Strategies for Enhancing Extracellular Electron Transfer in Environmental Biotechnology: A Review


Abstract
:1. Introduction
2. Mechanism of Extracellular Electron Transport in Microorganisms
2.1. Direct Electron Transfer (DET)
2.2. Indirect Electron Transfer (IET)
2.3. Factors Affecting EET Efficiency
3. Strategies for Enhancing EET in Environmental Biotechnology
3.1. Genetic Engineering Approaches
- (a)
- Overexpression of key EET genes
- (b)
- Exogenous EET
3.2. Biofilm Formation and Optimization
- (a)
- Biofilm matrix engineering
- (b)
- Quorum sensing manipulation for biofilm development
3.3. Electron Shuttles and Mediators
- (a)
- Use of natural or synthetic compounds as electron shuttles
- (b)
- The use of conductive nanowires
3.4. Electrode Modifications and Design
3.5. External Electricity or Electric Field
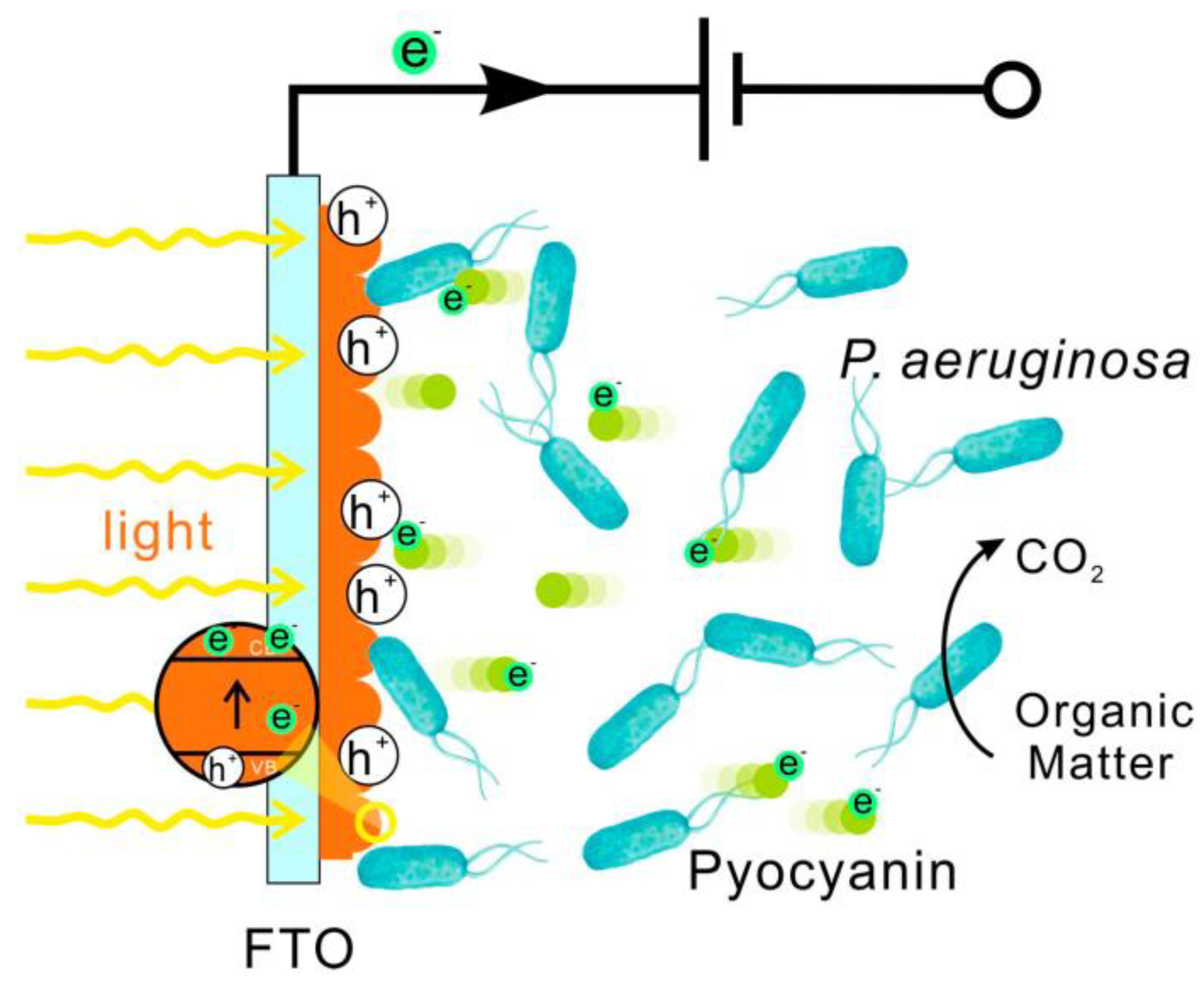
3.6. Enhancing EET by Photo
3.7. Enhancing EET by Magnetism
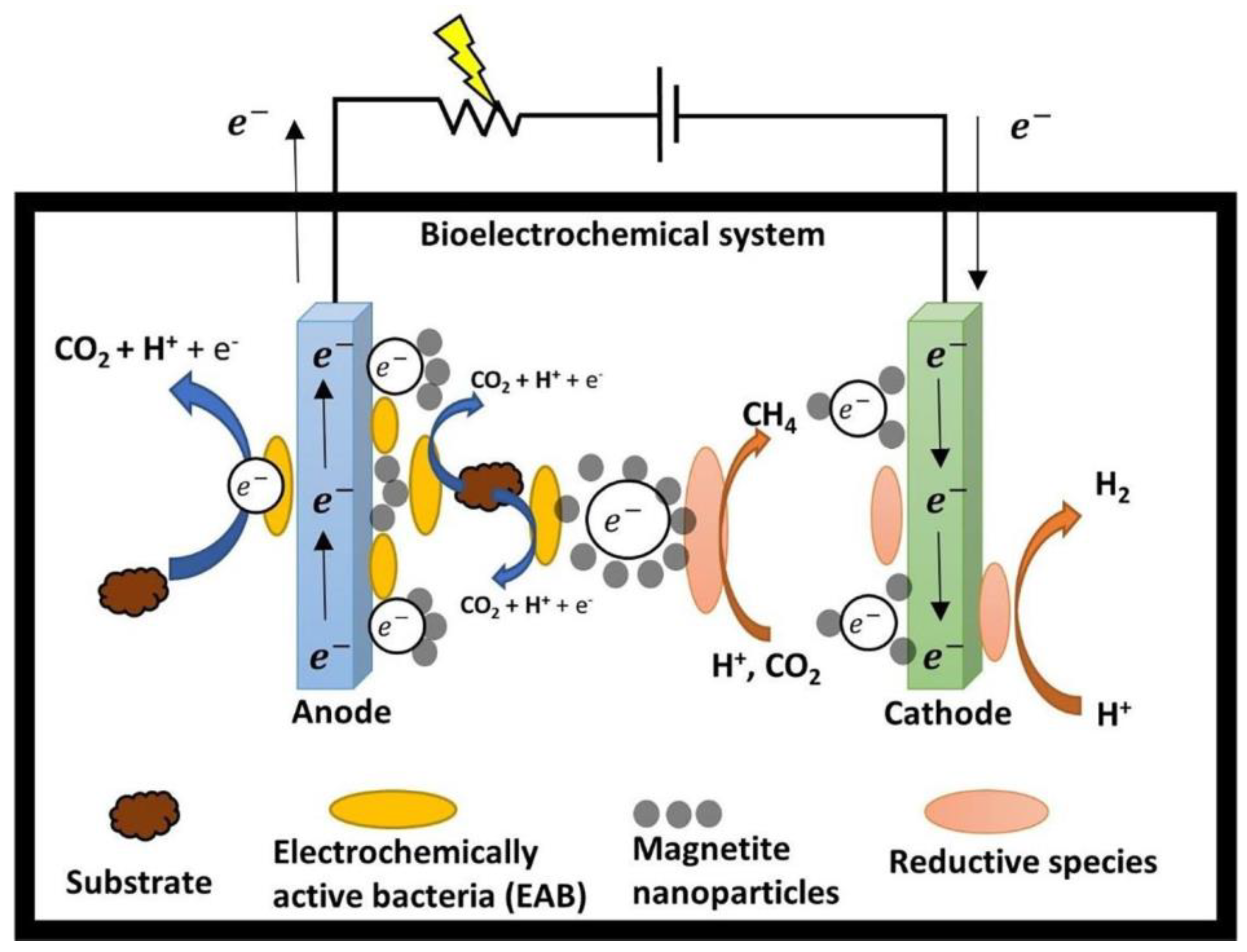
3.8. Integrated Strategies for Enhancing EET
- Additive manufacturing (AM): AM, commonly referred to as 3D printing, is a rapidly advancing technological innovation that has made significant contributions to MFC technology in recent times.A study by Calignano et al. [97] (Figure 5) explored the limits of AM technology in the creation of a powering device that is entirely based on AM. The study focused on utilizing low-density and open porosities to accommodate microorganisms, as well as developing systems that are easily fueled continually and operate securely. A maximum energy recovery of approximately 3 kWh m−3 per day was achieved, which is capable of supplying power to sensing devices and low-energy electronic gadgets. This facilitates data conveyance and analysis from remote and challenging situations. AM facilitates the design for the assembly of MFCs, thereby mitigating mistakes as a result of human intervention and expediting the installation procedure. In a study by Zawadzki et al. [98], AM techniques were also employed to construct an MFC with the incorporation of an exchangeable membrane slot. This enabled them to examine the effects of different separator membranes. The utilization of AM in the production of tango membranes has demonstrated promising prospects for membrane manufacturing and power-generating capabilities. However, it is worth noting that the power output achieved (0.92 μW) by this method was relatively low in comparison to the conventional cation exchange membrane technology (11.39 μW). Similarly, Slate [99] investigated enhancing the power outputs of MFCs with the integration of graphene into electrodes fabricated using 3D printing technology. The electrodes underwent an evaluation to assess their electrical efficiency, chemistry, and appearance, thereby showcasing the prospective application of AM in the production of MFCs. Furthermore, AM technology, such as inkjet printing, was used in a study by Sawa et al. [100]. The research showcased the practicability of employing a readily available commercial inkjet printer for the production of a thin-film paper-based biophotovoltaic cell. This cell comprised a layer of cyanobacterial cells positioned above a conducting surface composed of carbon nanotubes. The researchers discovered that the printed cyanobacteria can produce a continuous electrical current for a duration exceeding 100 h. This phenomenon occurs both in the absence of light, functioning as a “solar bio-battery”, and in the presence of light, functioning as a “bio-solar-panel”. These findings suggest its possible use in low-power devices. The utilization of the inkjet printing technique facilitated precise positioning of the cyanobacteria onto the anode surface, hence facilitating the proximity of the printed cathode. The printing technique was optimized to achieve ideal biofilm layer thicknesses to maximize power generation. The aforementioned authors were pioneers in employing an inkjet printer to print photosynthetic microorganisms, specifically cyanobacteria, onto a bio-photovoltaic (BPV) cell. Consequently, AM offers the potential to utilize diverse materials and 3D printing techniques to develop a comprehensive MFC tailored to different applications. This helps to expedite the optimization process of individual MFC components, leading to enhanced effectiveness and ease in reactor design and consequently, the system will experience functional improvements. AM has the potential to significantly impact the field of biological power generation devices due to its advantages such as decreased lead time, enhanced design flexibility, cost-effectiveness, and expedited prototyping capabilities. In addition, AM offers a shorter route from the research laboratory to large-scale production. Ultimately, the advancements offered by AM can be effectively implemented in several other bioelectronic systems, including biological sensors, implantable biomedical devices, and biocomputing systems [101].
- Artificial Intelligence (AI) and Machine Learning: Numerous research investigations have been undertaken to examine the performance of mathematical models in the field of artificial intelligence (AI). These studies include the works of [102,103,104,105]. A study by de Ramon-Fernande et al. [106] employed an artificial neural network (ANN) computational model to investigate the relationship between power output and flow rate in a ceramic MFC that was supplied with human excreta, specifically urine. The training, comparison, precision forecasting, and time to convergence assessment were performed using three algorithms: the quasi-Newton technique, the Levenberg–Marquardt algorithm, and the conjugate gradient method, all of which are of third order. The Levenberg–Marquardt algorithm had superior performance in comparison with alternative models in terms of precision, with an R-value of 95%. Additionally, it exhibited faster convergence, with a time of 7.8 s. The findings of the study demonstrate that the use of an ANN computational model is both helpful and accurate for predicting energy requirements in ceramic MFCs when subjected to varying flow rates. Additionally, the feasibility of adopting this model for energy forecasting in ceramic MFCs is established. Lesnik and Liu [107] also implemented the utilization of artificial neural network (ANN) models to forecast the biofilm communities and reactor performance of MFCs. The research observed that ANN models that integrated biotic interactions demonstrated superior accuracy in predicting reactor performance outcomes compared to models that did not incorporate such interactions. This finding suggests that data mining and machine-learning techniques can effectively forecast the behavior of microbial communities and the performance of bioelectrochemical systems. In another study by Zhang et al. [108], a four-input ANN model was effectively utilized to accurately forecast the levels of glycerol, 1,3-PDO, and biomass. The proposed model exhibits applicability not only in the realm of software for online measurement of glycerol and 1,3-PDO during the industrialization process but also offers valuable insights for other fermentation processes. In BESs, substrate prediction can also be carried out with AI-based techniques. In a study by Cai et al. [109], six machine learning algorithms were trained and it was discovered that feed substrates could be predicted using information about the microbial community. These algorithms included logistic regression multiclass (GLMNET), random forest (RF), the scalable tree boosting system (XGBOOST), the neural network (NNET), k-nearest neighbor (KNN), and the support vector machine with radial kernel (SVM). The NNET method demonstrated the highest accuracy (93 ± 6%), allowing for the distinction between various feed substrates [110]. Similarly, a study by Leropoulos et al. [111] applied automation by utilizing a robotic platform known as EcoBot. This platform was developed using artificial intelligence techniques derived from artificial evolution. The concept aimed to leverage the use of robots in facilitating evolutionary processes to create functional ecosystems that establish symbiotic relationships between living organisms and artificial life chemistries. This results in an enhancement of energy generation in MFCs. The conversion of a robot platform, derived from an open-source 3D printer, enables its utilization for liquid management and real-time monitoring of experimental outcomes. The outcomes derived from these illustrative instances demonstrate that artificial intelligence has the potential to enhance and evaluate the performance of MFCs. Furthermore, genetic programming (GP) is an additional technique that makes it possible to optimize the AI model’s structure by minimizing errors through the use of algorithms like cross-validation and the Kennard–Stone algorithm [110]. To incorporate novel components into bacterial cells, one can use the de novo protein design technique. The assembly of peptides into protein structure is connected to the design architecture of de novo and the bundling of α-helices into metal-binding proteins is one way to accomplish this. Short gene fragments are integrated into the genes of E. coli in bacterial surface display systems and LamB, OmpA, Lpp-OmpA, and PhoE coli OM proteins coordinate protein integration into the outer membrane cells [112]. Similarly, genetically modifying E. coli was used to facilitate EET by introducing a biosynthetic pathway for Pseudomonas phenazine-1-carboxylic acid [113]. The findings of the study demonstrated that the implementation of this electrochemical system resulted in a notable augmentation in the synthesis of phenazine-1-carboxylic acid, consequently improving the EET efficiency of MFCs. Another versatile algorithm for flexible diagnosis and control is fuzzy logic (FL). This technique is capable of observing data and forecasting system outputs logically. FL is frequently used as an intermediate step to streamline data processing and lessen the negative impact on model accuracy [114].
- Hybrid Systems: The hybrid technique is another system which integrates two treatment systems, resulting in the enhanced performance efficiency of the overall system. It was noted that the implementation of a constructed wetland (CW)-MFC containing activated carbon granules and manganese ore can enhance the biochemical processes involved in N-transformations in the absence of oxygen [115].
4. Challenges and Future Directions
- Diversity of microorganisms: The effectiveness of electrochemically active microbial communities in BESs can be impacted by their diverse features [136]. The coexistence of diverse species can lead to competition for resources and electron donors/acceptors, impacting the efficiency of EET. Syntrophic interactions between different species can also affect EET efficacy. Changes in microbial diversity resulting from variations in environmental circumstances or disturbances, such as pH, temperature, and oxygen levels, can induce modifications in EET. The structure and composition of biofilms on electrode surfaces can also be influenced by microbial diversity. Different microbial species use different methods for electron transfer, and the effectiveness and kinetics of EET can be influenced by the presence of different microbial species that possess unique electron transfer mechanisms. A diverse microbial community can also display a broad spectrum of redox potentials, potentially causing inefficiencies if there is a lack of alignment between the electron donor and acceptor potentials.
- Scaling up: While laboratory-scale studies have demonstrated the potential of BESs for various applications, scaling up these systems for practical implementation poses significant challenges [137], including the design and arrangement of electrodes, achieving uniform current distribution, and integrating nanostructured materials like carbon nanotubes or graphene into larger systems. Ensuring biofilm stability and uniformity across larger electrode surfaces is crucial for efficient performance. Additionally, expanding EET operations requires supplementary energy inputs, ensuring equilibrium between input and output, and considering the financial implications of implementing the technology. Adherence to environmental and safety regulations is also essential for real-world implementation, as the size of systems increases.
- Substrate availability and selectivity: The efficacy of EET processes is contingent upon the presence of appropriate electron acceptors or donors within the surrounding environment [138]. Furthermore, the selection and accessibility of acceptable substrates for bioelectrochemical systems may be constrained. Numerous systems are dependent on intricate organic substrates or wastewater, the accessibility and economic viability of which may not be generally prevalent. In the context of bioremediation applications, the utilization of EET by microorganisms to break down pollutants relies heavily on the presence of electron acceptors, specifically metal oxides or electrodes, which play a vital role in supporting and maintaining microbial activity. The presence of a limited amount of substrate can result in partial degradation of pollutants, hence diminishing the overall efficiency of remediation processes. Likewise, within MFCs, the process of microbial oxidation of organic materials to produce electricity via EET can be hindered by inadequate substrate availability, resulting in diminished power generation and compromised overall MFC efficiency. Additionally, there is the possibility of substrate competition, when microorganisms exhibit a preference for other pathways instead of EET, resulting in a decrease in the efficiency of electron transfer.
- Biofouling and system stability: The occurrence of biofouling results in the development of insulating layers on the surfaces of electrodes [139], and this can impede electron transmission between microbes and electrodes. Consequently, this results in instability in bioelectrochemical systems due to variations in biofilm thickness, microbial composition, and detachment–reattachment dynamics. Biofouling can also deteriorate electrode materials, necessitating frequent replacements. Effective management and prevention of biofouling often require increased maintenance efforts, which can be costly and result in logistical complexities in the practical implementation of BESs.
- Integration with renewable energy systems: Renewable energy sources, like solar or wind power, can be integrated with bioelectrochemical systems to improve their overall sustainability and energy efficiency [140]. Nevertheless, there are obstacles to overcome concerning power management, system integration, and the optimization of the synergy between the various components. Numerous forms of renewable energy, including wind and solar, are intermittent by nature. The intermittent nature of this power supply may result in variations in its accessibility, which may have an impact on the reliability and consistency of EET operations. It can be difficult to deal with the intermittent character of renewable energy sources with the continuous operation necessary for specific EET applications. An additional obstacle that may arise is the efficacy of transforming renewable energy into electrical power, which is subsequently employed in EET processes. It is essential to maximize overall efficiency by minimizing losses at each stage of the system and optimizing energy conversion processes. Unanticipated obstacles, such as fluctuations in microbial community dynamics or severe weather conditions, may affect the dependability and stability of the integrated system. Therefore, it is crucial that the stability and resilience of long-term practical implementation of integrated EET and renewable energy systems can be guaranteed.
- Leverage advanced molecular techniques for microbial community analysis and engineering: Advanced molecular techniques, including metagenomics, metatranscriptomics, and metaproteomics, offer the opportunity to thoroughly analyze the composition, functional capabilities, and activity of microbial communities in various ecological settings [141]. Through the implementation of these techniques, it becomes feasible to discern crucial microbial participants engaged in EET and attain a deeper understanding of their metabolic pathways, processes of electron transfer, and interconnections within intricate ecological systems. In addition, the utilization of sophisticated molecular techniques enables precise modification of microbial communities to enhance the efficiency of EET. This may encompass the engineering of crucial EET-related genes in dominant or electrochemically active microorganisms, including the creation of synthetic microbial communities with predetermined functions in electron transfer mechanisms. The utilization of molecular tools enables the development of customized strategies to address the adverse effects of microbial diversity on EET applications.
- Development of advanced electrode materials and reactor designs: It is possible to create electrodes that possess improved conductivity and surface characteristics that enable efficient long-distance electron transmission in larger-scale systems. Furthermore, the use of innovative reactor designs that enhance mass transport and promote biofilm development [142] can play a significant role in addressing the obstacles related to the scaling up of EET processes.
- Utilizing a wide range of substrates: Various microbes possess unique metabolic capacities. The provision of varied substrates will promote the proliferation of a heterogeneous microbial community, characterized by distinct enzymatic activities and pathways involved in EET. The adaptability of these entities enables them to adjust to varying circumstances and guarantees uninterrupted electron transport, even in situations when there is a scarcity of particular substrates. This phenomenon may also serve to bolster the stability and structure of biofilms, as a well-organized biofilm can improve the effectiveness of EET mechanisms by creating a conducive environment for microbial adhesion and electron transfer.
- Managing and controlling biofouling: The modification of surfaces involved in EET systems has the potential to mitigate the occurrence of biofouling. One such approach is the utilization of electrode materials possessing anti-fouling capabilities [139]. As biofouling is frequently driven by biofilm production, techniques aiming at stimulating the formation of conductive biofilms through the selection of electrode materials that enable the growth of conductive biofilms can be beneficial. Furthermore, efforts for controlling biofilm thickness and composition will aid in maintaining a balance between the advantages of biofilm-mediated electron transfer and the possible disadvantages of excessive biofilm growth.
- Introduction of energy storage and hybrid systems and smart grid technologies: Energy storage technologies, such as batteries and pumped hydro storage [140], can efficiently store surplus energy created during periods of high production and subsequently release it during periods of low output. By incorporating these storage devices with sustainable energy sources, the variations in power availability can be alleviated, hence offering a more uniform and reliable power provision for EET operations. Moreover, the integration of various renewable energy sources or the fusion of renewable energy with conventional power sources gives rise to hybrid systems. This strategy effectively broadens the range of energy sources utilized and reduces the impact of intermittent energy supply. Furthermore, the integration of smart grid technologies facilitates enhanced control and allocation of electrical power resources. These systems possess the potential to effectively manage the equilibrium between supply and demand by dynamically adjusting to variations in the availability of renewable energy sources.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Spietz, R.L.; Payne, D.; Szilagyi, R.; Boyd, E.S. Reductive biomining of pyrite by methanogens. Trends Microbiol. 2022, 30, 1072–1083. [Google Scholar] [CrossRef] [PubMed]
- Naaz, T.; Kumar, A.; Vempaty, A.; Singhal, N.; Pandit, S.; Gautam, P.; Jung, S.P. Recent advances in biological approaches towards anode biofilm engineering for improvement of extracellular electron transfer in microbial fuel cells. Environ. Eng. Res. 2023, 28, 220666. [Google Scholar] [CrossRef]
- Wang, Y.X.; Hou, N.; Liu, X.L.; Mu, Y. Advances in interfacial engineering for enhanced microbial extracellular electron transfer. Bioresour. Technol. 2022, 345, 126562. [Google Scholar] [CrossRef]
- Zou, L.; Huang, Y.-H.; Long, Z.-E.; Qiao, Y. On-going applications of Shewanella species in microbial electrochemical system for bioenergy, bioremediation and biosensing. World J. Microbiol. Biotechnol. 2019, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chiranjeevi, P.; Patil, S.A. Strategies for improving the electroactivity and specific metabolic functionality of microorganisms for various microbial electrochemical technologies. Biotechnol. Adv. 2020, 39, 107468. [Google Scholar] [CrossRef]
- Sibi, G. Environmental Biotechnology: Fundamentals to Modern Techniques; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Tucci, M.; Viggi, C.C.; Núnez, A.E.; Schievano, A.; Rabaey, K.; Aulenta, F. Empowering electroactive microorganisms for soil remediation: Challenges in the bioelectrochemical removal of petroleum hydrocarbons. Chem. Eng. J. 2021, 419, 130008. [Google Scholar] [CrossRef]
- Wang, R.; Li, H.; Sun, J.; Zhang, L.; Jiao, J.; Wang, Q.; Liu, S. Nanomaterials Facilitating Microbial Extracellular Electron Transfer at Interfaces. Adv. Mater. 2021, 33, 2004051. [Google Scholar] [CrossRef]
- Zhao, J.; Li, F.; Cao, Y.; Zhang, X.; Chen, T.; Song, H.; Wang, Z. Microbial extracellular electron transfer and strategies for engineering electroactive microorganisms. Biotechnol. Adv. 2020, 53, 107682. [Google Scholar] [CrossRef]
- Jiang, Y.; Zeng, R.J. Bidirectional extracellular electron transfers of electrode-biofilm: Mechanism and application. Bioresour. Technol. 2019, 271, 439–448. [Google Scholar] [CrossRef]
- Wen, L.; Huang, L.; Wang, Y.; Yuan, Y.; Zhou, L. Facet-engineered hematite boosts microbial electrogenesis by synergy of promoting electroactive biofilm formation and extracellular electron transfer. Sci. Total Environ. 2022, 819, 153154. [Google Scholar] [CrossRef]
- You, Z.; Li, J.; Wang, Y.; Wu, D.; Li, F.; Song, H. Advances in mechanisms and engineering of electroactive biofilms. Biotechnol. Adv. 2023, 66, 108170. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yu, H.; Zhang, J.; Qin, H. A short review of graphene in the microbial electrosynthesis of biochemicals from carbon dioxide. RSC Adv. 2022, 12, 22770–22782. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhang, J.; Zhang, Y.; Guo, Q.; Hu, T.; Xiao, H.; Lu, W.; Jia, J. Progress on anodic modification materials and future development directions in microbial fuel cells. J. Power Sources 2023, 556, 232486. [Google Scholar] [CrossRef]
- Tawalbeh, M.; Javed, R.M.N.; Al-Othman, A.; Almomani, F. The novel advancements of nanomaterials in biofuel cells with a focus on electrodes’ applications. Fuel 2022, 322, 124237. [Google Scholar] [CrossRef]
- Gemünde, A.; Lai, B.; Pause, L.; Krömer, J.; Holtmann, D. Redox mediators in microbial electrochemical systems. ChemElectroChem 2022, 9, e202200216. [Google Scholar] [CrossRef]
- Wang, X.; Chen, T.; Gao, C.; Xie, Y.; Zhang, A. Use of extracellular polymeric substances as natural redox mediators to enhance denitrification performance by accelerating electron transfer and carbon source metabolism. Bioresour. Technol. 2022, 345, 126522. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, J.A.; Bacame-Valenzuela, F.J.; Manríquez, J.; Castañeda-Zaldívar, F.; Reyes-Vidal, Y. Electrochemical analysis of extracellular electron transfer process of Pseudomonas aeruginosa NEJ07R using pyocyanin on a carbon electrode. J. Environ. Chem. Eng. 2023, 11, 110708. [Google Scholar] [CrossRef]
- Heijne, A.t.; Liu, D.; Sulonen, M.; Sleutels, T.; Fabregat-Santiago, F. Quantification of bio-anode capacitance in bioelectrochemical systems using Electrochemical Impedance Spectroscopy. J. Power Sources 2018, 400, 533–538. [Google Scholar] [CrossRef]
- Zhou, Y.; Takahashi, Y.; Fukuma, T.; Matsue, T. Scanning electrochemical microscopy for biosurface imaging. Curr. Opin. Electrochem. 2021, 29, 100739. [Google Scholar] [CrossRef]
- Fatima, S.; Aarti, T.; Sridhar, S. Overview of wastewater treatment approaches related to the microbial electrochemical system. In Advanced Nanomaterials and Nanocomposites for Bioelectrochemical Systems; Elsevier: Amsterdam, The Netherlands, 2023; pp. 57–80. [Google Scholar] [CrossRef]
- Zeppilli, M.; Cristiani, L.; Dell’Armi, E.; Villano, M. Potentiostatic vs galvanostatic operation of a Microbial Electrolysis Cell for ammonium recovery and biogas upgrading. Biochem. Eng. J. 2021, 167, 107886. [Google Scholar] [CrossRef]
- Doyle, L.; Marsili, E. The dynamics and characterization of electroactive biofilms. In Encyclopedia of Interfacial Chemistry: Surface Science and Electrochemistry; Elsevier BV: Amsterdam, The Netherlands, 2018; pp. 524–528. [Google Scholar]
- Matsena, M.T.; Nkhalambayausi Chirwa, E.M. Advances in microbial fuel cell technology for zero carbon emission energy generation from waste. In Biofuels and Bioenergy; Elsevier: Amsterdam, The Netherlands, 2022; pp. 321–358. [Google Scholar] [CrossRef]
- Zhou, J. Establishing Bisphenol a Degradation and Enhancing Microbial Fuel Cell Performance by Biofilm Optimization of Shewanella Oneidensis MR1. Ph.D. Thesis, Illinois Institute of Technology, Chicago, IL, USA, 2023. [Google Scholar]
- Hernandez, M.E.; Newman, D.K. Extracellular electron transfer. Cell. Mol. Life Sci. 2001, 58, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Kato, S. Biotechnological Aspects of Microbial Extracellular Electron Transfer. Microbes Environ. 2015, 30, 133–139. [Google Scholar] [CrossRef]
- Tavker, N.; Kumar, N. Bioelectrochemical systems: Understanding the basics and overcoming the challenges. In Development in Wastewater Treatment Research and Processes; Elsevier: Amsterdam, The Netherlands, 2023; pp. 79–98. [Google Scholar]
- Verma, M.; Singh, V.; Mishra, V. Moving towards the enhancement of extracellular electron transfer in electrogens. World J. Microbiol. Biotechnol. 2023, 39, 130. [Google Scholar] [CrossRef] [PubMed]
- Franza, T.; Gaudu, P. Quinones: More than electron shuttles. Res. Microbiol. 2022, 173, 103953. [Google Scholar] [CrossRef] [PubMed]
- Lusk, B.G.; Peraza, I.; Albal, G.; Marcus, A.K.; Popat, S.C.; Torres, C.I. pH Dependency in Anode Biofilms of Thermincola ferriacetica Suggests a Proton-Dependent Electrochemical Response. J. Am. Chem. Soc. 2018, 140, 5527–5534. [Google Scholar] [CrossRef] [PubMed]
- Kracke, F.; Vassilev, I.; Kromer, J.O. Microbial electron transport and energy conservation—The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6, 575. [Google Scholar] [CrossRef] [PubMed]
- Thapa, B.S.; Kim, T.; Pandit, S.; Song, Y.E.; Afsharian, Y.P.; Rahimnejad, M.; Kim, J.R.; Oh, S.E. Overview of electroactive microorganisms and electron transfer mechanisms in microbial electrochemistry. Bioresour. Technol. 2022, 347, 126579. [Google Scholar] [CrossRef]
- Levar, C.E.; Hoffman, C.L.; Dunshee, A.J.; Toner, B.M.; Bond, D.R. Redox potential as a master variable controlling pathways of metal reduction by Geobacter sulfurreducens. ISME J. 2017, 11, 741–752. [Google Scholar] [CrossRef]
- Liu, F.; Rotaru, A.-E.; Shrestha, P.M.; Malvankar, N.S.; Nevin, K.P.; Lovley, D.R. Promoting direct interspecies electron transfer with activated carbon. Energy Environ. Sci. 2012, 5, 8982–8989. [Google Scholar] [CrossRef]
- Rotaru, A.-E.; Shrestha, P.M.; Liu, F.; Markovaite, B.; Chen, S.; Nevin, K.P.; Lovley, D.R. Direct interspecies electron transfer between Geobacter metallireducens and Methanosarcina barkeri. Appl. Environ. Microbiol. 2014, 80, 4599–4605. [Google Scholar] [CrossRef]
- Zhang, K.; Deng, Y.; Liu, Z.; Feng, Y.; Hu, C.; Wang, Z. Biochar Facilitated Direct Interspecies Electron Transfer in Anaerobic Digestion to Alleviate Antibiotics Inhibition and Enhance Methanogenesis: A Review. Int. J. Environ. Res. Public Health 2023, 20, 2296. [Google Scholar] [CrossRef] [PubMed]
- Vellingiri, A.; Song, Y.E.; Munussami, G.; Kim, C.; Park, C.; Jeon, B.H.; Lee, S.G.; Kim, J.R. Overexpression of c-type cytochrome, CymA in Shewanella oneidensis MR-1 for enhanced bioelectricity generation and cell growth in a microbial fuel cell. J. Chem. Technol. Biotechnol. 2019, 94, 2115–2122. [Google Scholar] [CrossRef]
- Su, L.; Fukushima, T.; Prior, A.; Baruch, M.; Zajdel, T.J.; Ajo-Franklin, C.M. Modifying cytochrome c maturation can increase the bioelectronic performance of engineered Escherichia coli. ACS Synth. Biol. 2019, 9, 115–124. [Google Scholar] [CrossRef]
- Delgado, V.P.; Paquete, C.M.; Sturm, G.; Gescher, J. Improvement of the electron transfer rate in Shewanella oneidensis MR-1 using a tailored periplasmic protein composition. Bioelectrochemistry 2019, 129, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, Z.; Unver, T.; Zhang, B. CRISPR/Cas: A powerful tool for gene function study and crop improvement. J. Adv. Res. 2021, 29, 207–221. [Google Scholar] [CrossRef]
- Allen, R.M.; Bennetto, H.P. Microbial fuel-cells: Electricity production from carbohydrates. Appl. Biochem. Biotechnol. 1993, 39, 27–40. [Google Scholar] [CrossRef]
- Oz, M.; Lorke, D.E.; Hasan, M.; Petroianu, G.A. Cellular and molecular actions of methylene blue in the nervous system. Med. Res. Rev. 2011, 31, 93–117. [Google Scholar] [CrossRef]
- Arup, P. The reductase test for milk. Analyst 1918, 43, 20–31. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Zhang, Y.; Burns, K.; Zhao, N. Extracellular electron transfer in electroactive anaerobic granular sludge mediated by the phenothiazine derivative. J. Power Sources 2022, 527, 231212. [Google Scholar] [CrossRef]
- Babanova, S.; Hubenova, Y.; Mitov, M. Influence of artificial mediators on yeast-based fuel cell performance. J. Biosci. Bioeng. 2011, 112, 379–387. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, E.; Zhang, J.; Dai, Y.; Yang, Z.; Christensen, H.E.M.; Ulstrup, J.; Zhao, F. Extracellular polymeric substances are transient media for microbial extracellular electron transfer. Sci. Adv. 2017, 3, e1700623. [Google Scholar] [CrossRef] [PubMed]
- Song, R.B.; Wu, Y.; Lin, Z.Q.; Xie, J.; Tan, C.H.; Loo, J.S.C.; Cao, B.; Zhang, J.R.; Zhu, J.J.; Zhang, Q. Living and Conducting: Coating Individual Bacterial Cells with In Situ Formed Polypyrrole. Angew. Chem. Int. Ed. Engl. 2017, 56, 10516–10520. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. 2012, 109, 10042–10046. [Google Scholar] [CrossRef] [PubMed]
- Luckarift, H.R.; Sizemore, S.R.; Roy, J.; Lau, C.; Gupta, G.; Atanassov, P.; Johnson, G.R. Standardized microbial fuel cell anodes of silica-immobilized Shewanella oneidensis. Chem. Commun. 2010, 46, 6048–6050. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Fu, B.; Liu, F.; He, K.; Yang, H.; Ma, J.; Wang, H.; Zhang, X.; Liang, P.; Huang, X. Construction of innovative 3D-weaved carbon mesh anode network to boost electron transfer and microbial activity in bioelectrochemical system. Water Res. 2020, 172, 115493. [Google Scholar] [CrossRef]
- Zou, L.; Wu, X.; Huang, Y.; Ni, H.; Long, Z.E. Promoting Shewanella Bidirectional Extracellular Electron Transfer for Bioelectrocatalysis by Electropolymerized Riboflavin Interface on Carbon Electrode. Front. Microbiol. 2018, 9, 3293. [Google Scholar] [CrossRef]
- Bareia, T.; Pollak, S.; Eldar, A. Self-sensing in Bacillus subtilis quorum-sensing systems. Nat. Microbiol. 2018, 3, 83–89. [Google Scholar] [CrossRef]
- Reading, N.C.; Sperandio, V. Quorum sensing: The many languages of bacteria. FEMS Microbiol. Lett. 2006, 254, 1–11. [Google Scholar] [CrossRef]
- Hawver, L.A.; Jung, S.A.; Ng, W.-L. Specificity and complexity in bacterial quorum-sensing systems. FEMS Microbiol. Rev. 2016, 40, 738–752. [Google Scholar] [CrossRef]
- Venkataraman, A.; Rosenbaum, M.; Arends, J.B.; Halitschke, R.; Angenent, L.T. Quorum sensing regulates electric current generation of Pseudomonas aeruginosa PA14 in bioelectrochemical systems. Electrochem. Commun. 2010, 12, 459–462. [Google Scholar] [CrossRef]
- Yong, Y.-C.; Yu, Y.-Y.; Li, C.-M.; Zhong, J.-J.; Song, H. Bioelectricity enhancement via overexpression of quorum sensing system in Pseudomonas aeruginosa-inoculated microbial fuel cells. Biosens. Bioelectron. 2011, 30, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.B.; Chua, S.-L.; Cao, B.; Seviour, T.; Nesatyy, V.J.; Marsili, E.; Kjelleberg, S.; Givskov, M.; Tolker-Nielsen, T.; Song, H. Engineering PQS biosynthesis pathway for enhancement of bioelectricity production in Pseudomonas aeruginosa microbial fuel cells. PLoS ONE 2013, 8, e63129. [Google Scholar] [CrossRef] [PubMed]
- Monzon, O.; Yang, Y.; Li, Q.; Alvarez, P.J. Quorum sensing autoinducers enhance biofilm formation and power production in a hypersaline microbial fuel cell. Biochem. Eng. J. 2016, 109, 222–227. [Google Scholar] [CrossRef]
- Chen, S.; Jing, X.; Tang, J.; Fang, Y.; Zhou, S. Quorum sensing signals enhance the electrochemical activity and energy recovery of mixed-culture electroactive biofilms. Biosens. Bioelectron. 2017, 97, 369–376. [Google Scholar] [CrossRef]
- Li, F.; Wang, L.; Liu, C.; Wu, D.; Song, H. Engineering exoelectrogens by synthetic biology strategies. Curr. Opin. Electrochem. 2018, 10, 37–45. [Google Scholar] [CrossRef]
- Lovley, D.R.; Fraga, J.L.; Blunt-Harris, E.L.; Hayes, L.A.; Phillips, E.J.P.; Coates, J.D. Humic Substances as a Mediator for Microbially Catalyzed Metal Reduction. Acta Hydrochim. Hydrobiol. 1998, 26, 152–157. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E. Electromicrobiology: The ecophysiology of phylogenetically diverse electroactive microorganisms. Nat. Rev. Microbiol. 2022, 20, 5–19. [Google Scholar] [CrossRef]
- Xu, N.; Wang, T.L.; Li, W.J.; Wang, Y.; Chen, J.J.; Liu, J. Tuning Redox Potential of Anthraquinone-2-Sulfonate (AQS) by Chemical Modification to Facilitate Electron Transfer From Electrodes in Shewanella oneidensis. Front. Bioeng. Biotechnol. 2021, 9, 705414. [Google Scholar] [CrossRef]
- Yong, X.-Y.; Shi, D.-Y.; Chen, Y.-L.; Jiao, F.; Lin, X.; Zhou, J.; Wang, S.-Y.; Yong, Y.-C.; Sun, Y.-M.; OuYang, P.-K. Enhancement of bioelectricity generation by manipulation of the electron shuttles synthesis pathway in microbial fuel cells. Bioresour. Technol. 2014, 152, 220–224. [Google Scholar] [CrossRef]
- Feng, J.; Qian, Y.; Wang, Z.; Wang, X.; Xu, S.; Chen, K.; Ouyang, P. Enhancing the performance of Escherichia coli-inoculated microbial fuel cells by introduction of the phenazine-1-carboxylic acid pathway. J. Biotechnol. 2018, 275, 1–6. [Google Scholar] [CrossRef]
- Chen, Y.X.; Ing, N.L.; Wang, F.; Xu, D.; Sloan, N.B.; Lam, N.T.; Winter, D.L.; Egelman, E.H.; Hochbaum, A.I.; Clark, D.S. Structural determination of a filamentous chaperone to fabricate electronically conductive metalloprotein nanowires. ACS Nano 2020, 14, 6559–6569. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Tang, Q.; Li, Y.; Li, F.H.; Wu, J.H.; Li, W.W.; Yu, H.Q. Rapid and highly efficient genomic engineering with a novel iEditing device for programming versatile extracellular electron transfer of electroactive bacteria. Environ. Microbiol. 2021, 23, 1238–1255. [Google Scholar] [CrossRef] [PubMed]
- Ueki, T.; Walker, D.J.; Woodard, T.L.; Nevin, K.P.; Nonnenmann, S.S.; Lovley, D.R. An Escherichia coli chassis for production of electrically conductive protein nanowires. ACS Synth. Biol. 2020, 9, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, J.A.; Lopez, C.; Babanova, S.; Santoro, C.; Artyushkova, K.; Ista, L.; Schuler, A.J.; Atanassov, P. Surface modification for enhanced biofilm formation and electron transport in shewanella anodes. J. Electrochem. Soc. 2015, 162, H597. [Google Scholar] [CrossRef]
- Greenman, J.; Gajda, I.; You, J.; Mendis, B.A.; Obata, O.; Pasternak, G.; Ieropoulos, I. Microbial fuel cells and their electrified biofilms. Biofilm 2021, 3, 100057. [Google Scholar] [CrossRef]
- Zhao, C.E.; Gai, P.; Song, R.; Chen, Y.; Zhang, J.; Zhu, J.J. Nanostructured material-based biofuel cells: Recent advances and future prospects. Chem. Soc. Rev. 2017, 46, 1545–1564. [Google Scholar] [CrossRef]
- Jung, S.; Regan, J.M. Influence of external resistance on electrogenesis, methanogenesis, and anode prokaryotic communities in microbial fuel cells. Appl. Environ. Microbiol. 2011, 77, 564–571. [Google Scholar] [CrossRef]
- Caizan-Juanarena, L.; Borsje, C.; Sleutels, T.; Yntema, D.; Santoro, C.; Ieropoulos, I.; Soavi, F.; Ter Heijne, A. Combination of bioelectrochemical systems and electrochemical capacitors: Principles, analysis and opportunities. Biotechnol. Adv. 2020, 39, 107456. [Google Scholar] [CrossRef]
- Pasternak, G.; Greenman, J.; Ieropoulos, I. Dynamic evolution of anodic biofilm when maturing under different external resistive loads in microbial fuel cells. Electrochemical perspective. J. Power Sources 2018, 400, 392–401. [Google Scholar] [CrossRef]
- Cai, W.-F.; Geng, J.-F.; Pu, K.-B.; Ma, Q.; Jing, D.-W.; Wang, Y.-H.; Chen, Q.-Y.; Liu, H. Investigation of a two-dimensional model on microbial fuel cell with different biofilm porosities and external resistances. Chem. Eng. J. 2018, 333, 572–582. [Google Scholar] [CrossRef]
- Rossi, R.; Logan, B.E. Impact of external resistance acclimation on charge transfer and diffusion resistance in bench-scale microbial fuel cells. Bioresour. Technol. 2020, 318, 123921. [Google Scholar] [CrossRef]
- Koók, L.; Nemestóthy, N.; Bélafi-Bakó, K.; Bakonyi, P. The influential role of external electrical load in microbial fuel cells and related improvement strategies: A review. Bioelectrochemistry 2021, 140, 107749. [Google Scholar] [CrossRef]
- Dong, F.; Lee, Y.S.; Gaffney, E.M.; Liou, W.; Minteer, S.D. Engineering cyanobacterium with transmembrane electron transfer ability for bioelectrochemical nitrogen fixation. ACS Catal. 2021, 11, 13169–13179. [Google Scholar] [CrossRef]
- Sekar, N.; Jain, R.; Yan, Y.; Ramasamy, R.P. Enhanced photo-bioelectrochemical energy conversion by genetically engineered cyanobacteria. Biotechnol. Bioeng. 2016, 113, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Li, Y.; Jin, S.D.; Wang, X.; Wu, X.L.; Zeng, C.; Li, Y.; Ding, H.; Hao, R.-X.; Lv, M.; et al. Growth of non-phototrophic microorganisms using solar energy through mineral photocatalysis. Nat. Commun. 2012, 3, 768. [Google Scholar] [CrossRef]
- Cestellos-Blanco, S.; Zhang, H.; Kim, J.M.; Shen, Y.-X.; Yang, P. Photosynthetic semiconductor biohybrids for solar-driven biocatalysis. Nat. Catal. 2020, 3, 245–255. [Google Scholar] [CrossRef]
- Zhang, B.; Cheng, H.-Y.; Wang, A. Extracellular electron transfer through visible light induced excited-state outer membrane C-type cytochromes of Geobacter sulfurreducens. Bioelectrochemistry 2021, 138, 107683. [Google Scholar] [CrossRef]
- Ren, G.; Sun, Y.; Sun, M.; Li, Y.; Lu, A.; Ding, H. Visible light enhanced extracellular electron transfer between a hematite photoanode and Pseudomonas aeruginosa. Minerals 2017, 7, 230. [Google Scholar] [CrossRef]
- Ren, G.; Sun, Y.; Ding, Y.; Lu, A.; Li, Y.; Wang, C.; Ding, H. Enhancing extracellular electron transfer between Pseudomonas aeruginosa PAO1 and light driven semiconducting birnessite. Bioelectrochemistry 2018, 123, 233–240. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.; Lu, A.; Liu, Y.; Ren, G.; Li, Y.; Li, Y.; Ding, H. Extracellular electron transfer of electrochemically active bacteria community promoted by semiconducting minerals with photo-response in marine euphotic zone. Geomicrobiol. J. 2021, 38, 329–339. [Google Scholar] [CrossRef]
- Velasquez-Orta, S.B.; Head, I.M.; Curtis, T.P.; Scott, K.; Lloyd, J.R.; von Canstein, H. The effect of flavin electron shuttles in microbial fuel cells current production. Appl. Microbiol. Biotechnol. 2010, 85, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.H.; Yu, H.Q.; Li, W.W.; Wang, Y.K.; Sun, M.; Liu, X.W.; Sheng, G.P. Application of a weak magnetic field to improve microbial fuel cell performance. Ecotoxicology 2015, 24, 2175–2180. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ye, L.; Jin, J.; Chen, H.; Xu, X.; Zhu, L. Magnetite nanoparticles enhance the performance of a combined bioelectrode-UASB reactor for reductive transformation of 2,4-dichloronitrobenzene. Sci. Rep. 2017, 7, 10319. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayyahi, R.B.; Park, S.-G.; Jadhav, D.A.; Hussien, M.; Mohamed, H.O.; Castaño, P.; Al-Qaradawi, S.Y.; Chae, K.-J. Unraveling the influence of magnetic field on microbial and electrogenic activities in bioelectrochemical systems: A comprehensive review. Fuel 2023, 331, 125889. [Google Scholar] [CrossRef]
- Li, W.W.; Sheng, G.P.; Liu, X.W.; Cai, P.J.; Sun, M.; Xiao, X.; Wang, Y.K.; Tong, Z.H.; Dong, F.; Yu, H.Q. Impact of a static magnetic field on the electricity production of Shewanella-inoculated microbial fuel cells. Biosens. Bioelectron. 2011, 26, 3987–3992. [Google Scholar] [CrossRef]
- Yin, Y.; Huang, G.; Tong, Y.; Liu, Y.; Zhang, L. Electricity production and electrochemical impedance modeling of microbial fuel cells under static magnetic field. J. Power Sources 2013, 237, 58–63. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Ren, Y.; Wang, X. Effect of static magnetic field on the performances of and anode biofilms in microbial fuel cells. RSC Adv. 2016, 6, 82301–82308. [Google Scholar] [CrossRef]
- Zhou, H.; Mei, X.; Liu, B.; Xie, G.; Xing, D. Magnet anode enhances extracellular electron transfer and enrichment of exoelectrogenic bacteria in bioelectrochemical systems. Biotechnol. Biofuels 2019, 12, 1–9. [Google Scholar] [CrossRef]
- Tao, Q.; Zhou, S. Effect of static magnetic field on electricity production and wastewater treatment in microbial fuel cells. Appl. Microbiol. Biotechnol. 2014, 98, 9879–9887. [Google Scholar] [CrossRef]
- Chu, F.-J.; Sie, C.-Y.; Wan, T.-J.; Liu, S.-H.; Pai, T.-Y.; Kao, P.-M. Effects of magnetic fields on electricity generation in a photosynthetic ceramic microbial fuel cell. Int. J. Hydrogen Energy 2021, 46, 11411–11418. [Google Scholar] [CrossRef]
- Calignano, F.; Tommasi, T.; Manfredi, D.; Chiolerio, A. Additive manufacturing of a microbial fuel cell—A detailed study. Sci. Rep. 2015, 5, 17373. [Google Scholar] [CrossRef]
- Zawadzki, D.; Pędziwiatr, P.; Michalska, K. A novel microbial fuel cell with exchangable membrane-application of additive manufacturing technology for device fabrication. In Acta Innovations; Centrum Badań i Innowacji Pro-Akademia: Konstantynów Łódzki, Poland, 2018; pp. 20–31. [Google Scholar]
- Slate, A.J. Optimisation of a Pseudomonas aeruginosa Microbial Fuel Cell Coupled with Additive Manufacturing of Graphene Electrodes to Enhance Power Outputs. Ph.D. Thesis, Manchester Metropolitan University, Manchester, UK, 2019. [Google Scholar]
- Sawa, M.; Fantuzzi, A.; Bombelli, P.; Howe, C.J.; Hellgardt, K.; Nixon, P.J. Electricity generation from digitally printed cyanobacteria. Nat. Commun. 2017, 8, 1327. [Google Scholar] [CrossRef]
- Strack, G. Additive manufacturing approaches for biological power generation. Curr. Opin. Electrochem. 2019, 17, 167–173. [Google Scholar] [CrossRef]
- Choi, Y.; Jung, E.; Kim, S.; Jung, S. Membrane fluidity sensoring microbial fuel cell. Bioelectrochemistry 2003, 59, 121–127. [Google Scholar] [CrossRef]
- Picioreanu, C.; Katuri, K.P.; Van Loosdrecht, M.C.; Head, I.M.; Scott, K. Modelling microbial fuel cells with suspended cells and added electron transfer mediator. J. Appl. Electrochem. 2010, 40, 151–162. [Google Scholar] [CrossRef]
- Sousa, R., Jr.; Gonzalez, E.R. Mathematical modeling of polymer electrolyte fuel cells. J. Power Sources 2005, 147, 32–45. [Google Scholar] [CrossRef]
- Yao, K.; Karan, K.; McAuley, K.; Oosthuizen, P.; Peppley, B.; Xie, T. A review of mathematical models for hydrogen and direct methanol polymer electrolyte membrane fuel cells. Fuel Cells 2004, 4, 3–29. [Google Scholar] [CrossRef]
- de Ramón-Fernández, A.; Salar-García, M.J.; Fernández, D.R.; Greenman, J.; Ieropoulos, I. Evaluation of artificial neural network algorithms for predicting the effect of the urine flow rate on the power performance of microbial fuel cells. Energy 2020, 213, 118806. [Google Scholar] [CrossRef] [PubMed]
- Lesnik, K.L.; Liu, H. Predicting microbial fuel cell biofilm communities and bioreactor performance using artificial neural networks. Environ. Sci. Technol. 2017, 51, 10881–10892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.H.; Zhu, K.Y.; Zhuang, X.Y.; Liao, L.X.; Huang, S.Y.; Yao, C.Y.; Fang, B.S. A robust soft sensor to monitor 1, 3-propanediol fermentation process by Clostridium butyricum based on artificial neural network. Biotechnol. Bioeng. 2020, 117, 3345–3355. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Lesnik, K.L.; Wade, M.J.; Heidrich, E.S.; Wang, Y.; Liu, H. Incorporating microbial community data with machine learning techniques to predict feed substrates in microbial fuel cells. Biosens. Bioelectron. 2019, 133, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Guo, D.; Dang, Y.; Sun, D.; Li, P. Application of artificial intelligence-based methods in bioelectrochemical systems: Recent progress and future perspectives. J. Environ. Manag. 2023, 344, 118502. [Google Scholar] [CrossRef] [PubMed]
- Ieropoulos, I.; Melhuish, C.; Greenman, J.; Horsfield, I. EcoBot-II: An artificial agent with a natural metabolism. Int. J. Adv. Robot. Syst. 2005, 2, 31. [Google Scholar] [CrossRef]
- Dwivedi, K.A.; Huang, S.-J.; Wang, C.-T. Integration of various technology-based approaches for enhancing the performance of microbial fuel cell technology: A review. Chemosphere 2022, 287, 132248. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lu, Q.; Li, K.; Xu, S.; Wang, X.; Chen, K.; Ouyang, P. Construction of an Electron Transfer Mediator Pathway for Bioelectrosynthesis by Escherichia coli. Front. Bioeng. Biotechnol. 2020, 8, 590667. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.K.M.; Hussain, M.A.; Wahab, A.K.A. Fuzzy logic controller implementation on a microbial electrolysis cell for biohydrogen production and storage. Chin. J. Chem. Eng. 2021, 40, 149–159. [Google Scholar] [CrossRef]
- Wang, Y.; Song, X.; Cao, X.; Xu, Z.; Huang, W.; Wang, Y.; Ge, X. Integration of manganese ores with activated carbon granules into CW-MFC to trigger anoxic electron transfer and removal of ammonia nitrogen. J. Clean. Prod. 2022, 334, 130202. [Google Scholar] [CrossRef]
- Strik, D.P.; Hamelers, H.; Snel, J.F.; Buisman, C.J. Green electricity production with living plants and bacteria in a fuel cell. Int. J. Energy Res. 2008, 32, 870–876. [Google Scholar] [CrossRef]
- Schamphelaire, L.D.; Bossche, L.V.d.; Dang, H.S.; Höfte, M.; Boon, N.; Rabaey, K.; Verstraete, W. Microbial fuel cells generating electricity from rhizodeposits of rice plants. Environ. Sci. Technol. 2008, 42, 3053–3058. [Google Scholar] [CrossRef]
- Timmers, R.A.; Strik, D.P.; Hamelers, H.V.; Buisman, C.J. Long-term performance of a plant microbial fuel cell with Spartina anglica. Appl. Microbiol. Biotechnol. 2010, 86, 973–981. [Google Scholar] [CrossRef]
- Helder, M.; Strik, D.; Hamelers, H.; Kuhn, A.; Blok, C.; Buisman, C. Concurrent bio-electricity and biomass production in three Plant-Microbial Fuel Cells using Spartina anglica, Arundinella anomala and Arundo donax. Bioresour. Technol. 2010, 101, 3541–3547. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.V.; Mohanakrishna, G.; Chiranjeevi, P. Sustainable power generation from floating macrophytes based ecological microenvironment through embedded fuel cells along with simultaneous wastewater treatment. Bioresour. Technol. 2011, 102, 7036–7042. [Google Scholar] [CrossRef] [PubMed]
- Chiranjeevi, P.; Mohanakrishna, G.; Mohan, S.V. Rhizosphere mediated electrogenesis with the function of anode placement for harnessing bioenergy through CO2 sequestration. Bioresour. Technol. 2012, 124, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Xing, D.; Ren, Z.J. Microbial community structure accompanied with electricity production in a constructed wetland plant microbial fuel cell. Bioresour. Technol. 2015, 195, 115–121. [Google Scholar] [CrossRef]
- Klaisongkram, N.; Holasut, K. Electricity Generation of Plant Microbial Fuel Cell (PMFC) using Cyperus Involucratus R. Eng. Appl. Sci. Res. 2015, 42, 117–124. [Google Scholar]
- Wetser, K.; Sudirjo, E.; Buisman, C.J.; Strik, D.P. Electricity generation by a plant microbial fuel cell with an integrated oxygen reducing biocathode. Appl. Energy 2015, 137, 151–157. [Google Scholar] [CrossRef]
- Oon, Y.-L.; Ong, S.-A.; Ho, L.-N.; Wong, Y.-S.; Oon, Y.-S.; Lehl, H.K.; Thung, W.-E. Hybrid system up-flow constructed wetland integrated with microbial fuel cell for simultaneous wastewater treatment and electricity generation. Bioresour. Technol. 2015, 186, 270–275. [Google Scholar] [CrossRef]
- Ueoka, N.; Sese, N.; Sue, M.; Kouzuma, A.; Watanabe, K. Sizes of anode and cathode affect electricity generation in rice paddy-field microbial fuel cells. J. Sustain. Bioenergy Syst. 2016, 6, 10. [Google Scholar] [CrossRef]
- Li, J.; Han, H.; Chang, Y.; Wang, B. The material-microorganism interface in microbial hybrid electrocatalysis systems. Nanoscale 2023, 15, 6009–6024. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, C.; Li, Z.; Liu, J.; Xiao, X.; Li, D.; Chen, C.; Yu, M.; Feng, Y. Accelerating the extracellular electron transfer of Shewanella oneidensis MR-1 by carbon dots: The role of carbon dots concentration. Electrochim. Acta 2022, 421, 140490. [Google Scholar] [CrossRef]
- Liu, S.; Yi, X.; Wu, X.; Li, Q.; Wang, Y. Internalized Carbon Dots for Enhanced Extracellular Electron Transfer in the Dark and Light. Small 2020, 16, e2004194. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhou, X.; Qu, Y.; Liu, J.; Feng, Y.; Ren, N. Enhanced electricity generation and pollutant degradation by hybrid photoelectrochemical and microbial fuel cells. Energy Technol. 2017, 5, 402–405. [Google Scholar] [CrossRef]
- Tong, Y.; Wei, J.; Mo, R.; Ma, H.; Ai, F. Photocatalytic Microbial Fuel Cells and Performance Applications: A Review. Front. Chem. 2022, 10, 953434. [Google Scholar] [CrossRef] [PubMed]
- Tee, P.-F.; Abdullah, M.O.; Tan, I.A.W.; Amin, M.A.M.; Nolasco-Hipolito, C.; Bujang, K. Performance evaluation of a hybrid system for efficient palm oil mill effluent treatment via an air-cathode, tubular upflow microbial fuel cell coupled with a granular activated carbon adsorption. Bioresour. Technol. 2016, 216, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.T.; Baron, D.B.; Naranjo, B.; Bond, D.R.; Schmidt-Dannert, C.; Gralnick, J.A. Enhancement of survival and electricity production in an engineered bacterium by light-driven proton pumping. Appl. Environ. Microbiol. 2010, 76, 4123–4129. [Google Scholar] [CrossRef] [PubMed]
- Myers, B.; Hill, P.; Rawson, F.; Kovács, K. Enhancing Microbial Electron Transfer Through Synthetic Biology and Biohybrid Approaches: Part II: Combining approaches for clean energy. Johns. Matthey Technol. Rev. 2022, 66, 455–465. [Google Scholar] [CrossRef]
- Pankratova, G.; Hasan, K.; Leech, D.; Hederstedt, L.; Gorton, L. Electrochemical wiring of the Gram-positive bacterium Enterococcus faecalis with osmium redox polymer modified electrodes. Electrochem. Commun. 2017, 75, 56–59. [Google Scholar] [CrossRef]
- Widder, S.; Allen, R.J.; Pfeiffer, T.; Curtis, T.P.; Wiuf, C.; Sloan, W.T.; Cordero, O.X.; Brown, S.P.; Momeni, B.; Shou, W.; et al. Challenges in microbial ecology: Building predictive understanding of community function and dynamics. ISME J. 2016, 10, 2557–2568. [Google Scholar] [CrossRef]
- Patwardhan, S.B.; Savla, N.; Pandit, S.; Gupta, P.K.; Mathuriya, A.S.; Lahiri, D.; Jadhav, D.A.; Rai, A.K.; KanuPriya; Ray, R.R. Microbial fuel cell united with other existing technologies for enhanced power generation and efficient wastewater treatment. Appl. Sci. 2021, 11, 10777. [Google Scholar] [CrossRef]
- Vassilev, I.; Averesch, N.J.H.; Ledezma, P.; Kokko, M. Anodic electro-fermentation: Empowering anaerobic production processes via anodic respiration. Biotechnol. Adv. 2021, 48, 107728. [Google Scholar] [CrossRef]
- Pasternak, G.; de Rosset, A.; Tyszkiewicz, N.; Widera, B.; Greenman, J.; Ieropoulos, I. Prevention and removal of membrane and separator biofouling in bioelectrochemical systems: A comprehensive review. iScience 2022, 25, 104510. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Ajo-Franklin, C.M. Reaching full potential: Bioelectrochemical systems for storing renewable energy in chemical bonds. Curr. Opin. Biotechnol. 2019, 57, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Zhi, W.; Ge, Z.; He, Z.; Zhang, H. Methods for understanding microbial community structures and functions in microbial fuel cells: A review. Bioresour. Technol. 2014, 171, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Flimban, S.G.; Ismail, I.M.; Kim, T.; Oh, S.-E. Overview of recent advancements in the microbial fuel cell from fundamentals to applications: Design, major elements, and scalability. Energies 2019, 12, 3390. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactor Configuration | Plant | Maximum Power Generated | Ref. |
|---|---|---|---|
| Plant–MFC | Glyceria maxima | 67 mW/m2 | [116] |
| Plant–MFC | O. sativa | 330 mW/m2 | [117] |
| Plant–MFC | Spartina anglica | 100 mW/m2 | [118] |
| Plant–MFC | S. anglica | 222 mW/m2 | [119] |
| Plant–MFC | Arundinella anomala | 222 mW/m2 | [119] |
| Plant–MFC | Echinorriea crassipes | 224.93 mA/m2 | [120] |
| Plant–MFC | Pennisetum setaceum | 8.84 mW/m2 | [121] |
| Plant–MFC | Ipomoea aquatica | 105 mA/m2 | [122] |
| Plant–MFC | Cyperus involucratus | 5.99 mW/m2 | [123] |
| Plant–MFC | S. anglica | 240 mW/m2 | [124] |
| Plant–MFC | Typha latifolia | 6.12 mW/m2 | [125] |
| Plant–MFC | O. sativa | 140 mW/m2 | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazzan, O.O.; Zhao, B.; Xiao, Y. Strategies for Enhancing Extracellular Electron Transfer in Environmental Biotechnology: A Review. Appl. Sci. 2023, 13, 12760. https://doi.org/10.3390/app132312760
Hazzan OO, Zhao B, Xiao Y. Strategies for Enhancing Extracellular Electron Transfer in Environmental Biotechnology: A Review. Applied Sciences. 2023; 13(23):12760. https://doi.org/10.3390/app132312760
Chicago/Turabian StyleHazzan, Oluwadamilola Oluwatoyin, Biyi Zhao, and Yong Xiao. 2023. "Strategies for Enhancing Extracellular Electron Transfer in Environmental Biotechnology: A Review" Applied Sciences 13, no. 23: 12760. https://doi.org/10.3390/app132312760
APA StyleHazzan, O. O., Zhao, B., & Xiao, Y. (2023). Strategies for Enhancing Extracellular Electron Transfer in Environmental Biotechnology: A Review. Applied Sciences, 13(23), 12760. https://doi.org/10.3390/app132312760