the pH-optimum of activity and stability of proteins. Proteins 2010, 78, 2699–2706. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Yuan, L.; Wang, H.; Chen, H.; Brash, J.L. Improving the protein activity and stability under acidic conditions via site-specific conjugation of a pH-responsive polyelectrolyte. J. Mater. Chem. B 2015, 3, 498–504. [Google Scholar] [CrossRef]
- Kornmann, H.; Duboc, P.; Marison, I.; Stockar, U. von. Influence of nutritional factors on the nature, yield, and composition of exopolysaccharides produced by Gluconacetobacter xylinus I-2281. Appl. Environ. Microbiol. 2003, 69, 6091–6098. [Google Scholar] [CrossRef] [Green Version]
- Nisha, S.; Arun Karthick, S.; Gobi, N. A review on methods, application and properties of immobilizsed enzyme. Chem. Sci. Rev. Lett. 2012, 1, 148–155. [Google Scholar]
- Brás, N.F.; Fernandes, P.A.; Ramos, M.J. QM/MM Studies on the β-galactosidase catalytic mechanism: Hydrolysis and transglycosylation reactions. J. Chem. Theory Comput. 2010, 6, 421–433. [Google Scholar] [CrossRef]
- Ul-Islam, M.; Ha, J.H.; Khan, T.; Park, J.K. Effects of glucuronic acid oligomers on the production, structure and properties of bacterial cellulose. Carbohydr. Polym. 2013, 92, 360–366. [Google Scholar] [CrossRef]
- Choi, S.M.; Shin, E.J. The Nanofication and functionalization of bacterial cellulose and its applications. Nanomaterials 2020, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, G.; de Mello, C.V.; Chiari-Andréo, B.G.; Isaac, V.L.B.; Ribeiro, S.J.L.; Pecoraro, É.; Trovatti, E. Bacterial cellulose skin masks-Properties and sensory tests. J. Cosmet. Dermatol. 2018, 17, 840–847. [Google Scholar] [CrossRef]
- Ullah, M.W.; Ul-Islam, M.; Khan, S.; Kim, Y.; Park, J.K. Structural and physico-mechanical characterization of bio-cellulose produced by a cell-free system. Carbohydr. Polym. 2016, 136, 908–916. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Archer, A.J.; Chen, X.; Liu, C.; Yang, G.; Liu, Y. Dehydration of bacterial cellulose and the water content effects on its viscoelastic and electrochemical properties. Sci. Technol. Adv. Mater. 2018, 19, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Ul-Islam, M.; Khan, T.; Park, J.K. Water holding and release properties of bacterial cellulose obtained by in situ and ex situ modification. Carbohydr. Polym. 2012, 88, 596–603. [Google Scholar] [CrossRef]
- Treesuppharat, W.; Rojanapanthu, P.; Siangsanoh, C.; Manuspiya, H.; Ummartyotin, S. Synthesis and characterization of bacterial cellulose and gelatin-based hydrogel composites for drug-delivery systems. Biotechnol. Rep. 2017, 15, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Yang, G.; Hong, F. Preparation and evaluation of a kind of bacterial cellulose dry films with antibacterial properties. Carbohydr. Polym. 2011, 84, 533–538. [Google Scholar] [CrossRef]
- Wan, Y.Z.; Luo, H.; He, F.; Liang, H.; Huang, Y.; Li, X.L. Mechanical, moisture absorption, and biodegradation behaviours of bacterial cellulose fibre-reinforced starch biocomposites. Compos. Sci. Technol. 2009, 69, 1212–1217. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Initial Amount of Enzyme in Medium (U/mL) | Remaining Activity * |
|---|---|---|
| (%) | ||
| β-D-Fru | 1.6 | 34.04 ± 14.35 ˟ |
| 3.2 | 29.50 ± 14.04 | |
| 4.8 | 38.05 ± 16.66 | |
| 6.2 | 48.20 ± 19.91 | |
| β-Gal | 3.0 | 12.59 ± 9.43 |
| 6.0 | 7.37 ± 5.19 | |
| 12 | 3.93 ± 2.07 | |
| 60 | 1.66 ± 1.90 | |
| GLA | 5.0 | 14.15 ± 2.77 |
| 10 | 23.60 ± 6.86 | |
| 20 | 32.69 ± 3.12 | |
| 40 | 50.96 ± 6.44 |
| Sample | Initial Amount of Enzyme in Medium (U/mL) | XRD | ATR-FTIR | ||
|---|---|---|---|---|---|
| CrI (%) | Crystallites Size (nm) | LOI | TCI | ||
| Control | — | 93.6 | 5.9 | 1.75 ± * 0.095 | 1.46 ± 0.10 |
| β-D-Fru | 1.6 | 95.1 | 5.9 | 2.04 ± 0.64 | 1.49 ± 0.31 |
| 3.2 | 92.4 | 5.4 | 1.80 ± 0.38 | 1.46 ± 0.18 | |
| 4.8 | 90.1 | 5.6 | 1.89 ± 0.51 | 1.37 ± 0.07 | |
| 6.2 | 91.8 | 5.6 | 1.79 ± 0.37 | 1.33 ± 0.15 | |
| β-Gal | 3.0 | 91.6 | 5.6 | 2.17 ± 0.42 | 1.53 ± 0.23 |
| 6.0 | 91.6 | 5.6 | 2.17 ± 0.48 | 1.74 ± 0.14 | |
| 12 | 94.1 | 5.5 | 2.12 ± 0.40 | 1.63 ± 0.14 | |
| 60 | 94.2 | 5.8 | 2.06± 0.34 | 2.23 ± 0.19 | |
| GLA | 5.0 | 93.9 | 5.9 | 1.59 ± 0.26 | 1.23 ± 0.12 |
| 10 | 90.9 | 5.7 | 1.69 ± 0.23 | 1.35 ± 0.12 | |
| 20 | 91.8 | 5.6 | 1.80 ± 0.19 | 1.39 ± 0.08 | |
| 40 | 91.2 | 5.5 | 1.62 ± 0.35 | 1.44 ± 0.12 | |
| Enzyme | Initial Amount in the Medium (U/mL) | Moisture Content Ratio (%) |
|---|---|---|
| Control | — | 98.99 ± * 0.08 |
| β-D-Fru | 1.6 | 98.92 ± 0.13 |
| 3.2 | 99.00 ± 0.10 | |
| 4.8 | 99.01 ± 0.06 | |
| 6.2 | 98.97 ± 0.06 | |
| β-Gal | 3.0 | 99.01 ± 0.17 |
| 6.0 | 99.07 ± 0.13 | |
| 12 | 98.96 ± 0.12 | |
| 60 | 98.62 ± 0.45 | |
| GLA | 5.0 | 98.83 ± 0.13 |
| 10 | 98.96 ± 0.16 | |
| 20 | 99.02 ± 0.18 | |
| 40 | 99.05 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przygrodzka, K.; Charęza, M.; Banaszek, A.; Zielińska, B.; Ekiert, E.; Drozd, R. Bacterial Cellulose Production by Komagateibacter xylinus with the Use of Enzyme-Degraded Oligo- and Polysaccharides as the Substrates. Appl. Sci. 2022, 12, 12673. https://doi.org/10.3390/app122412673
Przygrodzka K, Charęza M, Banaszek A, Zielińska B, Ekiert E, Drozd R. Bacterial Cellulose Production by Komagateibacter xylinus with the Use of Enzyme-Degraded Oligo- and Polysaccharides as the Substrates. Applied Sciences. 2022; 12(24):12673. https://doi.org/10.3390/app122412673
Chicago/Turabian StylePrzygrodzka, Katarzyna, Magdalena Charęza, Agnieszka Banaszek, Beata Zielińska, Ewa Ekiert, and Radosław Drozd. 2022. "Bacterial Cellulose Production by Komagateibacter xylinus with the Use of Enzyme-Degraded Oligo- and Polysaccharides as the Substrates" Applied Sciences 12, no. 24: 12673. https://doi.org/10.3390/app122412673
APA StylePrzygrodzka, K., Charęza, M., Banaszek, A., Zielińska, B., Ekiert, E., & Drozd, R. (2022). Bacterial Cellulose Production by Komagateibacter xylinus with the Use of Enzyme-Degraded Oligo- and Polysaccharides as the Substrates. Applied Sciences, 12(24), 12673. https://doi.org/10.3390/app122412673