2.1. Double Pulses in SPIDER
Previous publications addressing instability in SPIDER have included some analytical considerations of averaged SPIDER measurements [
7,
8], and we will use those calculations as a starting point for considering the specific case of double pulses. Given a spectral shear
and an internal pulse separation
in the SPIDER apparatus, a general expression for the ideal SPIDER trace of a single pulse as a function of the pulse complex electric field
is:
To understand how the pulse is retrieved from a SPIDER trace, this equation may be rewritten in terms of the spectral amplitude
and the derivative of the spectral phase (the group delay)
as:
The spectral fringes in the SPIDER trace depend on the group delay , and the spectral phase can be retrieved by integrating the measured group delay.
To understand the effect of multiple pulsing, we consider the SPIDER trace of a double pulse comprising two identical pulses with a temporal separation
a relative phase
. The SPIDER trace of this double pulse is then:
Expanding Equation (3) and collecting terms results in a lengthy, but tractable expression:
The first two terms above describe the SPIDER trace’s overall intensity envelope. This envelope contains spectral oscillations characteristic of a double pulse. The remaining four terms describe the SPIDER signal’s interference fringes. The first two oscillatory terms are typical fringes that would appear in common SPIDER measurements of each single pulse. As in all SPIDER measurements, each individual pulse interferes with a spectrally sheared replica of itself, creating fringes that are a function of the group delay. The second pair of fringe terms depend on both pulses in the double pulse. Here the first of the two pulses interferes with the sheared replica of the second pulse, and the second pulse interferes with the sheared replica of the first pulse. These fringe terms have different periodicities due to the different relative delays, and they also depend on the relative phase between the two pulses.
If
varies randomly from 0 to
, then in an averaged measurement some of the terms in Equation (4) would average to zero. Specifically, the spectral modulations in the envelope even out and the last two fringe terms disappear. In this case, the SPIDER-trace expression the simplifies to:
Notice that this expression differs only slightly from the general expression for SPIDER in Equation (2). In this expression there are two sets of fringes instead of just one. The two fringe terms are nearly identical and only differ by a factor of (the pulse separation times the spectral shear). To put the spectral shear in context, the temporal range in a SPIDER measurement is . is therefore times the ratio of the pulse separation and the SPIDER temporal range. The second fringe term has a phase offset from the first fringe term that is proportional to how large the pulse separation is compared to the temporal range of the SPIDER measurement. This ratio depends on both the pulse’s characteristics and the device parameters.
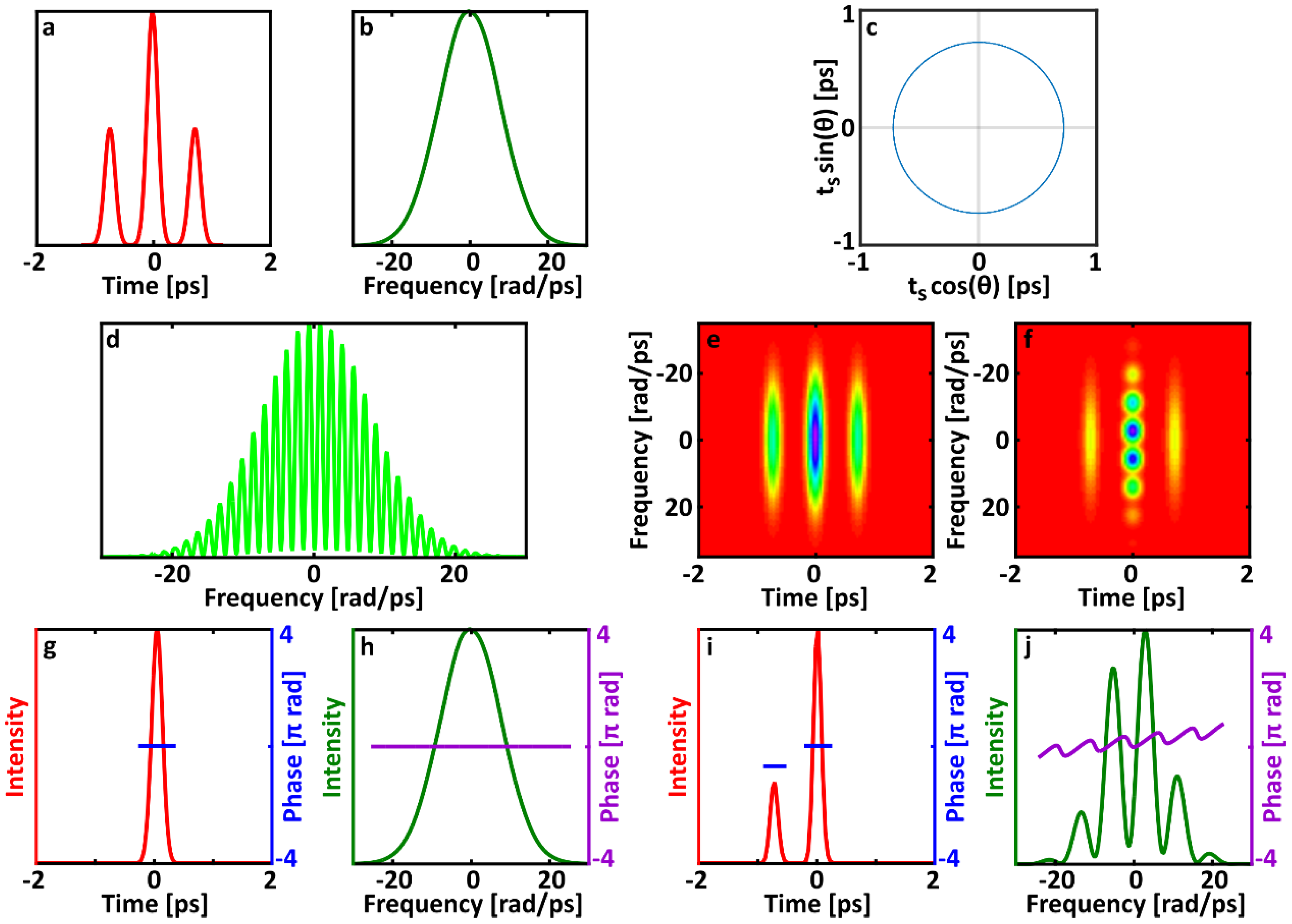
When the pulse separation is stable, there is a consistent phase offset between the fringes. Because the fringe terms are otherwise identical, their sum gives the same group delay as the individual terms. Thus, if the relative phase between the two pulses varies randomly, SPIDER will interpret double pulses as a single pulse with the temporal width of one of the individual pulses. In other words, ones of the pulses will be ignored completely. The size of the phase offset, which is controlled by the ratio of the pulse separation to the temporal support, determines how much background is present in the trace. If the separation between pulses is small compared to the temporal range (that is, support), then the sum of the two fringe terms cannot be distinguished from a single cosine and has almost no background. As the pulse separation increases, the fringe terms are less in phase and the background increases (see
Figure 2). Regardless of the amount of background, a SPIDER measurement of a double pulse with varying relative phase will always yield a single pulse. What is particularly interesting about this case is that it is possible to have very good fringe visibility and still have a very wrong measurement. In contrast with studies of other types of unstable pulses in SPIDER, having very low background is not a guarantee of measuring a pulse correctly.
If the pulse separation is not stable, then the phase offset between the two fringe terms is variable. There will be partial cancellation of fringes from the unstable pulse and some background present. As found previously, the fringes from the second pulse will vanish from the measurement if its relative arrival time varies by the temporal support.
In real SPIDER measurements, there is another issue that may affect how double pulses are measured. Because the spectral shear experienced by the pulse replicas is generated by interacting with different parts of an extremely chirped pulse, parts of the pulse occurring at different times will experience slightly different frequencies. While the frequency difference between the original pulse and the replica should remain constant in time, a double pulse will end up with slightly different frequencies for both pulses in the original pulse-shape and for both pulses in the replica. The modified expression for the SPIDER signal is:
where
is the difference in shear experienced by the two pulses in the double pulse. Compared to Equation (3), both the second pulse and its sheared replica experience an additional shear of
. This is not likely to have a large impact on the measurement, and this effect is not taken into account by the typical SPIDER retrieval procedure.
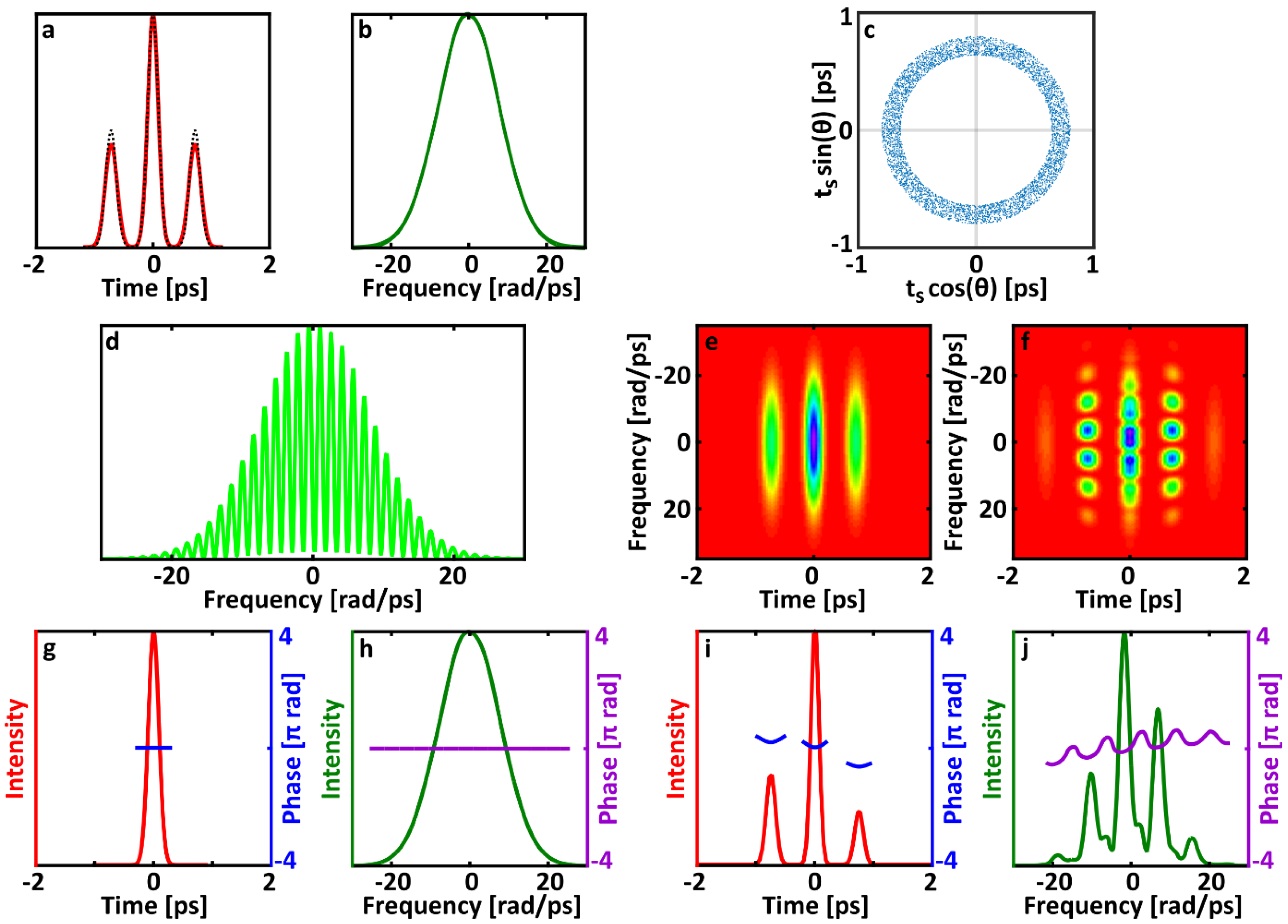
However, we have included the differences in shear for individual pulses in our simulations to obtain a more accurate approximation for the actual measurement process. Interestingly, comparing the averaged SPIDER traces in
Figure 3,
Figure 4 and
Figure 5 to previous simulations of double pulses in SPIDER [
18] shows that adding the differences in shear has essentially no impact on the SPIDER signal in this situation.
2.2. Double Pulses in FROG
General analytical consideration of FROG traces is usually impractical because expressions for self-referenced spectrograms tend to be quite complicated. Considering double pulses is simpler, however, and, assuming that the two pulses are both flat-phase Gaussians simplifies the calculation further. While these assumptions are restrictive, the resulting calculation is still quite informative for more general situations. Writing the time-domain electric field for a Gaussian double pulse:
the corresponding SHG FROG measurement is:
where
is again the delay between pulse replicas. After some calculations, this expression becomes:
The second and third terms inside the brackets are negligibly small because the pulse separation
is typically larger than
. The last (fourth) term can be written in a more intuitive form, resulting:
This expression describes the archetypal SHG FROG trace for double-pulses. The Gaussian frequency and delay amplitudes are equivalent to the FROG trace for a simple flat-phase Gaussian pulse. As is the case for all FROG traces of double-pulses, spectrogram has three intensity lobes, centered at and respectively. The middle lobe (the first term) has fringes that depend on the pulse separation as well as relative phase. The side lobes are half as intense as the center lobe but have the same temporal and spectral widths as does the center lobe.
Allowing the relative phase
to vary freely erases the fringes in the center lobe of the trace (see, for example,
Figure 3e). Significantly, the resulting trace, with three smooth (fringeless) lobes, which is the sum of many different traces corresponding to the many different double-pulse relative phases, is not a valid FROG trace for any pulse, no matter how complex. This is because the trace constrains both the temporal and spectral structure of the integrated electric field over many different pulses. Lobes separated in delay in a FROG trace clearly require the field to have satellite pulses, as in intensity autocorrelation. However, important for our purposes, stable satellite pulses also cause spectral fringes, and therefore the FROG trace should have frequency fringes in the center lobe. In fact, double pulses are used for calibration of the frequency axis in FROG for exactly this reason. Equivalently, the spectral structure of the FROG trace is constrained by the fact that the autoconvolution of the pulse spectrum must be equal to the frequency marginal (the sum of the trace over delay) [
11]. Integrating the smooth averaged trace over delay—computing the frequency marginal—yields a spectral profile that is smooth. Thus, the lack of spectral structure in the trace requires the spectrum to be smooth, but the temporal structure requires the pulse spectrum to be modulated. There is no electric field that satisfies both conditions. Consequently, the averaged FROG trace of double pulses with varying relative phase cannot correspond to any single pulse. This is a very useful and important feature of FROG—this discrepancy indicates the presence of instability. Indeed, such discrepancies were also found to occur in the presence of partial mode-locking in our previous work and so meant that FROG is a good indicator of that type of instability. This works because the FROG trace is two-dimensional, whereas the pulse is only one-dimensional, and the resulting over-determination of the pulse by the trace yields this additional information.
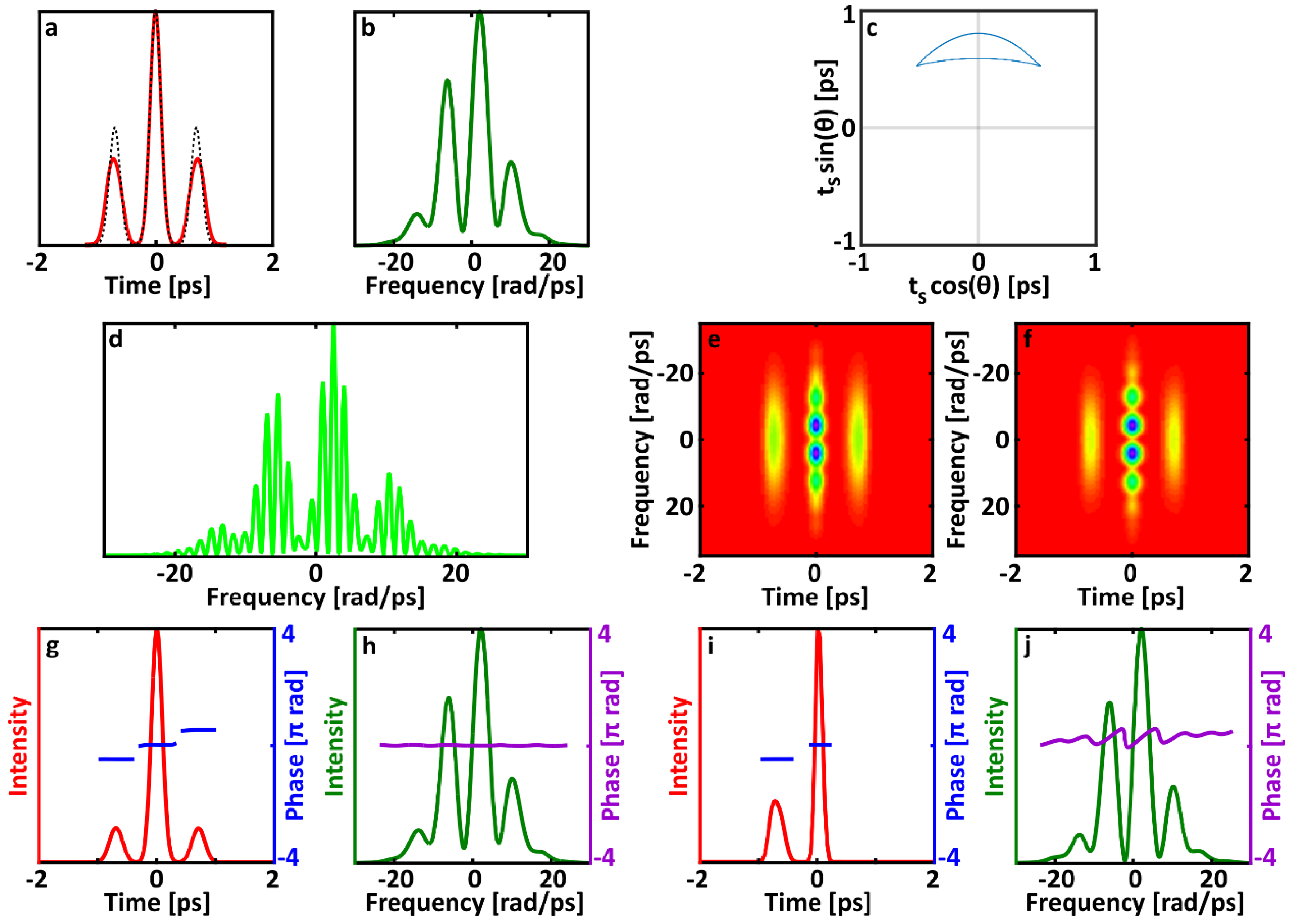
In the case of unstable double-pulsing, a variable pulse separation causes the position of the side lobes to vary. It also causes variations the periodicity of the fringes in the center lobe. Averaging over a range of pulse separations results in less intense, temporally wider side lobes in the averaged trace. Broadening the side lobes also causes the trace to become invalid for a single electric field. The spectrum of the side lobes is unchanged, but becoming longer in time with an equally broad spectrum requires consequently the addition of some phase variations, such as chirp. However, since the central lobe does not become wider in time, the field must also be nearly transform-limited. Variations in either pulse separation or relative phase lead to measured traces that are impossible to match with a single electric field. As a result, the FROG pulse-retrieval algorithm responds to these averaged spectrograms of unstable pulses cannot be predicted and is best studied using simulations.
On the other hand, the averaged SHG FROG trace still contains a significant amount of information about the ensemble of double pulses. In particular, the SHG FROG trace of a double pulse is easily recognized, and the lack of fringes in the central lobe is a clear indicator that there are variations in the relative phase of the second pulse. This allows a simple post-processing technique to find the relative intensity of the satellite pulse. Specifically, the relative energy of the two pulses can always be determined without ambiguity (even if the order in which they arrive is ambiguous) based on the energy contained in each lobe in the trace. Because the side lobes are a cross-correlation between the first and second pulses, their energy is proportional to the product of the energy of each individual pulse. The center lobe contains the autocorrelation of each pulse, and therefore its energy is proportional to the sum of the squares of the energy in each individual pulse. If one pulse has an energy of 1 and the other pulse has relative energy
, we can write expressions for the energy in the center lobe
and a side lobe
as:
The constants of proportionality are the same for each expression, and the exact energy of the pulses is often not of interest. Consequently, it makes sense to take the ratio of these two expressions. Multiplying through by
and rearranging gives a quadratic expression that is easily solved:
The energy in each lobe can be determined by integrating the corresponding sections of the measured FROG trace over frequency and delay. One the ratio of the energies has been found, then this quadratic expression can be solved to yield the relative energy, , of the second pulse.
Equation (13) holds true even when the relative phase and separation between the two pulses changes. Note that and refer to the total energy in each lobe, not merely their intensity. Thus, even though changing the separation of the two pulses in the double pulse will change the position of the side lobe, the energy will always be recorded in the trace. The total energy in the side lobes is unchanged, even though their peak intensity is lower when the pulse separation varies. This expression also holds true when the two pulses have different spectra or temporal profiles, although the relationship between the pulse energy and the pulse intensity will be different for each pulse in that case. If the relative energy of the two pulses varies, then the calculated relative energy will be the average relative energy. This very simple analysis can provide an additional check on FROG for double pulses. If the retrieved temporal pulse shape has pulses with a different relative energy from the one derived from the energy in the lobes of the trace, then there is further evidence of instability.
Our simulations will attempt to confirm the FROG post-processing approach presented above and study the FROG algorithm’s response to traces generated from unstable double pulses. With simulations, we can also investigate cases that are difficult to address analytically, such as the presence of small variations in the relative phase or of correlations between the pulse relative phase and separation.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





