
Isolation and Characterization of Biohydrogen-Producing Bacteria for Biohydrogen Fermentation Using Oil Palm Biomass-Based Carbon Source

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection and Raw Material Preparation
2.1.1. Soil Sample
2.1.2. Oil Palm Empty Fruit Bunches Preconditioning
2.2. Biohydrogen-Producing Bacterium Isolation
2.2.1. Isolation Medium
2.2.2. Isolation and Purification
2.3. Bacterial Characterization
2.3.1. Gram Staining
2.3.2. BIOLOG
2.3.3. 16S rRNA Sequencing
- (a)
- Deoxyribonucleic Acid (DNA) Extraction
- (b)
- Polymerase Chain Reaction (PCR)
- (c)
- PCR Product Purification
- (d)
- Agarose Gel Electrophoresis
- (e)
- 16S rRNA Nucleotide Analysis
2.3.4. Field Emission Scanning Electron Microscopy (FESEM)
2.4. Biohydrogen Fermentability Test in Serum Bottles
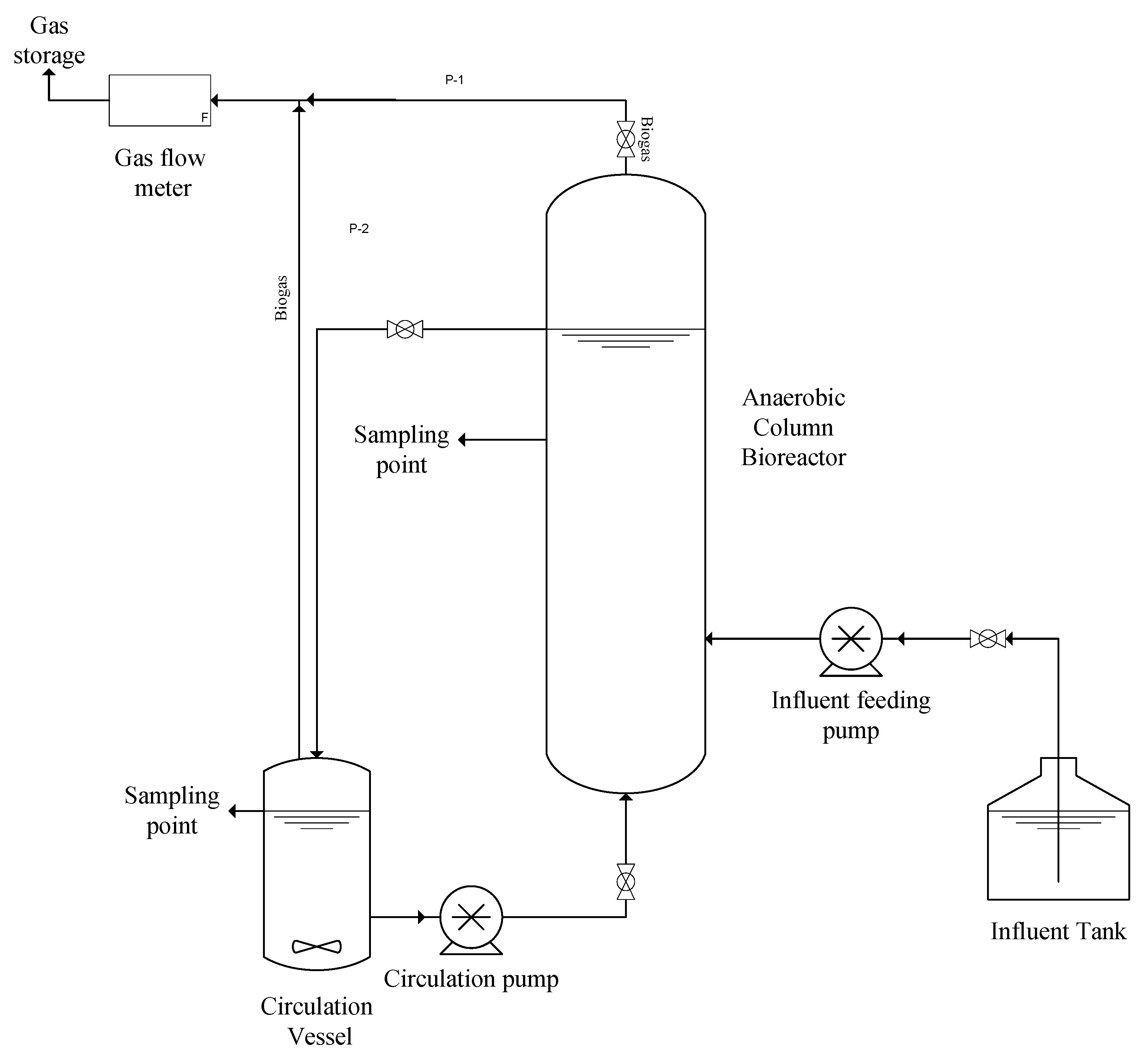
2.5. Biohydrogen Fermentation in 1.5 L Anaerobic Column Bioreactor (ACBR)
3. Results and Discussion
3.1. Bacterial Characterization
3.1.1. Gram Staining
3.1.2. BIOLOG
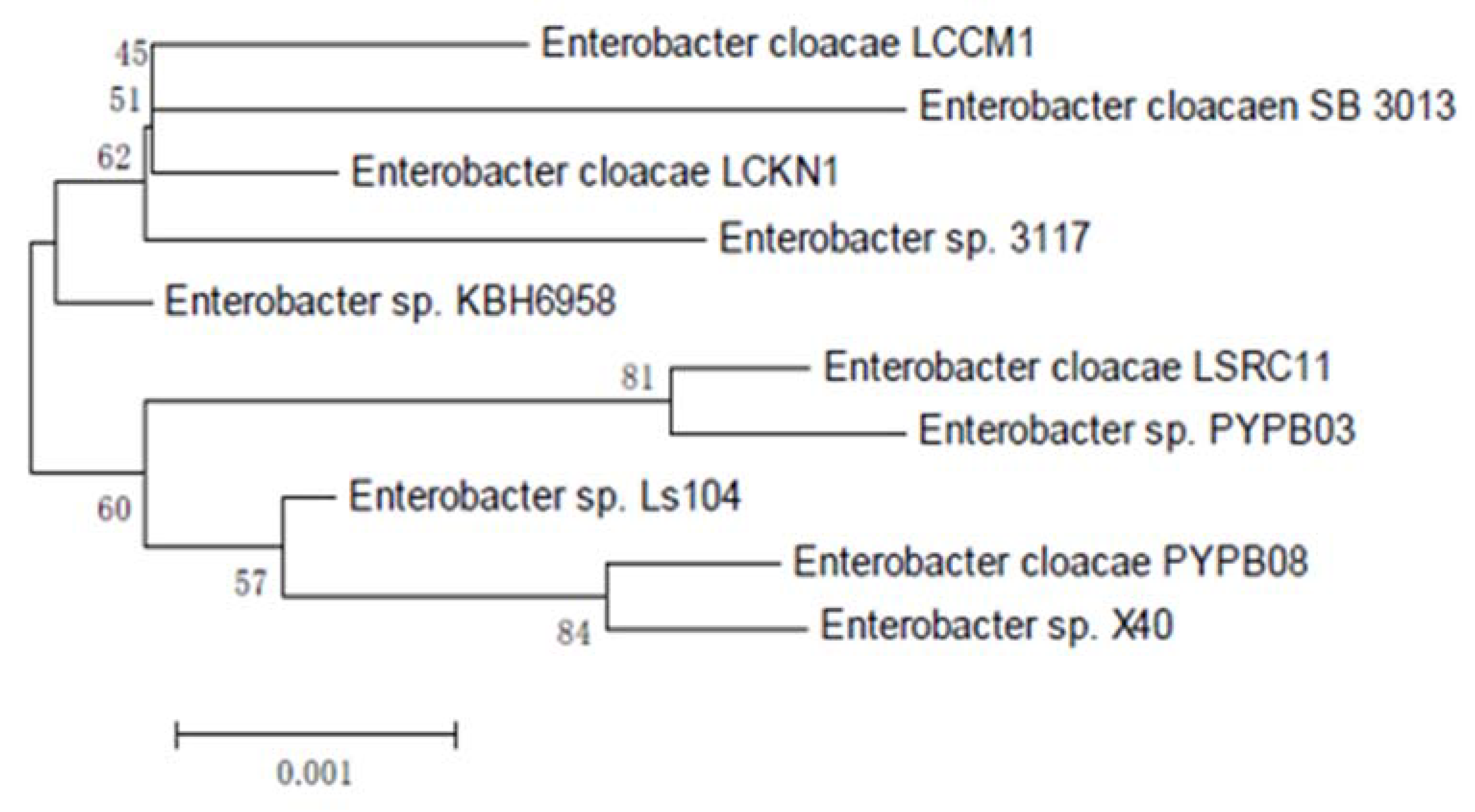
3.1.3. 16S rRNA Sequencing
3.1.4. Field Emission Scanning Electron Microscopy (FESEM)
3.2. Metabolite Analysis
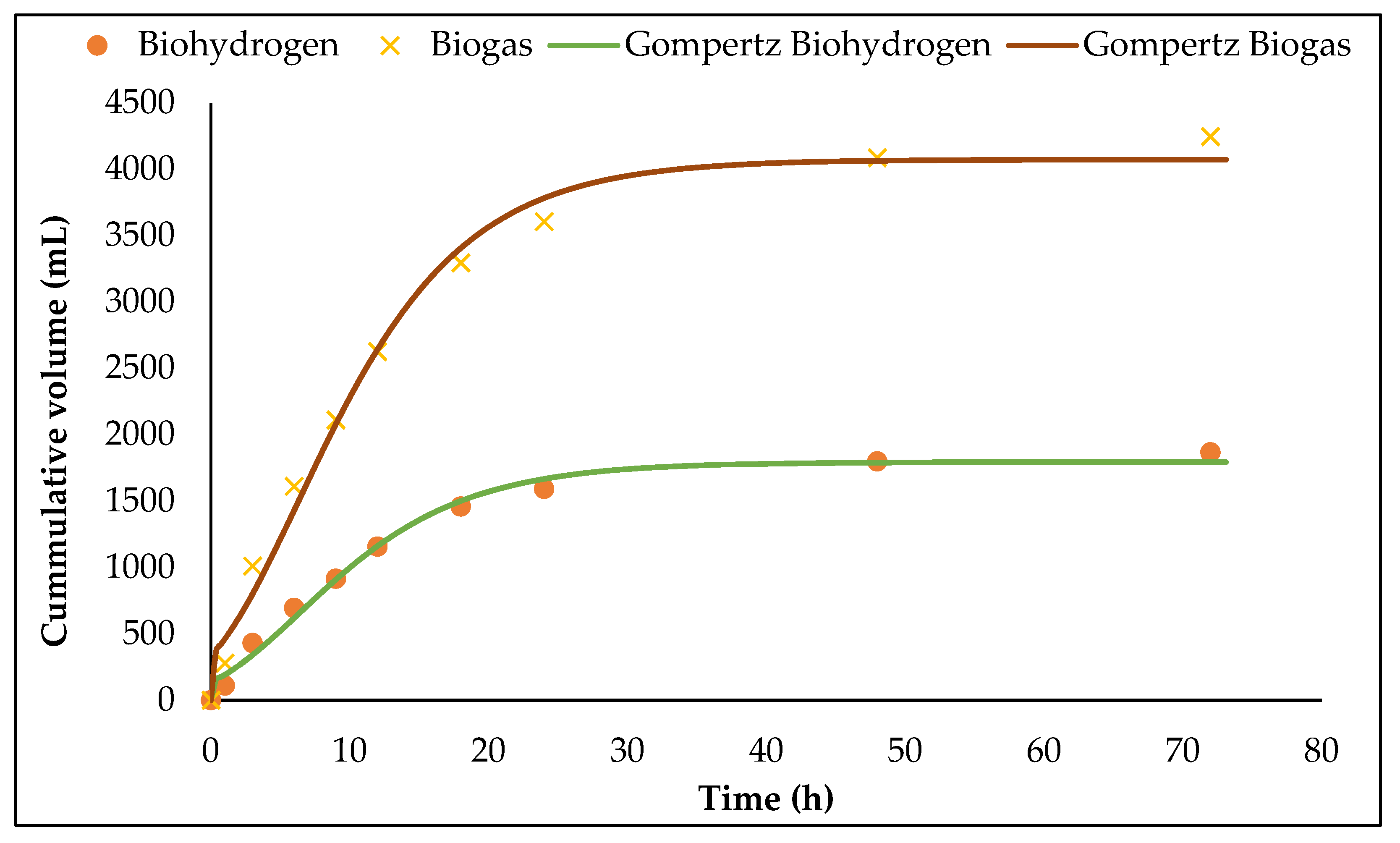
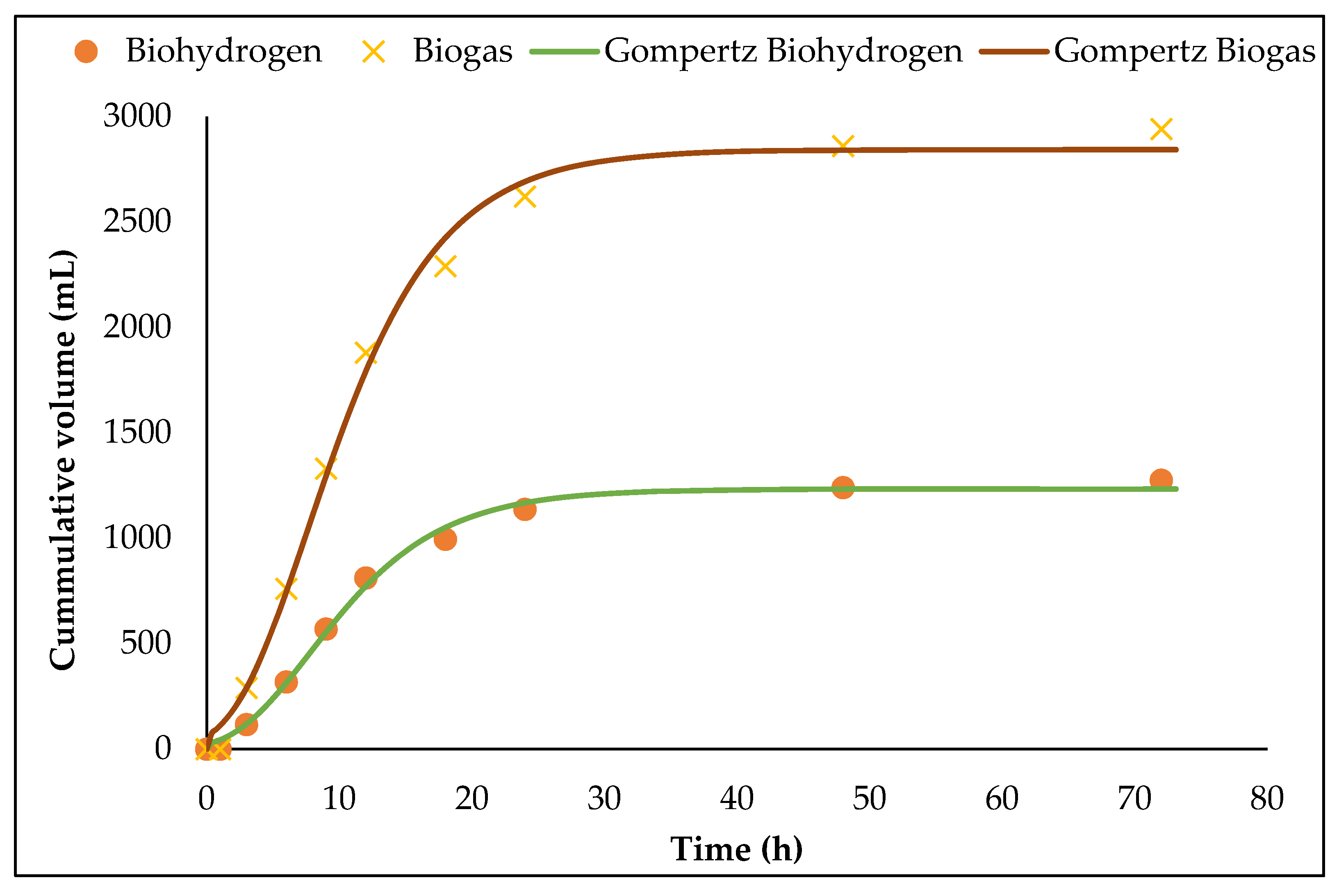
3.3. Biohydrogen Fermentation in 1.5 L ACBR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lutpi, N.A.; Jahim, J.M.; Mumtaz, T.; Abdul, P.M.; Mohd Nor, M.T. Physicochemical characteristics of attached biofilm on granular activated carbon for thermophilic biohydrogen production. RSC Adv. 2015, 5, 19382–19392. [Google Scholar] [CrossRef]
- Trchounian, K.; Sawers, R.G.; Trchounian, A. Improving biohydrogen productivity by microbial dark-and photo-fermentations: Novel data and future approaches. Renew. Sustain. Energy Rev. 2017, 80, 1201–1216. [Google Scholar] [CrossRef]
- Hawkes, F.; Dinsdale, R.; Hawkes, D.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimisation. Int. J. Hydrogen Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Kotay, S.M.; Das, D. Biohydrogen as a renewable energy resource—Prospects and potentials. Int. J. Hydrogen Energy 2008, 33, 258–263. [Google Scholar]
- Kapdan, I.K.; Kargi, F. Bio-hydrogen production from waste materials. Enz. Microb. Technol. 2006, 38, 569–582. [Google Scholar] [CrossRef]
- Ntaikou, I.; Antonopoulou, G.; Lyberatos, G. Biohydrogen production from biomass and wastes via dark fermentation: A review. Waste Biomass Valor. 2010, 1, 21–39. [Google Scholar] [CrossRef]
- Wang, J.; Yin, Y. Principle and application of different pretreatment methods for enriching hydrogen-producing bacteria from mixed cultures. Int. J. Hydrogen Energy 2017, 42, 4804–4823. [Google Scholar] [CrossRef]
- Harun, I.; Jahim, J.M.; Anuar, N.; Hassan, O. Hydrogen production performance by Enterobacter cloacae KBH3 isolated from termite guts. Int. J. Hydrogen Energy 2012, 37, 15052–15061. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Abo-Hashesh, M.; Ghosh, D. Strategies for improving biological hydrogen production. Bioresour. Technol. 2012, 110, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Pandey, A. An evaluative report and challenges for fermentative biohydrogen production. Int. J. Hydrogen Energy 2011, 36, 7460–7478. [Google Scholar] [CrossRef]
- Yasin, N.H.M.; Fukuzaki, M.; Maeda, T.; Miyazaki, T.; Hakiman Che Maail, C.M.; Ariffin, H.; Wood, T.K. Biohydrogen production from oil palm frond juice and sewage sludge by a metabolically engineered Escherichia coli strain. Int. J. Hydrogen Energy 2013, 38, 10277–10283. [Google Scholar] [CrossRef]
- Norfadilah, N.; Raheem, A.; Harun, R. Bio-hydrogen production from palm oil mill effluent (POME): A preliminary study. Int. J. Hydrogen Energy 2016, 41, 11960–11964. [Google Scholar] [CrossRef]
- Abdul, P.M.; Jahim, J.M.; Harun, S.; Markom, M.; Hassan, O.; Mohammad, A.W.; Asis, A.J. Biohydrogen production from pentose-rich oil palm empty fruit bunch molasses: A first trial. Int. J. Hydrogen Energy 2013, 38, 15693–15699. [Google Scholar] [CrossRef]
- MPOB. Fresh Fruit Bunch (FFB) Received by Mill for the Month of December 2014. Available online: http://bepi.mpob.gov.my/index.php/statistics/sectoral-status/127-sectoral-status-2014/674-ffb-received-by-mill-2014.html (accessed on 6 July 2015).
- Rahman, S.H.A.; Choudhury, J.P.; Ahmad, A.L.; Kamaruddin, A.H. Optimization studies on acid hydrolysis of oil palm empty fruit bunch fiber for production of xylose. Bioresour. Technol. 2007, 98, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Prasertsan, S.; Prasertsan, P. Biomass residues from palm oil mills in Thailand: An overview on quantity and potential usage. Biomass Bioenerg. 1996, 11, 387–395. [Google Scholar] [CrossRef]
- Lau, M.J.; Lau, M.W.; Gunawan, C.; Dale, B.E. Ammonia fiber expansion (AFEX) pretreatment, enzymatic hydrolysis, and fermentation on empty palm fruit bunch fiber (EPFBF) for cellulosic ethanol production. Appl. Biochem. Biotechnol. 2010, 162, 1847–1857. [Google Scholar] [CrossRef]
- Luthfi, A.A.I.; Jahim, J.M.; Harun, S.; Tan, J.P.; Mohammad, A.W. Biorefinery approach towards greener succinic acid production from oil palm frond bagasse. Proc. Biochem. 2016, 51, 1527–1537. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, Z.; Jiao, Y.; Li, C.; Zheng, Y.; Lin, Y.; Xiao, J.; Chen, Z.; Cao, P. Histological identification of Propionibacterium acnes in nonpyogenic degenerated intervertebral discs. BioMed Res. Int. 2017, 2017, 6192935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Molec. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Mecozzi, M. Estimation of total carbohydrate amount in environmental samples by the phenol–sulphuric acid method assisted by multivariate calibration. Chemom. Intell. Lab. Syst. 2005, 79, 84–90. [Google Scholar] [CrossRef]
- Turner, R.D.; Hurd, A.F.; Cadby, A.; Hobbs, J.K.; Foster, S.J. Cell wall elongation mode in Gram-negative bacteria is determined by peptidoglycan architecture. Nat. Commun. 2013, 4, 1469. [Google Scholar] [CrossRef] [PubMed]
- Wragg, P.; Randall, L.; Whatmore, A.M. Comparison of Biolog GEN III MicroStation semi-automated bacterial identification system with matrix-assisted laser desorption ionization-time of flight mass spectrometry and 16S ribosomal RNA gene sequencing for the identification of bacteria of veterinary interest. J. Microbiol. Methods 2014, 105, 16–21. [Google Scholar] [PubMed] [Green Version]
- Goswami, A.; Stewart, J. Organic Synthesis Using Biocatalysis; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Sundara Sekar, B.; Seol, E.; Park, S. Co-production of hydrogen and ethanol from glucose in Escherichia coli by activation of pentose-phosphate pathway through deletion of phosphoglucose isomerase (pgi) and overexpression of glucose-6-phosphate dehydrogenase (zwf) and 6-phosphogluconate dehydrogenase (gnd). Biotechnol. biofuel. 2017, 10, 85. [Google Scholar]
- Rahim, N.A.; Indera Luthfi, A.A.; Abdul, P.M.; Jahim, J.M.; Bukhari, N.A. Towards Sustainable Production of Bio-based Lactic Acid via a Bio-based Technical Route: Recent Developments and the Use of Palm Kernel Cakes in the Bioconversion. BioResources 2022, 17, 3781–3809. [Google Scholar] [CrossRef]
- de Amorim, E.L.C.; Sader, L.T.; Silva, E.L. Effect of substrate concentration on dark fermentation hydrogen production using an anaerobic fluidized bed reactor. Appl. Biochem. Biotechnol. 2012, 166, 1248–1263. [Google Scholar] [CrossRef] [PubMed]
- Bundhoo, M.Z.; Mohee, R. Inhibition of dark fermentative bio-hydrogen production: A review. Int. J. Hydrogen Energy 2016, 41, 6713–6733. [Google Scholar] [CrossRef]
- Public Health. Identification of Enterobacteriaceae. 2015, Volume 16, pp. 1–34. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/423601/ID_16i4.pdf (accessed on 23 December 2022).
- Luthfi, A.A.I.; Jahim, J.M.; Harun, S.; Tan, J.P.; Mohammad, A.W. Potential use of coconut shell activated carbon as an immobilisation carrier for high conversion of succinic acid from oil palm frond hydrolysate. RSC Adv. 2017, 7, 49480–49489. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Nakashimada, Y.; Kakizono, T.; Nishio, N. High-yield production of hydrogen by Enterobacter aerogenes mutants with decreased α-acetolactate synthase activity. J. Biosci. Bioeng. 2004, 97, 227–232. [Google Scholar] [CrossRef]
- Khanna, N.; Kotay, S.M.; Gilbert, J.J.; Das, D. Improvement of biohydrogen production by Enterobacter cloacae IIT-BT 08 under regulated pH. J. Biotechnol. 2011, 152, 9–15. [Google Scholar] [CrossRef]
- Hasibar, B.; Ergal, İ.; Moser, S.; Bochmann, G.; Simon, K.-M.R.; Fuchs, W. Increasing biohydrogen production with the use of a co-culture inside a microbial electrolysis cell. Biochem. Eng. J. 2020, 164, 107802. [Google Scholar] [CrossRef]
- Sikora, A.; Błaszczyk, M.; Jurkowski, M.; Zielenkiewicz, U. Lactic Acid Bacteria in hydrogen-Producing Consortia: On Purpose or by Coincidence? INTECH Open Science Open Minds: London, UK, 2013. [Google Scholar]
- Sivagurunathan, P.; Sen, B.; Lin, C.-Y. Overcoming propionic acid inhibition of hydrogen fermentation by temperature shift strategy. Int. J. Hydrogen Energy 2014, 39, 19232–19241. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lay, C. Effects of carbonate and phosphate concentrations on hydrogen production using anaerobic sewage sludge microflora. Int. J. Hydrogen Energy 2004, 29, 275–281. [Google Scholar] [CrossRef]
- Khamtib, S.; Reungsang, A. Biohydrogen production from xylose by Thermoanaerobacterium thermosaccharolyticum KKU19 isolated from hot spring sediment. Int. J. Hydrogen Energy 2012, 37, 12219–12228. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lay, C.-H.; Sen, B.; Chu, C.-Y.; Kumar, G.; Chen, C.-C.; Chang, J.-S. Fermentative hydrogen production from wastewaters: A review and prognosis. Int. J. Hydrogen Energy 2012, 37, 15632–15642. [Google Scholar] [CrossRef]
- Singh, R.; White, D.; Blum, P. Mutant microorganisms and methods of making and using. U.S. Patent 11,332,763, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Substrate | AN | Gen III | Type | Substrate | AN | Gen III |
|---|---|---|---|---|---|---|---|
| Amine/Amide | Alaninamide | 78 | N/A | Carbohydrate | Sucrose | 100 | 87 |
| Glucuronamide | N/A | 85 | D-Trehalose | 100 | 81 | ||
| Succinamic Acid | 0 | N/A | Turanose | 88 | 87 | ||
| Amino Acid | L-Alanine | 100 | 93 | Carboxylic acid | D-Galacturonic acid | 100 | 93 |
| L-Alanyl-L- Glutamine | 100 | N/A | D-Gluconic acid | 100 | 94 | ||
| L-Alanyl-L-histidine | 66 | N/A | Glucuronic Acid | N/A | 93 | ||
| L-Alanyl-L-threonine | 100 | N/A | D-Glucosaminic acid | 0 | N/A | ||
| L-Arginine | N/A | 78 | Acetic acid | 38 | 87 | ||
| L-Asparagine | 78 | N/A | Formic acid | 0 | 82 | ||
| D-Aspartic Acid | N/A | 73 | Fumaric acid | 78 | N/A | ||
| L-Histidine | N/A | 89 | Glyoxylic acid | 0 | N/A | ||
| L-Glutamic acid | 82 | 93 | α-Hydroxybutyric acid | 0 | 13 | ||
| L-Glutamine | 100 | N/A | β-Hydroxybutyric acid | 24 | 88 | ||
| Glycyl-L-aspartic acid | 72 | N/A | Itaconic acid | 0 | N/A | ||
| Glycyl-L-glutamine | 100 | N/A | α-Ketobutyric acid | 0 | 2 | ||
| Glycyl-L-methionine | 0 | N/A | α-Ketovaleric acid | 0 | N/A | ||
| Glycyl-L-proline | 100 | 94 | α-Ketoglutaric acid | N/A | 60 | ||
| L-Methionine | 0 | N/A | D,L-Lactic acid | 60 | N/A | ||
| L-Phenylalanine | 0 | N/A | L-Lactic acid | 100 | 94 | ||
| L-pyroglutamic acid | N/A | 79 | D-Malic acid | 0 | 68 | ||
| D-Serine | N/A | 10 | L-Malic acid | 100 | 95 | ||
| L-Serine | 100 | 94 | Propionic acid | 0 | 5 | ||
| L-Threonine | 0 | N/A | Pyruvic acid | 100 | N/A | ||
| L-Valine | 0 | N/A | D-Saccharic acid | 100 | 94 | ||
| L-Valine + L-aspartic acid | 100 | N/A | Succinic acid | 100 | N/A | ||
| Carbohydrate | N-Acetyl-D-galactosamine | 0 | 44 | m-Tartaric acid | 100 | N/A | |
| N-Acetyl-D-glucosamine | 100 | 94 | Mucic Acid | N/A | 93 | ||
| N-Acetyl-D-mannosamine | 100 | 94 | Quinic Acid | N/A | 68 | ||
| Adonitol | 0 | N/A | Citric Acid | N/A | 94 | ||
| D-Arabitol | 0 | 67 | Acetoacetic Acid | N/A | 55 | ||
| Arbutin | 82 | N/A | Miscellaneous | Amygdalin | 0 | N/A | |
| D-Cellobiose | 100 | 92 | Glycerol | 24 | 87 | ||
| Dulcitol | 16 | N/A | D,L-α-Glycerol phosphate | 100 | N/A | ||
| Erythritol | 0 | N/A | Glucose-1-phosphate | 100 | N/A | ||
| D-Fructose | 100 | 85 | Glucose-6-phosphate | 100 | 95 | ||
| D-Fucose | N/A | 45 | Frucose-6-phosphate | N/A | 93 | ||
| L-Fucose | 0 | 67 | D-Salicin | 78 | 85 | ||
| D-Galactose | 100 | 84 | D,L-Lactic acid methyl ester | 0 | 78 | ||
| Gentiobiose | 78 | 88 | Pyruvic acid methyl ester | 100 | 89 | ||
| α-D-Glucose | 100 | 90 | Succinic acid mono-methyl ester | 56 | N/A | ||
| m-Inositol | 60 | 93 | Urocanic acid | 100 | N/A | ||
| α-D-Lactose | 100 | 86 | 2′-Deoxy adenosine | 100 | N/A | ||
| Lactulose | 58 | N/A | Inosine | 80 | 92 | ||
| Maltose | 100 | 83 | Thymidine | 100 | N/A | ||
| Maltotriose | 100 | N/A | Uridine | 100 | N/A | ||
| D-Mannitol | 100 | 87 | Thymidine-5′-monophosphate | 24 | N/A | ||
| D-Mannose | 100 | 86 | Uridine-5′-monophosphate | 78 | N/A | ||
| D-Melezitose | 0 | N/A | N-Acetyl Neuraminic Acid | N/A | 18 | ||
| D-Melibiose | 100 | 86 | L-Galactonic Acid Lactone | N/A | 94 | ||
| 3-Methyl-D-glucose | 32 | 39 | p-Hydroxy-Phenylacetic Acid | N/A | 92 | ||
| α-Methyl-D-galactose | 44 | N/A | Bromo-Succinic Acid | N/A | 92 | ||
| β-Methyl-D-galactose | 24 | N/A | Tween 40 | N/A | 51 | ||
| α-Methyl-D-glucoside | 24 | N/A | γ-Amino-Butryric Acid | N/A | 10 | ||
| β-Methyl-D-glucoside | 100 | 85 | Polymer | α-Cyclodextrin | 0 | N/A | |
| Palatinose | 100 | N/A | β-Cyclodextrin | 0 | N/A | ||
| D-Raffinose | 100 | 87 | Dextrin | 100 | 84 | ||
| L-Rhamnose | 78 | 89 | Gelatin | N/A | 69 | ||
| D-Sorbitol | 100 | 91 | Pectin | N/A | 82 | ||
| Stachyose | 100 | 88 |
| Medium | Metabolites | Biohydrogen Potential (mmol/L) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Butyric Acid | Acetic Acid | Propionic Acid | Lactic Acid | Formic Acid | Ethanol | 2,3-butandiol | Other | ||
| RCM | + | + | + | − | − | + | + | Ethyl isopropyl ether | 45.6 ± 4.9 |
| Glucose and xylose | + | + | − | − | − | + | − | − | 46.1 ± 3.5 |
| AFEX OPEFB hydrolysate | + | + | − | − | − | + | − | Acetamide Ethyl isopropyl ether | 50.6 ± 5.3 |
| DAA OPEFB molasses | + | + | + | − | − | + | + | 2-furan methanol, Ethyl isopropyl ether | 35.3 ± 3.7 |
| Soluble Compounds | AFEX OPEFB Hydrolysate | DAA OPEFB Molasses |
|---|---|---|
| Initial Total Carbohydrates | 7.1 ± 0.1 | 7.1 ± 0.1 |
| Oligosaccharide: | ||
| Glucan | n.d. | 0.6 ± 0.1 |
| Xylan | n.d. | 2.9 ± 0.1 |
| Monosaccharide: | ||
| Glucose | 4.5 ± 0.2 | 0.2 ± 0.1 |
| Xylose | 2.3 ± 0.1 | 3.2 ± 0.1 |
| Furfural | n.d. | 0.51 ± 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luthfi, A.A.I.; Abdul, P.M.; Jahim, J.M.; Engliman, N.S.; Jamali, N.S.; Tan, J.P.; Manaf, S.F.A.; Sajab, M.S.; Bukhari, N.A. Isolation and Characterization of Biohydrogen-Producing Bacteria for Biohydrogen Fermentation Using Oil Palm Biomass-Based Carbon Source. Appl. Sci. 2023, 13, 656. https://doi.org/10.3390/app13010656
Luthfi AAI, Abdul PM, Jahim JM, Engliman NS, Jamali NS, Tan JP, Manaf SFA, Sajab MS, Bukhari NA. Isolation and Characterization of Biohydrogen-Producing Bacteria for Biohydrogen Fermentation Using Oil Palm Biomass-Based Carbon Source. Applied Sciences. 2023; 13(1):656. https://doi.org/10.3390/app13010656
Chicago/Turabian StyleLuthfi, Abdullah Amru Indera, Peer Mohamed Abdul, Jamaliah Md Jahim, Nurul Sakinah Engliman, Nur Syakina Jamali, Jian Ping Tan, Shareena Fairuz Abdul Manaf, Mohd Shaiful Sajab, and Nurul Adela Bukhari. 2023. "Isolation and Characterization of Biohydrogen-Producing Bacteria for Biohydrogen Fermentation Using Oil Palm Biomass-Based Carbon Source" Applied Sciences 13, no. 1: 656. https://doi.org/10.3390/app13010656
APA StyleLuthfi, A. A. I., Abdul, P. M., Jahim, J. M., Engliman, N. S., Jamali, N. S., Tan, J. P., Manaf, S. F. A., Sajab, M. S., & Bukhari, N. A. (2023). Isolation and Characterization of Biohydrogen-Producing Bacteria for Biohydrogen Fermentation Using Oil Palm Biomass-Based Carbon Source. Applied Sciences, 13(1), 656. https://doi.org/10.3390/app13010656