

Comparative Study of the Anticancer Effects of Selenium Nanoparticles and Selenium Nanorods: Regulation of Ca2+ Signaling, ER Stress and Apoptosis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Morphology of Selenium Nanorods
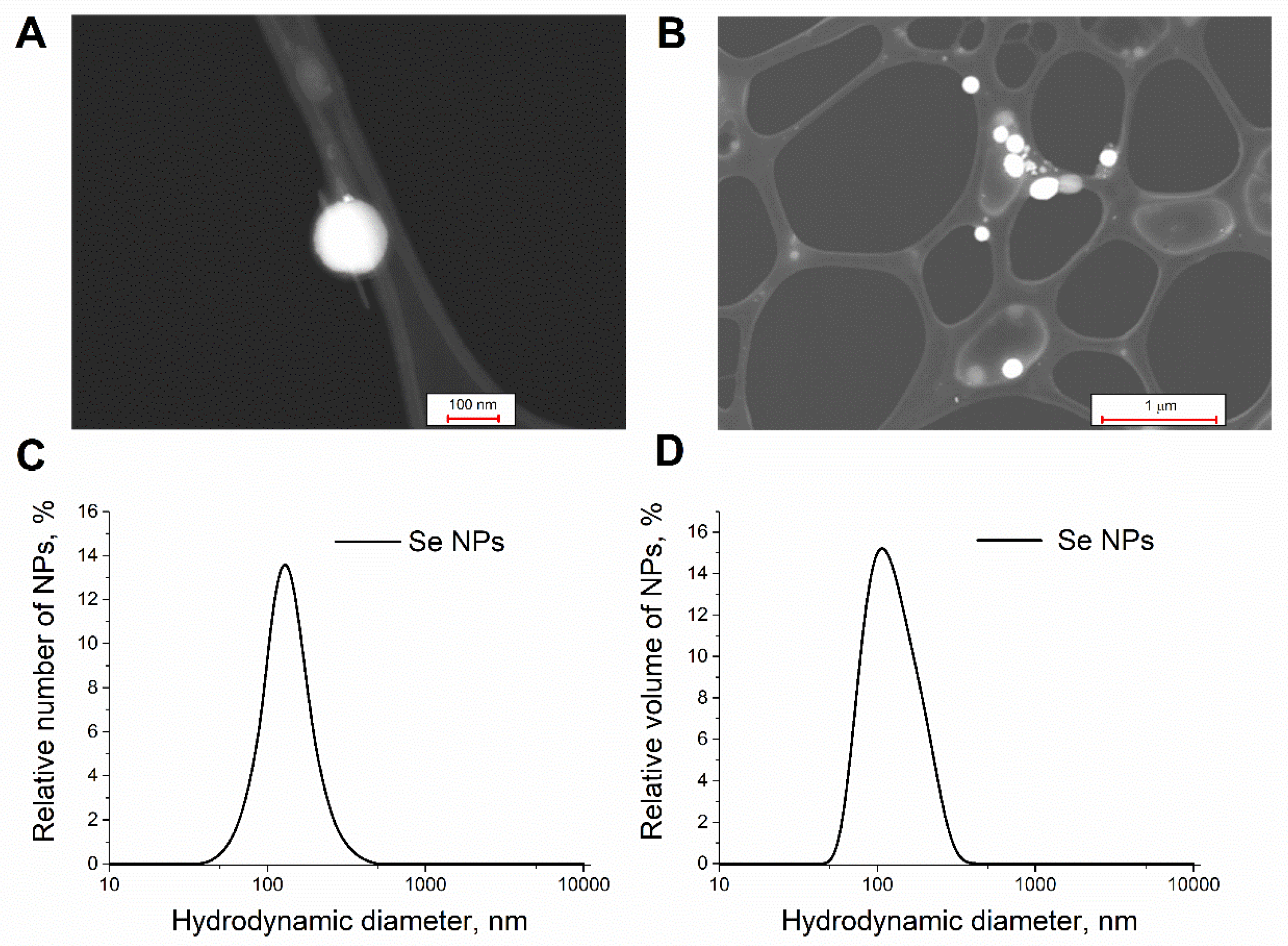
2.2. Synthesis and Morphology of Selenium Nanoparticles
2.3. Cell Culture
2.4. Assessment of Cell Viability
2.5. Fluorescent Ca2+ Imaging
2.6. Extraction of RNA and Real-Time Polymerase Chain Reaction (RT-qPCR)
3. Results
3.1. Comparative Analysis of the Selenium Nanorods (SeNrs) and Selenium Nanoparticles (SeNPs) Effects on the Induction of Ca2+ Signals in the A-172 Human Glioblastoma Cells and L-929 Mouse Fibroblasts
3.2. Comparative Analysis of the Proapoptotic Effect of Selenium Nanofibers and Selenium Nanoparticles on Human Glioblastoma Cells A-172 and Mouse Fibroblast Cells L-929

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h Pre-Incubation | 48 h Pre-Incubation | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | |
| Viable | 100% | 30% | 43% | 23% | 100% | 12% | 0% | 0% |
| Early apoptotic | 0% | 15% | 5% | 10% | 0% | 19% | 24% | 26% |
| Apoptotic | 0% | 55% | 52% | 67% | 0% | 69% | 73% | 69% |
| Necrosis | 0% | 0% | 0% | 0% | 0% | 0% | 3% | 5% |
3.3. Effects of SeNrs and SeNPs on Expression Patterns of Genes Encoding Selenoproteins and Selenium-Containing Proteins, Genes Regulating Apoptosis and Endoplasmic Reticulum Stress
3.4. The Effect of Various SeNrs and SeNPs Concentrations on the Ca2+ Capacity of the Endoplasmic Reticulum of Human Glioblastoma Cells and Mouse Fibroblasts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Minich, W.B. Selenium Metabolism and Biosynthesis of Selenoproteins in the Human Body. Biochemistry 2022, 87, S168–S177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, X.; Wei, Y. Selenium and Selenoproteins in Health. Biomolecules 2023, 13, 799. [Google Scholar] [CrossRef] [PubMed]
- Serov, D.A.; Khabatova, V.V.; Tikhonova, I.V.; Reut, V.E.; Pobedonostsev, R.V.; Astashev, M.E. Study of the Effects of Selenium Nanoparticles and Their Combination with Immunoglobulins on the Survival and Functional State of Polymorphonuclear Cells. Opera Med. Physiol. 2022, 4, 137–159. [Google Scholar]
- Mal’tseva, V.N.; Gudkov, S.V.; Turovsky, E.A. Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo. Int. J. Mol. Sci. 2022, 23, 13651. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, M.H.; Mahdavi, M.; Varastehmoradi, B.; Faramarzi, M.A.; Shahverdi, A.R. The immunostimulatory effect of biogenic selenium nanoparticles on the 4T1 breast cancer model: An in vivo study. Biol. Trace Elem. Res. 2012, 149, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esquivias, F.; Gutiérrez-Angulo, M.; Pérez-Larios, A.; Sánchez-Burgos, J.A.; Becerra-Ruiz, J.S.; Guzmán-Flores, J.M. Anticancer Activity of Selenium Nanoparticles In Vitro Studies. Anticancer Agents Med. Chem. 2022, 22, 1658–1673. [Google Scholar] [CrossRef]
- Ferro, C.; Florindo, H.F.; Santos, H.A. Selenium Nanoparticles for Biomedical Applications: From Development and Characterization to Therapeutics. Adv. Healthc. Mater. 2021, 10, 2100598. [Google Scholar] [CrossRef]
- Deepasree, K.; Subhashree, V. Therapeutic potential of selenium nanoparticles. Front. Nanotechnol. 2022, 4, 1042338. [Google Scholar]
- Jiang, X.; Kemal, L.; Yu, A. Silver-induced growth of selenium nanowires in aqueous solution. Mater. Lett. 2007, 61, 2584–2588. [Google Scholar] [CrossRef]
- Sarkar, J.; Saha, S.; Dey, P.; Acharya, K. Production of selenium nanorods by phytopathogen, Alternaria alternata. Adv. Sci. Lett. 2012, 10, 111–114. [Google Scholar] [CrossRef]
- Kumar, A.; Sevonkaev, I.; Goia, D.V. Synthesis of selenium particles with various morphologies. J. Colloid Interface Sci. 2014, 416, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xie, Y.; Zhang, S.; Li, F. Ultra-thin trigonal selenium nanoribbons developed from series-wound beads. Adv. Mater. 2004, 16, 649–653. [Google Scholar] [CrossRef]
- Yin, H.; Xu, Z.; Bao, H.; Bai, J.; Zheng, Y. Single crystal trigonal selenium nanoplates converted from selenium nanoparticles. Chem. Lett. 2005, 34, 122–123. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, D.; Ji, Y.; Ma, X.; Xu, J.; Que, D. Selenium nanotubes synthesized by a novel solution phase approach. J. Phys. Chem. B 2004, 108, 1179–1182. [Google Scholar] [CrossRef]
- Zhang, B.; Dai, W.; Ye, X.; Zuo, F.; Xie, Y. Photothermally assisted solution-phase synthesis of microscale tubes, rods, shuttles, and an urchin-like assembly of single-crystalline trigonal selenium. Angew. Chem. Int. Ed. Engl. 2006, 45, 2571–2574. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Ran, F.; Cui, Y.; Liu, C.; Zhao, Q.; Gao, Y.; Wang, D.; Wang, S. A comparison between sphere and rod nanoparticles regarding their in vivo biological behavior and pharmacokinetics. Sci. Rep. 2017, 7, 4131. [Google Scholar] [CrossRef]
- Drexler, H.G.; Uphoff, C.C. Mycoplasma contamination of cell cultures: Incidence, sources, effects, detection, elimination, prevention. Cytotechnology 2002, 39, 75–90. [Google Scholar] [CrossRef]
- Gaidin, S.G.; Turovskaya, M.V.; Gavrish, M.S.; Babaev, A.A.; Mal’tseva, V.N.; Blinova, E.V.; Turovsky, E.A. The selective BDNF overexpression in neurons protects neuroglial networks against OGD and glutamate-induced excitotoxicity. Int. J. Neurosci. 2020, 130, 363–383. [Google Scholar] [CrossRef]
- Schmid, I.; Uittenbogaart, C.; Jamieson, B.D. Live-cell assay for detection of apoptosis by dual-laser flow cytometry using Hoechst 33342 and 7-amino-actino-mycin D. Nat. Protoc. 2007, 2, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Turovsky, E.A.; Varlamova, E.G. Mechanism of Ca2+-Dependent Pro-Apoptotic Action of Selenium Nanoparticles, Mediated by Activation of Cx43 Hemichannels. Biology 2021, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Lunin, S.M.; Novoselova, E.G.; Glushkova, O.V.; Parfenyuk, S.B.; Kuzekova, A.A.; Novoselova, T.V.; Sharapov, M.G.; Mubarakshina, E.K.; Goncharov, R.G.; Khrenov, M.O. Protective effect of exogenous peroxiredoxin 6 and thymic peptide thymulin on BBB conditions in an experimental model of multiple sclerosis. Arch. Biochem. Biophys. 2023, 746, 109729. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Turovsky, E.A.; Babenko, V.A.; Plotnikov, E.Y. The Mechanisms Underlying the Protective Action of Selenium Nanoparticles against Ischemia/Reoxygenation Are Mediated by the Activation of the Ca2+ Signaling System of Astrocytes and Reactive Astrogliosis. Int. J. Mol. Sci. 2021, 22, 12825. [Google Scholar] [CrossRef] [PubMed]
- Khabatova, V.V.; Serov, D.A.; Tikhonova, I.V.; Astashev, M.E.; Nagaev, E.I.; Sarimov, R.M.; Matveyeva, T.A.; Simakin, A.V. Selenium Nanoparticles Can Influence the Immune Response Due to Interactions with Antibodies and Modulation of the Physiological State of Granulocytes. Pharmaceutics 2022, 14, 2772. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Gudkov, S.V.; Plotnikov, E.Y.; Turovsky, E.A. Size-Dependent Cytoprotective Effects of Selenium Nanoparticles during Oxygen-Glucose Deprivation in Brain Cortical Cells. Int. J. Mol. Sci. 2022, 23, 7464. [Google Scholar] [CrossRef] [PubMed]
- Danese, A.; Leo, S.; Rimessi, A.; Wieckowski, M.R.; Fiorica, F.; Giorgi, C.; Pinton, P. Cell death as a result of calcium signaling modulation: A cancer-centric prospective. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119061. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Khabatova, V.V.; Gudkov, S.V.; Turovsky, E.A. Ca2+-Dependent Effects of the Selenium-Sorafenib Nanocomplex on Glioblastoma Cells and Astrocytes of the Cerebral Cortex: Anticancer Agent and Cytoprotector. Int. J. Mol. Sci. 2023, 24, 2411. [Google Scholar] [CrossRef]
- Alifieris, C.; Trafalis, D.T. Glioblastoma multiforme: Pathogenesis and treatment. Pharmacol. Ther. 2015, 152, 63–82. [Google Scholar] [CrossRef]
- Davis, M.E. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.A. Nanotechnology approaches for the regeneration and neuroprotection of the central nervous system. Surg. Neurol. 2005, 63, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Drug transport across the blood-brain barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.; Yasser, M.M.; Sholkamy, E.N.; Ali, A.M.; Mehanni, M.M. Anticancer activity of biostabilized selenium nanorods synthesized by Streptomyces bikiniensis strain Ess_amA-1. Int. J. Nanomed. 2015, 10, 3389–3401. [Google Scholar]
- Varlamova, E.G.; Goltyaev, M.V.; Simakin, A.V.; Gudkov, S.V.; Turovsky, E.A. Comparative Analysis of the Cytotoxic Effect of a Complex of Selenium Nanoparticles Doped with Sorafenib, “Naked” Selenium Nanoparticles, and Sorafenib on Human Hepatocyte Carcinoma HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 6641. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Su, J.; Zhao, Z.; Zheng, W.; Wu, H.; Zhang, Y.; Chen, T. Differential effects of amino acid surface decoration on the anticancer efficacy of selenium nanoparticles. Dalton Trans. 2014, 43, 1854–1861. [Google Scholar] [CrossRef]
- Aboul-Fadl, T. Selenium derivatives as cancer preventive agents. Curr. Med. Chem.-Anti-Cancer Agents 2005, 5, 637–652. [Google Scholar] [CrossRef]
- Baskar, G.; Lalitha, K.; Aiswarya, R.; Naveenkumar, R. Synthesis, characterization and synergistic activity of cerium-selenium nanobiocomposite of fungal l-asparaginase against lung cancer. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 93, 809–815. [Google Scholar] [CrossRef]
- Shigemi, Z.; Manabe, K.; Hara, N.; Baba, Y.; Hosokawa, K.; Kagawa, H.; Watanabe, T.; Fujimuro, M. Methylseleninic acid and sodium selenite induce severe ER stress and subsequent apoptosis through UPR activation in PEL cells. Chem.-Biol. Interact. 2017, 266, 28–37. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Goltyaev, M.V.; Turovsky, E.A. The Role of Selenoproteins SELENOM and SELENOT in the Regulation of Apoptosis, ER Stress, and Calcium Homeostasis in the A-172 Human Glioblastoma Cell Line. Biology 2022, 11, 811. [Google Scholar] [CrossRef] [PubMed]
- Bruskov, V.I.; Karmanova, E.E.; Chernikov, A.V.; Usacheva, A.M.; Ivanov, V.E.; Emel’yanenko, V.I. Formation of Hydrated Electrons in Water under Thermal Electromagnetic Exposure. Phys. Wave Phenom. 2021, 29, 94–97. [Google Scholar] [CrossRef]
- Yu, B.; Liu, T.; Du, Y.; Luo, Z.; Zheng, W.; Chen, T. X-ray-responsive selenium nanoparticles for enhanced cancer chemo-radiotherapy. Colloids Surf. B Biointerfaces 2016, 139, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.; Weissman, A. The Unfolded Protein Response, Degradation from the Endoplasmic Reticulum, and Cancer. Genes Cancer 2010, 1, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.; Ron, D. Endoplasmic Reticulum Stress Signaling in Disease. Physiol. Rev. 2006, 86, 1133–1149. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Yan, S.; Dou, X.; Song, X.; Chang, J.; Pi, S.; Zhang, X.; Xu, C. Biogenic Selenium Nanoparticles Alleviate Intestinal Epithelial Barrier Damage through Regulating Endoplasmic Reticulum Stress-Mediated Mitophagy. Oxidative Med. Cell. Longev. 2022, 2022, 3982613. [Google Scholar] [CrossRef] [PubMed]
- Grosskreutz, J.; Van Den Bosch, L.; Keller, B.U. Calcium dysregulation in amyotrophic lateral sclerosis. Cell Calcium 2010, 47, 165–174. [Google Scholar] [CrossRef]
- Andjelíc, S.; Khanna, A.; Suthanthiran, M.; Nikolić-Zugić, J. Intracellular Ca2+ elevation and cyclosporin A synergistically induce TGF-beta 1-mediated apoptosis in lymphocytes. J. Immunol. 1997, 158, 2527–2534. [Google Scholar] [CrossRef]
- Crompton, M. The mitochondrial permeability transition pore and its role in cell death. Biochem. J. 1999, 341, 233–249. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, J. Thapsigargin triggers cardiac contractile dysfunction via NADPH oxidase-mediated mitochondrial dysfunction: Role of Akt dephosphorylation. Free Radic. Biol. Med. 2011, 51, 2172–2184. [Google Scholar] [CrossRef]









| Gene Name | Forward Primer 5′–>3′ | Reverse Primer 5′–>3′ |
|---|---|---|
| GAPDH | ACATCGCTCAGACACCATG | GCCAGTGAGCTTCCCGTT |
| SELENOT | TCTCCTAGTGGCGGCGTC | GTCTATATATTGGTTGAGGGAGG |
| SELENOM | AGCCTCCTGTTGCCTCCGC | AGGTCAGCGTGGTCCGAAG |
| SELENOF | GGAGGAAGCACAATTTGAAACC | TATGCGTTCCAACTTTTCACTC |
| SELENOK | TTTACATCTCGAACGGACAAG | CAGCCTTCCACTTCTTGATG |
| SELENOS | TGGGACAGCATGCAAGAAG | GCGTCCAGGTCTCCAGG |
| SELENON | TGATCTGCCTGCCCAATG | TCAGGAACTGCATGTAGGTGG |
| DIO2 | AGCTTCCTCCTCGATGCC | AAAGGAGGTCAAGTGGCTG |
| CHOP | GCTCTGATTGACCGAATGG | TCTGGGAAAGGTGGGTAGTG |
| GADD34 | CTCCGAGAAGGTCACTGTCC | GACGAGCGGGAAGGTGTGG |
| PUMA | CAGATATGCGCCCAGAGAT | CCATTCGTGGGTGGTCTTC |
| BIM | GGACGACCTCAACGCACAGTACGAG | GTAAGGGCAGGAGTCCCA |
| CAS–3 | GCATTGAGACAGACAGTGGTG | AATAGAGTTCTTTTGTGAGCATG |
| CAS–4 | CACGCCTGGCTCTCATCATA | TAGCAAATGCCCTCAGCG |
| BAX | GGGCTGGACATTGGACTTC | AACACAGTCCAAGGCAGCTG |
| BAK | GAGAGTGGCATCAATTGGGG | CAGCCACCCCTCTGTGCAATCCA |
| BCL-2 | GGTGAACTGGGGGAGGATTG | AGCCAGGAGAAATCAAACAGAG |
| ATF–4 | GTGTTCTCTGTGGGTCTGCC | GACCCTTTTCTTCCCCCTTG |
| ATF–6 | AACCCTAGTGTGAGCCCTGC | GTTCAGAGCACCCTGAAGA |
| XBPu | ACTCAGACTACGTGCACCTC | GTCAATACCGCCAGAATCC |
| XBPs | CTGAGTCCGCAGCGGTGCAGG | GGTCCAAGTTGTCCAGAATG |
| GPX1 | AAGATCCAACCCAAGGGCAAG | CATGAGTGCCGGTGGAAGG |
| GPX4 | AAGATCCAACCCAAGGGCAAG | CATGAGTGCCGGTGGAAGG |
| TXNRD1 | CAACAAATGTTATGCAAAAATAATC | ACACTGGGGCTTAACCTCAG |
| TXNRD3 | CTCTTTAGAAAAGTGTGATTATATT | GCCCACATTTCATTGCAGCTG |
| Nf-kB | TACTTTCTCACTTTTTGCCCAC | GGTCTACAGGAAGGCGTGG |
| Gene Name | Forward Primer 5′–>3′ | Reverse Primer 5′–>3′ |
|---|---|---|
| GAPDH | AAGGTGGTGAAGCAGGCATC | CTCTTGCTCAGTGTCCTTGC |
| SELENOT | TGATTGAGAACCAGTGTATGTC | GGTACAACGAGCCTGCCAAG |
| SELENOM | CGCCTAAAGGAGGTGAAGGC | CTTGCGGTAGAAGCCGAGCTC |
| SELENOF | AGGGTGCTGTCAGGAAGAAG | CGTTCCAACTTCTCGCTCAG |
| SELENOK | GAAGAGGGCCACCAGGAAAC | GGAATTCCCAGCATGACCTC |
| SELENOS | GGACCAAGCCGAGACTGTTC | CTTCTTGCATGCTGTCCCAC |
| SELENON | AAGATGGCTTCCTAGGGGTC | CTGAGGGGCAAAGCGGGTC |
| DIO2 | GCTTATCTCTGCCCCCATTG | CACACATAAACGACCTCCTTC |
| CHOP | CAGCTGGGAGCTGGAAGCCTG | GACCACTCTGTTTCCGTTTCC |
| GADD34 | GAGTCCCATGAAGAGATTGTAC | ACCAGCCCAGCAGCACTTAG |
| PUMA | TGAAGATCTGCGCCGGGAG | GAGAGGGACATGACGCGTG |
| BIM | AATGGCCGGCTATGGATGATG | GCCAATTGGGTTCACTGTCTG |
| CAS–3 | GACCCGTCCTTTGAATTTCTC | CTCTTCATCATTCAGGCCTGC |
| CAS–4 | TTTTCTTTTCTTCTCAGCTACAG | TGTTGGTGTTATCATTTGGAGG |
| BAX | TAAAGTGCCCGAGCTGATCAGAAC | CTTCCCAGCCACCCTGGTCTT |
| BAK | CAGATGGATCGCACAGAGAG | GCGTCTTTGCCCTGGGGAG |
| BCL-2 | GGTGAACTGGGGGAGGATTG | AGCCAGGAGAAATCAAACAGAG |
| ATF–4 | TCGGGTTTGGGGGCTGAAG | AAACAGAGCATCGAAGTCAAAC |
| ATF–6 | AGGAGGGGAGATACGTTTTAC | CGAGGAGCTTTTGATGTGGAG |
| XBPu | GAGTCCGCAGCAGAGTCCGCAGC | GGAGGCTGGTAAGGAACTAG |
| XBPs | AGTCCGCAGCACAGCAGGT | AGAGAAAGGGAGGCTGGTAAG |
| GPX1 | GGGGAGCCTGTGAGCCTGG | GGACGTACTTGAGGGAATTC |
| GPX4 | GATGAAAGTCCAGCCCAAGG | GAAGGCTCCAGGGGTCACAG |
| TXNRD1 | CAACAAATGTTATGCAAAAATAATC | ACACTGGGGCTTAACCTCAG |
| TXNRD3 | CTCTTTAGAAAAGTGTGATTATATT | GCCCACATTTCATTGCAGCTG |
| Nf-kB | AAGTGCAAAGGAAACGCCAGAA | ACTACCGAACATGCCTCCACCA |
| 24 h Pre-Incubation | 48 h Pre-Incubation | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | |
| Viable | 100% | 76% | 37% | 18% | 100% | 39% | 12% | 0% |
| Early apoptotic | 0% | 6% | 8% | 2% | 0% | 11% | 17% | 4% |
| Apoptotic | 0% | 18% | 47% | 59% | 0% | 47% | 57% | 44% |
| Necrosis | 0% | 0% | 8% | 21% | 0% | 3% | 14% | 52% |
| 24 h Pre-Incubation | 48 h Pre-Incubation | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | |
| Viable | 100% | 89% | 94% | 96% | 100% | 91% | 89% | 86% |
| Early apoptotic | 0% | 11% | 6% | 4% | 0% | 9% | 11% | 14% |
| Apoptotic | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| Necrosis | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| 24 h Pre-Incubation | 48 h Pre-Incubation | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | Control | 1 µg/mL | 5 µg/mL | 10 µg/mL | |
| Viable | 100% | 100% | 98% | 94% | 100% | 93% | 81% | 51% |
| Early apoptotic | 0% | 0% | 2% | 6% | 0% | 7% | 11% | 28% |
| Apoptotic | 0% | 0% | 0% | 0% | 0% | 0% | 8% | 16% |
| Necrosis | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varlamova, E.G.; Baimler, I.V.; Gudkov, S.V.; Turovsky, E.A. Comparative Study of the Anticancer Effects of Selenium Nanoparticles and Selenium Nanorods: Regulation of Ca2+ Signaling, ER Stress and Apoptosis. Appl. Sci. 2023, 13, 10763. https://doi.org/10.3390/app131910763
Varlamova EG, Baimler IV, Gudkov SV, Turovsky EA. Comparative Study of the Anticancer Effects of Selenium Nanoparticles and Selenium Nanorods: Regulation of Ca2+ Signaling, ER Stress and Apoptosis. Applied Sciences. 2023; 13(19):10763. https://doi.org/10.3390/app131910763
Chicago/Turabian StyleVarlamova, Elena G., Ilya V. Baimler, Sergey V. Gudkov, and Egor A. Turovsky. 2023. "Comparative Study of the Anticancer Effects of Selenium Nanoparticles and Selenium Nanorods: Regulation of Ca2+ Signaling, ER Stress and Apoptosis" Applied Sciences 13, no. 19: 10763. https://doi.org/10.3390/app131910763
APA StyleVarlamova, E. G., Baimler, I. V., Gudkov, S. V., & Turovsky, E. A. (2023). Comparative Study of the Anticancer Effects of Selenium Nanoparticles and Selenium Nanorods: Regulation of Ca2+ Signaling, ER Stress and Apoptosis. Applied Sciences, 13(19), 10763. https://doi.org/10.3390/app131910763