
Orientin Alleviates Liver Inflammation via Downregulation of ZEB-2/PTEN Markers—Hepatic Stellate Cells Approach

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Isolation and Culture of Primary Hepatic Stellate Cells
2.3. Estimation of Cytokines for Liver Inflammation
2.4. Nitrate Detection Assay
2.5. RNA Extraction and RT-PCR Analysis
2.6. Western Blot
2.7. Molecular Docking
2.7.1. Protein Preparation
2.7.2. Ligand Preparation
2.8. Statistical Analysis
3. Results
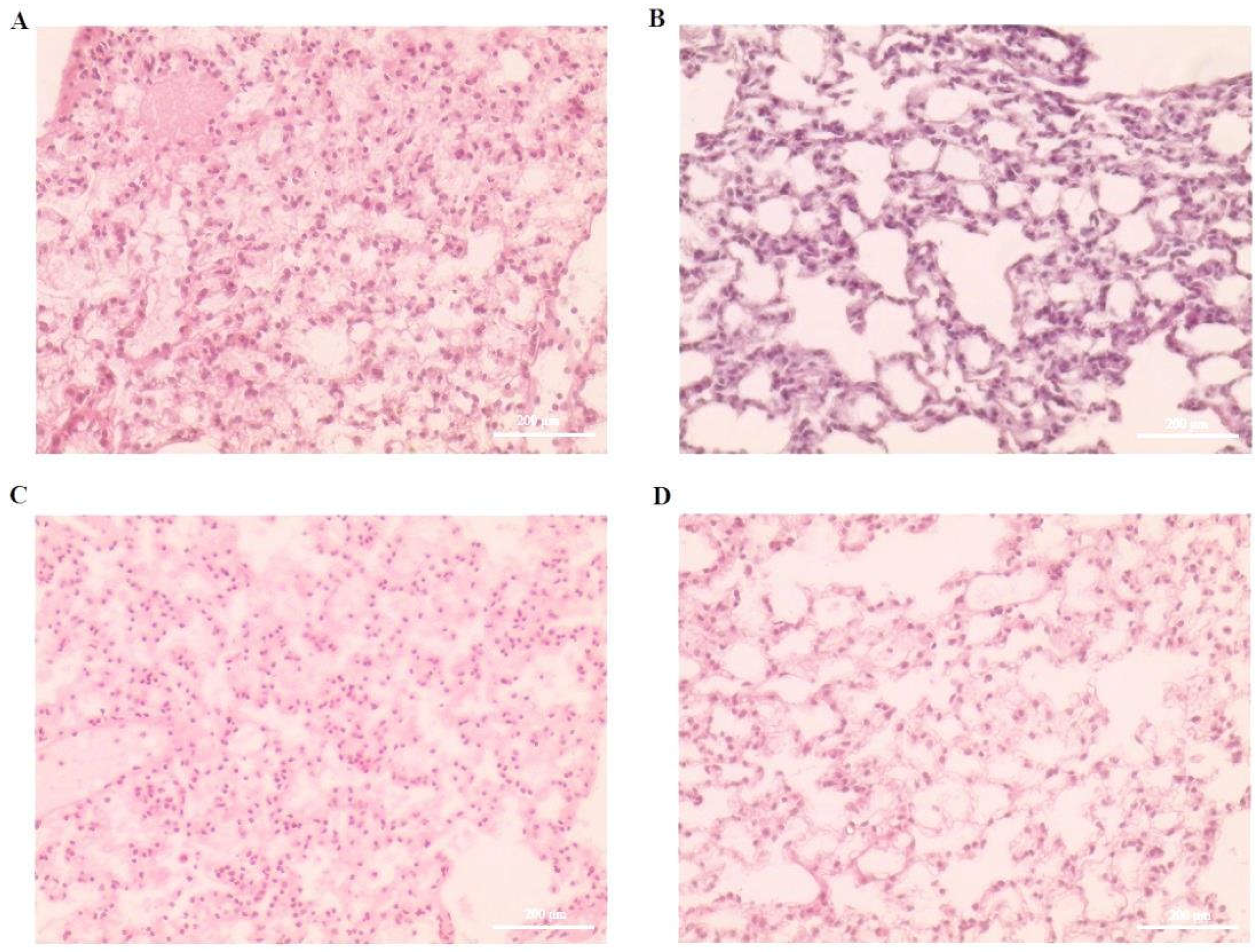
3.1. Effect of Orientin on CCl4-Induced Liver Inflammation and Immune Dysfunction
3.2. Effect of Orientin on Expression of Inflammatory Markers in CCl4-Induced Liver Tissues
3.3. Docking of Orientin with Mouse PTEN and ZEB-2
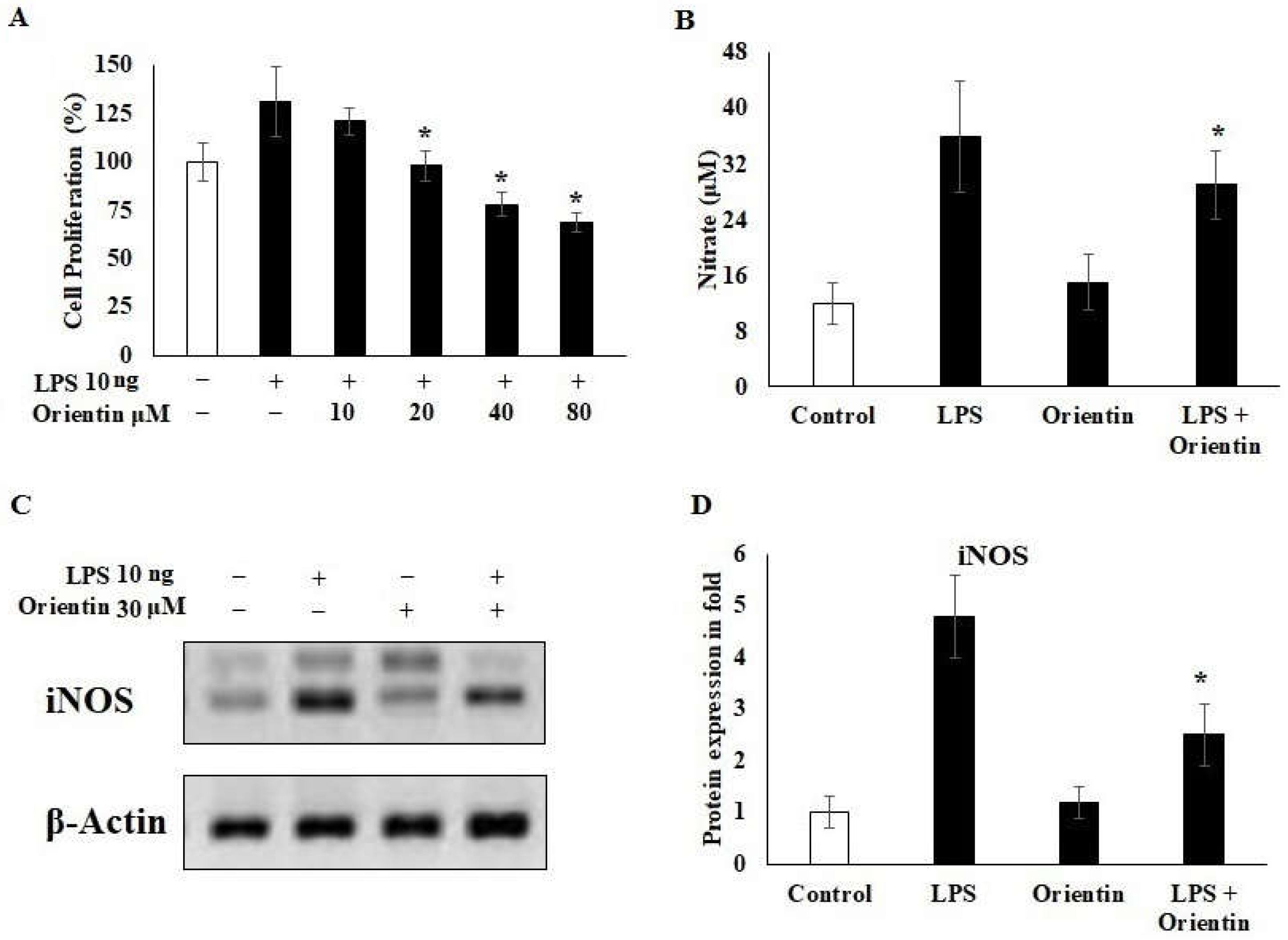
3.4. Effect of Orientin on LPS-Induced Inflammation in Hepatic Stellate Cells
3.5. Effect of Orientin on Hepatic Fibrotic Signaling Markers ZEB-2, PTEN, and TNF-α
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del Campo, J.A.; Gallego, P.; Grande, L. Role of inflammatory response in liver diseases: Therapeutic strategies. World J. Hepatol. 2018, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Calvente, C.J.; Tameda, M.; Johnson, C.D.; Del Pilar, H.; Lin, Y.C.; Adronikou, N.; Du Jeu, X.D.M.; Llorente, C.; Boyer, J.; Feldstein, A.E. Neutrophils contribute to spontaneous resolution of liver inflammation and fibrosis via microRNA-223. J. Clin. Investig. 2019, 129, 4091–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [PubMed]
- Nallagangula, K.S.; Nagaraj, S.K.; Venkataswamy, L.; Chandrappa, M. Liver fibrosis: A compilation on the biomarkers status and their significance during disease progression. Future Sci. OA 2017, 4, FSO250. [Google Scholar]
- De Haan, W.; Dheedene, W.; Apelt, K.; Décombas-Deschamps, S.; Vinckier, S.; Verhulst, S.; Conidi, A.; Deffieux, T.; Staring, M.W.; Vandervoort, P. Endothelial Zeb2 preserves the hepatic angioarchitecture and protects against liver fibrosis. Cardiovasc. Res. 2021, 1–14. [Google Scholar] [CrossRef]
- Jin, H.; Li, C.; Dong, P.; Huang, J.; Yu, J.; Zheng, J. Circular RNA cMTO1 promotes PTEN expression through sponging miR-181b-5p in liver fibrosis. Front. Cell. Dev. Biol. 2020, 8, 714. [Google Scholar] [CrossRef]
- Wandrer, F.; Liebig, S.; Marhenke, S.; Vogel, A.; John, K.; Manns, M.P.; Teufel, A.; Itzel, T.; Longerich, T.; Maier, O. TNF-Receptor-1 inhibition reduces liver steatosis, hepatocellular injury and fibrosis in NAFLD mice. Cell Death Dis. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Prystupa, A.; Kiciński, P.; Sak, J.; Boguszewska-Czubara, A.; Toruń-Jurkowska, A.; Załuska, W. Proinflammatory cytokines (IL-1α, IL-6) and hepatocyte growth factor in patients with alcoholic liver cirrhosis. Gastroenterol. Res. Pract. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Attallah, A.M.; El-Far, M.; Zahran, F.; Shiha, G.E.; Farid, K.; Omran, M.M.; Abdelrazek, M.A.; Attallah, A.A.; El-Beh, A.A.; El-Hosiny, R.M. Interferon-gamma is associated with hepatic dysfunction in fibrosis, cirrhosis, and hepatocellular carcinoma. J. Immunoass. Immunochem. 2016, 37, 597–610. [Google Scholar] [CrossRef]
- Gonzalez-Gallego, J.; Sánchez-Campos, S.; Tunon, M. Anti-inflammatory properties of dietary flavonoids. Nutr. Hosp. 2007, 22, 287–293. [Google Scholar] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Ciumărnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, Ș.C.; Răchișan, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.-G.; Para, I. The effects of flavonoids in cardiovascular diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef] [PubMed]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential role of flavonoids in treating chronic inflammatory diseases with a special focus on the anti-inflammatory activity of apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Aquino, E.; Muriel, P. Beneficial effects of naringenin in liver diseases: Molecular mechanisms. World J. Gastroenterol. 2018, 24, 1679. [Google Scholar] [CrossRef]
- Raso, G.M.; Meli, R.; Di Carlo, G.; Pacilio, M.; Di Carlo, R. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by flavonoids in macrophage J774A. 1. Life Sci. 2001, 68, 921–931. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Ku, S.-K.; Bae, J.-S. Vascular barrier protective effects of orientin and isoorientin in LPS-induced inflammation in vitro and in vivo. Vascul. Pharmacol. 2014, 62, 3–14. [Google Scholar] [CrossRef]
- Dantas-Medeiros, R.; Furtado, A.A.; Zanatta, A.C.; Torres-Rêgo, M.; Lourenço, E.M.G.; Alves, J.S.F.; Galinari, É.; de Oliveira Rocha, H.A.; Guerra, G.C.B.; Vilegas, W. Mass spectrometry characterization of Commiphora leptophloeos leaf extract and preclinical evaluation of toxicity and anti-inflammatory potential effect. J. Ethnopharmacol. 2021, 264, 113229. [Google Scholar] [CrossRef]
- Lam, K.Y.; Ling, A.P.K.; Koh, R.Y.; Wong, Y.P.; Say, Y.H. A review on medicinal properties of orientin. Adv. Pharmacol. Sci. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Batool, R.; Kalsoom, A.; Akbar, I.; Arshad, N.; Jamil, N. Antilisterial Effect of Rosa damascena and Nymphaea alba in Mus musculus. Biomed. Res. Int. 2018, 2018, 1–9. [Google Scholar]
- Abtulov, M.; Valcheva-Kuzmanova, S. Flavonoids and the liver—A review. Scr. Sci. Pharm. 2021, 8, 7–16. [Google Scholar]
- Wu, Y.; He, Y.; Wang, R.; Zhao, X. Preventive Effect of Flavonoid Extract from the Peel of Gonggan (Citrus reticulata Blanco Var. Gonggan) on CCl4-Induced Acute Liver Injury in Mice. J. Inflamm. Res. 2021, 14, 5111. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, O.R.; Oguntibeju, O.O.; Marnewick, J.L. Amelioration of lipopolysaccharide-induced liver injury by aqueous rooibos (Aspalathus linearis) extract via inhibition of pro-inflammatory cytokines and oxidative stress. BMC Complement. Altern. Med. 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.-H.; Huang, J.-H.; Sun, M.-S.; Tzeng, I.; Hsu, Y.-C.; Kuo, C.-Y. Wild bitter melon extract regulates LPS-induced hepatic stellate cell activation, inflammation, endoplasmic reticulum stress, and ferroptosis. Evid. Based Complement. Alternat. Med. 2021, 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rong, X.; Xu, J.; Jiang, Y.; Li, F.; Chen, Y.; Dou, Q.P.; Li, D. Citrus peel flavonoid nobiletin alleviates lipopolysaccharide-induced inflammation by activating IL-6/STAT3/FOXO3a-mediated autophagy. Food Funct. 2021, 12, 1305–1317. [Google Scholar] [CrossRef]
- Yan, G.; Chen, L.; Wang, H.; Wu, S.; Li, S.; Wang, X. Baicalin inhibits LPS-induced inflammation in RAW264. 7 cells through miR-181b/HMGB1/TRL4/NF-κB pathway. Am. J. Transl. Res. 2021, 13, 10127. [Google Scholar]
- Zamani-Garmsiri, F.; Emamgholipour, S.; Rahmani Fard, S.; Ghasempour, G.; Jahangard Ahvazi, R.; Meshkani, R. Polyphenols: Potential anti-inflammatory agents for treatment of metabolic disorders. Phytother. Res. 2021, 36, 415–432. [Google Scholar] [CrossRef]
- Mederacke, I.; Dapito, D.H.; Affò, S.; Uchinami, H.; Schwabe, R.F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nat. Protoc. 2015, 10, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Goshi, E.; Zhou, G.; He, Q. Nitric oxide detection methods in vitro and in vivo. Med. Gas Res. 2019, 9, 192. [Google Scholar] [CrossRef]
- Khalil, H.E.; Ibrahim, H.-I.M.; Ahmed, E.A.; Emeka, P.M.; Alhaider, I.A. Orientin, a Bio-Flavonoid from Trigonella hamosa L., Regulates COX-2/PGE-2 in A549 Cell Lines via miR-26b and miR-146a. Pharmaceuticals 2022, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.E.; Ibrahim, H.-I.M.; Darrag, H.M.; Matsunami, K. Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers. Plants 2022, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanasambantham, K.; Muralidaran, S.; Mandal, A.K.A. Molecular cloning, computational and expression analysis of anthocyanidin reductase in tea (Camellia sinensis). Appl. Biochem. Biotechnol. 2014, 174, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Takasu, C.; Vaziri, N.D.; Li, S.; Robles, L.; Vo, K.; Takasu, M.; Pham, C.; Farzaneh, S.H.; Shimada, M.; Stamos, M.J. Treatment with dimethyl fumarate ameliorates liver ischemia/reperfusion injury. World J. Gastroenterol. 2017, 23, 4508. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Chakraborty, A.K.; Dey, P.; Kar, P.; Guha, P.; Sen, S.; Kumar, A.; Sen, A.; Chaudhuri, T.K. Amelioration of CCl4 induced liver injury in swiss albino mice by antioxidant rich leaf extract of Croton bonplandianus Baill. PLoS ONE 2018, 13, e0196411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.N.; Kim, S.H.; Dey, D.K.; Park, S.M.; Nasif, O.; Bajpai, V.K.; Kang, S.C.; Lee, J.; Park, J.G. 5-O-Demethylnobiletin alleviates CCl4-Induced acute liver injury by equilibrating ROS-mediated apoptosis and autophagy induction. Int. J. Mol. Sci. 2021, 22, 1083. [Google Scholar] [CrossRef]
- Akindele, A.J.; Oludadepo, G.O.; Amagon, K.I.; Singh, D.; Osiagwu, D.D. Protective effect of carvedilol alone and coadministered with diltiazem and prednisolone on doxorubicin and 5-fluorouracil-induced hepatotoxicity and nephrotoxicity in rats. Pharmacol. Res. Perspect. 2018, 6, e00381. [Google Scholar] [CrossRef] [Green Version]
- Gharbia, S.; Balta, C.; Herman, H.; Rosu, M.; Váradi, J.; Bácskay, I.; Vecsernyés, M.; Gyöngyösi, S.; Fenyvesi, F.; Voicu, S.N. Enhancement of Silymarin Anti-fibrotic Effects by Complexation With Hydroxypropyl (HPBCD) and Randomly Methylated (RAMEB) β-Cyclodextrins in a Mouse Model of Liver Fibrosis. Front. Pharmacol. 2018, 9, 883. [Google Scholar] [CrossRef] [Green Version]
- Owumi, S.; Lewu, D.; Arunsi, U.; Oyelere, A. Luteolin attenuates doxorubicin-induced derangements of liver and kidney by reducing oxidative and inflammatory stress to suppress apoptosis. Hum. Exp. Toxicol. 2021, 40, 1656–1672. [Google Scholar] [CrossRef]
- Okda, T.; Abd-Alhaseeb, M.; Barka, K.; Ragab, N. Ginger potentiates the effects of silymarin on liver fibrosis induced by CCl4: The role of galectin-8. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 885–891. [Google Scholar] [PubMed]
- Ullah, H.; Khan, A.; Baig, M.W.; Ullah, N.; Ahmed, N.; Tipu, M.K.; Ali, H.; Khan, S. Poncirin attenuates CCl4-induced liver injury through inhibition of oxidative stress and inflammatory cytokines in mice. BMC Complement. Med. Ther. 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yoon, J.-H.; Won, H.-J.; Ji, H.-S.; Yuk, H.J.; Park, K.H.; Park, H.-Y.; Jeong, T.-S. Isotrifoliol inhibits pro-inflammatory mediators by suppression of TLR/NF-κB and TLR/MAPK signaling in LPS-induced RAW264. 7 cells. Int. Immunopharmacol. 2017, 45, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Zhao, Y.; Ma, L.; Piao, R. Orientin reverses acetaminophen-induced acute liver failure by inhibiting oxidative stress and mitochondrial dysfunction. J. Pharmacol. Sci. 2022, in press. [CrossRef]
- Bello, O.M.; Ogbesejana, A.B.; Adetunji, C.O.; Oguntoye, S.O. Flavonoids isolated from Vitex grandifolia, an underutilized vegetable, exert monoamine A & B inhibitory and anti-inflammatory effects and their structure-activity relationship. Turk. J. Pharm. Sci. 2019, 16, 437. [Google Scholar]
- Mohamed, T.K.; Kamal, A.M.; Nassar, M.I.; Ahmed, M.A.; Haggag, M.G.; Ezzat, H.A. Phenolic contents of Gleditsia triacanthos leaves and evaluation of its analgesic, anti-inflammatory, hepatoprotective and antimicrobial activities. Life Sci. 2013, 10, 3445–3446. [Google Scholar]
- Jayaraman, J.; Jesudoss, V.A.S.; Menon, V.P.; Namasivayam, N. Anti-inflammatory role of naringenin in rats with ethanol induced liver injury. Toxicol. Mech. Methods 2012, 22, 568–576. [Google Scholar] [CrossRef]
- Qian, Y.; Han, J.; Zhou, L.; Yu, Q.; Xv, J.; Jin, Z.; Yang, Y.; Jiang, L.; Lou, D. Inhibition of epidermal growth factor receptor (EGFR) reduces lipopolysaccharide (LPS)-induced activation and inflammatory cytokines in hepatic stellate cells in vitro. Med. Sci. Monit. 2018, 24, 5533. [Google Scholar] [CrossRef]
- Yin, L.; Qi, Y.; Xu, Y.; Xu, L.; Han, X.; Tao, X.; Song, S.; Peng, J. Dioscin inhibits HSC-T6 cell migration via adjusting SDC-4 expression: Insights from iTRAQ-based quantitative proteomics. Front. Pharmacol. 2017, 8, 665. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Xu, Y.; Han, X.; Yin, L.; Xu, L.; Qi, Y.; Zhao, Y.; Liu, K.; Peng, J. Dioscin alleviates alcoholic liver fibrosis by attenuating hepatic stellate cell activation via the TLR4/MyD88/NF-κB signaling pathway. Sci. Rep. 2015, 5, 1–13. [Google Scholar]
- Dey, P.; Panga, V.; Raghunathan, S. A cytokine signalling network for the regulation of inducible nitric oxide synthase expression in rheumatoid arthritis. PLoS ONE 2016, 11, e0161306. [Google Scholar] [CrossRef]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular mechanisms of liver fibrosis. Front. Pharmacol. 2021, 12, 1072. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Bian, E.-B.; Huang, C.; Ma, T.-T.; Tao, H.; Zhang, H.; Cheng, C.; Lv, X.-W.; Li, J. DNMT1-mediated PTEN hypermethylation confers hepatic stellate cell activation and liver fibrogenesis in rats. Toxicol. Appl. Pharmacol. 2012, 264, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, J.; Robuffo, I.; Spalletta, S.; Giambuzzi, G.; De Iuliis, V.; Toniato, E.; Martinotti, S.; Conti, P.; Flati, V. The epithelial-to-mesenchymal transition as a possible therapeutic target in fibrotic disorders. Front. Cell Dev. Biol. 2020, 8, 1–32. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Energy | Ligand Efficiency | Intermole Energy | Ligand Atoms (Ring) | Docked Amino Acid Residue (Bond Length) |
|---|---|---|---|---|
| −6.17 | −0.19 | −6.38 | Conventional Hydrogen Bond: | |
| Chain B—C3′-O | Chain A: ARG′36 ′HN′ (2.80 Å) | |||
| Chain B—C3′-OH | Chain A: ALA′32 ′O′ (1.80 Å) | |||
| Chain B—C4′-OH | Chain A: ALA′32 ′O′ (1.80 Å) | |||
| Chain C—C4-O | Chain A: THR′49 ′HG1′ (1.88 Å) | |||
| Chain A—C5-OH | Chain A: THR′49 ′OG1′ (1.93 Å) | |||
| C2-OH | Chain A: THR′19 ′O′ (2.20 Å) | |||
| C3-OH | Chain A: THR′19 ′OG1′ (2.18 Å) | |||
| C3-OH | Chain A: THR′19 ′O′ (2.61 Å) | |||
| Pi-Alkyl Hydrophobic bond: | ||||
| O | Chain A: ALA′32′ (4.36 Å) | |||
| O | Chain A: LEU′20′ (5.00 Å) | |||
| O | Chain A: LEU′20′ (5.41 Å) | |||
| O | Chain A: PRO′51′ (5.18 Å) | |||
| Pi-sigma bond: | ||||
| O | Chain A: LEU′20′ CD2′ (3.44 Å) | |||
| O | Chain A: ARG′166′ (4.77 Å) | |||
| Binding Energy | Ligand Efficiency | Intermole Energy | Ligand Atoms (Ring) | Docked Amino Acid Residue (Bond Length) |
|---|---|---|---|---|
| −6.44 | −0.2 | −6.44 | -Conventional Hydrogen Bond: | |
| Chain B—C4′-O | Chain A: ASN′323 ′HD22′ (1.92 Å) | |||
| Chain B—C4′-OH | Chain A: THR′161 ′O′ (1.83 Å) | |||
| Chain B—C3′-OH | Chain A: THR′161 ′O′ (2.24 Å) | |||
| Chain C—C4′-O | Chain A: GLN′143 ′HE22′ (1.90 Å) | |||
| Chain C—C4′-O | Chain A: GLN′143 ′HE21′ (2.44 Å) | |||
| C4-OH | Chain A: ARG′167 ′HH12′ (2.35 Å) | |||
| -Carbon Hydrogen Bond | ||||
| C | Chain A: ASP′318′OD2′ (3.19 Å) | |||
| C | Chain A: ARG′167′CD′ (2.90 Å) | |||
| -Pi-Alkyl Hydrophobic bond: | ||||
| O | Chain A: TRY′170′ (5.65 Å) | |||
| O | Chain A: TRY′170′ (4.81 Å) | |||
| -Pi-Pi T-shaped bond: | ||||
| O | Chain A: PRO′163′ (4.56 Å) | |||
| O | Chain A: ARG′166′ (4.77 Å) | |||
| -Pi-cation bond: | ||||
| O | Chain A: ARG′166′ NH2′ (4.35 Å) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, H.E.; Ibrahim, H.-I.M.; El-Fass, K.A.; Akrawi, S.H.; Morsy, M.A. Orientin Alleviates Liver Inflammation via Downregulation of ZEB-2/PTEN Markers—Hepatic Stellate Cells Approach. Appl. Sci. 2022, 12, 2725. https://doi.org/10.3390/app12052725
Khalil HE, Ibrahim H-IM, El-Fass KA, Akrawi SH, Morsy MA. Orientin Alleviates Liver Inflammation via Downregulation of ZEB-2/PTEN Markers—Hepatic Stellate Cells Approach. Applied Sciences. 2022; 12(5):2725. https://doi.org/10.3390/app12052725
Chicago/Turabian StyleKhalil, Hany Ezzat, Hairul-Islam Mohamed Ibrahim, Kareem Ahmed El-Fass, Sabah H. Akrawi, and Mohamed A. Morsy. 2022. "Orientin Alleviates Liver Inflammation via Downregulation of ZEB-2/PTEN Markers—Hepatic Stellate Cells Approach" Applied Sciences 12, no. 5: 2725. https://doi.org/10.3390/app12052725
APA StyleKhalil, H. E., Ibrahim, H. -I. M., El-Fass, K. A., Akrawi, S. H., & Morsy, M. A. (2022). Orientin Alleviates Liver Inflammation via Downregulation of ZEB-2/PTEN Markers—Hepatic Stellate Cells Approach. Applied Sciences, 12(5), 2725. https://doi.org/10.3390/app12052725