An Automatic Analysis System for High-Throughput Clostridium Difficile Toxin Activity Screening


 ,
,  ,
,
Abstract
:1. Introduction
1.1. Clinical Definition and Motivation
1.2. Related Works
2. Methods
2.1. Automated Classification System
2.2. Image Preprocessing
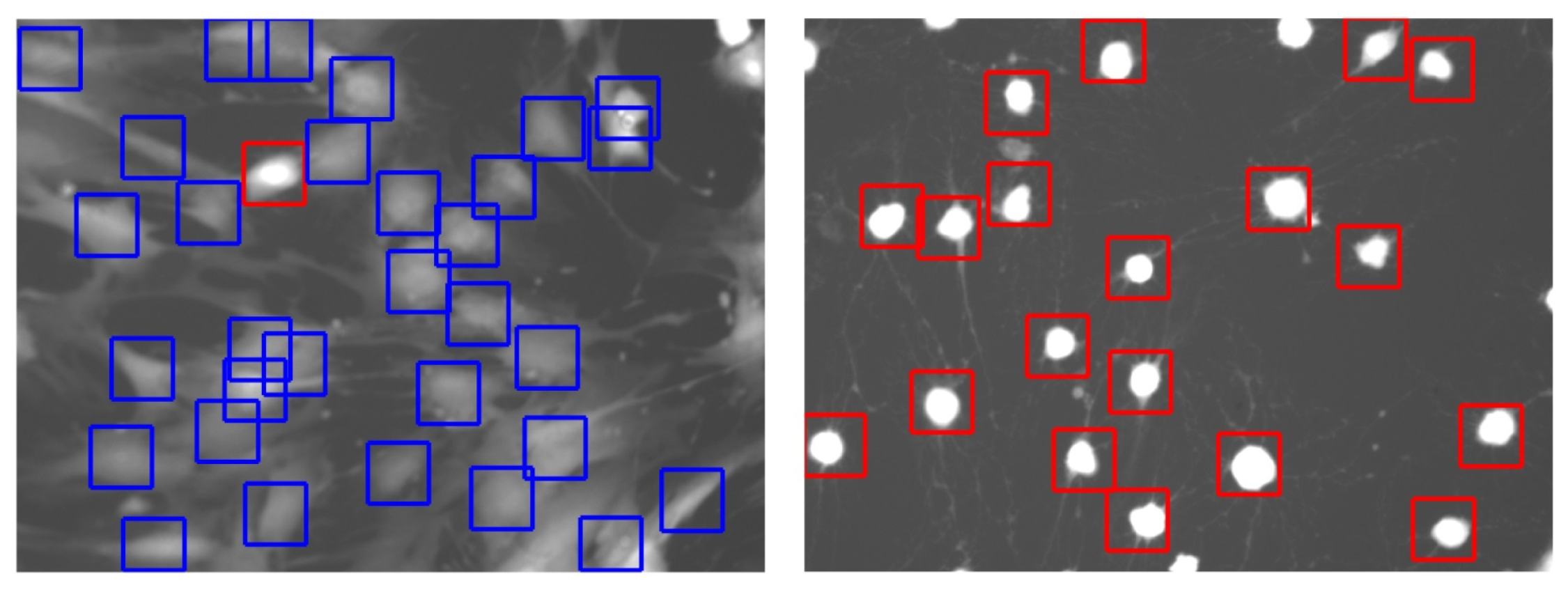
2.3. Cell Detection
2.4. Cell Segmentation
2.5. Classification
2.6. Cell-Rounding Assay
3. Results
3.1. Image Database
3.2. Classification Results
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carroll, K.C.; Bartlett, J.G. Biology of Clostridium difficile: Implications for epidemiology and diagnosis. Annu. Rev. Microbiol. 2011, 65, 501–521. [Google Scholar] [PubMed]
- Goudarzi, M.; Seyedjavadi, S.S.; Goudarzi, H.; Mehdizadeh Aghdam, E.; Nazeri, S. Clostridium difficile Infection: Epidemiology, Pathogenesis, Risk Factors, and Therapeutic Options. Scientifica 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J. Multistate point-prevalence survey of health care-associated infections. N. Engl. J. Med. 2014, 370, 1198–208. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.; Karlen, A. Discovery and preclinical development of new antibiotics. Ups. J. Med. Sci. 2014, 119, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, S. Meeting the societal need for new antibiotics: The challenges for the pharmaceutical industry. Br. J. Clin. Pharmacol. 2015, 79, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Lacy, D.B. Toward a structural understanding of Clostridium difficile toxins A and B. Front. Cell. Infect. Microbiol. 2012, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. QuickStats: Rates of Clostridium Difficile Infection among Hospitalized Patients Aged ≥65 Years,* by Age Group—National Hospital Discharge Survey, United States, 1996–2009. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6034a7.htm (accessed on 22 August 2018).
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium difficile colitis: pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, S.; Pardi, D.S. Clostridium difficile Infection: New Insights Into Management. Mayo Clin. Proc. 2012, 87, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.; Louie, T.J.; Gerding, D.N.; Cornely, O.A.; Chasan-Taber, S.; Fitts, D.; Gelone, S.P.; Broom, C.; Davidson, D.M. For the Polymer Alternative for CDI Treatment (PACT) Investigators. Vancomycin, metronidazole, or tolevamer for Clostridium difficile infection: Results from two multinational, randomized, controlled trials. Clin. Infect. Dis. 2014, 59, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P. Can we identify patients at high risk of recurrent Clostridium difficile infection? Clin. Microbiol. Infect. 2012, 18, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Garland, M.; Loscher, S.; Bogyo, M. Chemical Strategies To Target Bacterial Virulence. Chem. Rev. 2017, 117, 4422–4461. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.; Mooyottu, S.; Yin, H.; Surendran Nair, M.; Bhattaram, V.; Venkitanarayanan, K. Inhibiting Microbial Toxins Using Plant-Derived Compounds and Plant Extracts. Medicines 2015, 2, 186–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyne, L.; Warny, M.; Qamar, A.; Kelly, C.P. Asymptomatic Carriage of Clostridium difficile and Serum Levels of IgG Antibody against Toxin A. N. Engl. J. Med. 2000, 342, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.K.; Johnson, S.; Samore, M.H.; Bliss, D.Z.; Gerding, D.N. Primary symptomless colonisation by Clostridium difficile and decreased risk of subsequent diarrhoea. Lancet 1998, 351, 633–636. [Google Scholar] [CrossRef]
- Shen, A. Clostridium difficile toxins: Mediators of inflammation. J. Innate Immun. 2012, 4, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.O.; Garland, M.; Ferreyra, J.A.; Hryckowian, A.J.; Child, M.A.; Puri, A.W.; Solow-Cordero, D.E.; Higginbottom, S.K.; Segal, E.; Banaei, N.; et al. A small-molecule antivirulence agent for treating Clostridium difficile infection. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Lacy, D.B. Pre-Clinical Evaluation of Clostridium Difficile Toxin Inhibitors. Available online: http://grantome.com/grant/NIH/I01-BX002943-01 (accessed on 22 August 2018).
- Larabee, J.L.; Bland, S.J.; Hunt, J.J.; Ballard, J.D. Intrinsic toxin-derived peptides destabilize and inactivate Clostridium difficile TcdB. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Letourneau, J.J.; Stroke, I.L.; Hilbert, D.W.; Sturzenbecker, L.J.; Marinelli, B.A.; Quintero, J.G.; Sabalski, J.; Ma, L.; Diller, D.J.; Stein, P.D.; et al. Identification and initial optimization of inhibitors of Clostridium difficile (C. difficile) toxin B (TcdB). Bioorg. Med. Chem. Lett. 2018, 28, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Beilhartz, G.L.; Auger, A.; Gupta, P.; Therien, A.G.; Melnyk, R.A. Small Molecule Inhibitors of Clostridium difficile Toxin B-Induced Cellular Damage. Chem. Biol. 2015, 22, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Dordea, A.C.; Bray, M.-A.; Allen, K.; Logan, D.J.; Fei, F.; Malhotra, R.; Gregory, M.S.; Carpenter, A.E.; Buys, E.S. An open-source computational tool to automatically quantify immunolabeled retinal ganglion cells. Exp. Eye Res. 2016, 147, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumont, F. Manual Counting: Documentation. Available online: www.icy.bioimageanalysis.org/plugin/Manual_Counting. (accessed on 22 August 2018).
- Helmy, I.M.; Abdel Azim, A.M. Efficacy of ImageJ in the assessment of apoptosis. Diagn. Pathol. 2012, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocher, J.; Wagner, T. BioVoxxel Toolbox (ImageJ/Fiji). Available online: www.biovoxxel.de (accessed on 22 August 2018).
- Feng, Y.; Zhao, H.; Li, X.; Zhang, X.; Li, H. A multi-scale 3D Otsu thresholding algorithm for medical image segmentation. Digit. Signal Process. 2017, 60, 186–199. [Google Scholar] [CrossRef]
- Meyer, F. Topographic distance and watershed lines. Signal Process. 1994, 38, 113–125. [Google Scholar] [CrossRef]
- Zanaty, E.A.; Afifi, A. A watershed approach for improving medical image segmentation. Comput. Meth. Biomech. Biomed. Eng. 2013, 16, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.; Tibshirani, R.; Friedman, J.H. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer: Berlin, Germany, 2009. [Google Scholar]
- Japkowicz, N.; Shah, M. Evaluating Learning Algorithms: A Classification Perspective; Cambridge University Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Bishop, C.M. Pattern Recognition and Machine Learning; Springer: Berlin, Germany, 2006. [Google Scholar]
- Devroye, L.; Gyorfi, L.; Lugosi, G. A Probabilistic Theory of Pattern Recognition; Springer: Berlin, Germany, 1996. [Google Scholar]
- Duda, R.O.; Hart, P.E.; Stork, D.G. Pattern Classification, 2nd ed.; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Mitchell, T. Machine Learning; McGraw Hill: New York, NY, USA, 1997. [Google Scholar]
- Webb, A.R.; Copsey, K.D. Statistical Pattern Recognition, 3rd ed.; Wiley: New York, NY, USA, 2011. [Google Scholar]
- Al-Aidaroos, K.M.; Bakar, A.A.; Othman, Z. Medical data classification with Naïve Bayes approach. Inform. Technol. J. 2012, 11, 1166–1174. [Google Scholar] [CrossRef]
- John, G.H.; Langley, P. Estimating continuous distributions in bayesian classifiers. In Proceedings of the 11th Conference on Uncertainty in Artificial Intelligence, Montreal, QC, Canada, 18–20 August 1995. [Google Scholar]
- Khan, A.U.R.; Khan, M.; Khan, M.B. Naïve Multi-label Classification of YouTube Comments Using Comparative Opinion Mining. Procedia Comput. Sci. 2016, 82, 57–64. [Google Scholar] [CrossRef]
- Ontivero-Ortega, M.; Lage-Castellanos, A.; Valente, G.; Goebel, R.; Valdes-Sosa, M. Fast Gaussian Naïve Bayes for searchlight classification analysis. Neuroimage 2017, 163, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Sheikhian, H.; Delavar, M.R.; Stein, A. Predictive Modelling of Seismic Hazard Applying Naïve Bayes and Granular Computing Classifiers. Procedia Environ. Sci. 2015, 26, 49–52. [Google Scholar] [CrossRef]
- Vangelis, M.; Ion, A.; Geogios, P. Spam Filtering with Naïve Bayes—Which Naïve Bayes? In Proceedings of the 3rd Conference on Email and Anti-Spam, Mountain View, CA, USA, 27–28 July 2006.
- Hajian-Tilaki, K. Sample size estimation in diagnostic test studies of biomedical informatics. J. Biomed. Inform. 2014, 48, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, T. An Introduction to ROC Analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Feng, D.; Cortese, G.; Baumgartner, R. A comparison of confidence/credible interval methods for the area under the ROC curve for continuous diagnostic tests with small sample size. Stat. Methods Med. Res. 2015, 26, 2603–2621. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Chung, B.D.; Lee, J.S. Incorporating receiver operating characteristics into naive Bayes for unbalanced data classification. Computing 2017, 99, 203–218. [Google Scholar] [CrossRef]
- Airola, A.; Pahikkala, T.; Waegeman, W.; De Baets, B.; Salakoski, T. An experimental comparison of cross-validation techniques for estimating the area under the ROC curve. Comput. Stat. Data Anal. 2011, 55, 1828–1844. [Google Scholar] [CrossRef]
- Bamber, D. The area above the ordinal dominance graph and the area below the receiver operating characteristic graph. J. Math. Psychol. 1975, 12, 387–415. [Google Scholar] [CrossRef]
- Cortes, C.; Mohri, M. AUC optimization vs. error rate minimization. In Advances in Neural Information Processing Systems; MIT Press: Cambridge, MA, USA, 2004. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Software | Classification Algorithm | SN (%) | SP (%) | ACC (%) |
|---|---|---|---|---|
| ImageJ | Circularity | 67 | 71 | 69 |
| BioVoxxel Toolbox | Shape analysis | 82 | 84 | 82 |
| Described algorithm | Preprocessing, image enhancement, shape analysis, pixel brightness | 93 | 91 | 92.6 |
| Adherent (Predicted) | Rounded (Predicted) | Total | |
|---|---|---|---|
| Adherent (Actual) | 2552 () | 213 () | 2765 |
| Rounded (Actual) | 164 () | 2167 () | 2331 |
| Sensitivity: 93% | Specificity: 91% | Total: 5096 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garland, M.; Jaworek-Korjakowska, J.; Libal, U.; Bogyo, M.; Sieńczyk, M. An Automatic Analysis System for High-Throughput Clostridium Difficile Toxin Activity Screening. Appl. Sci. 2018, 8, 1512. https://doi.org/10.3390/app8091512
Garland M, Jaworek-Korjakowska J, Libal U, Bogyo M, Sieńczyk M. An Automatic Analysis System for High-Throughput Clostridium Difficile Toxin Activity Screening. Applied Sciences. 2018; 8(9):1512. https://doi.org/10.3390/app8091512
Chicago/Turabian StyleGarland, Megan, Joanna Jaworek-Korjakowska, Urszula Libal, Matthew Bogyo, and Marcin Sieńczyk. 2018. "An Automatic Analysis System for High-Throughput Clostridium Difficile Toxin Activity Screening" Applied Sciences 8, no. 9: 1512. https://doi.org/10.3390/app8091512
APA StyleGarland, M., Jaworek-Korjakowska, J., Libal, U., Bogyo, M., & Sieńczyk, M. (2018). An Automatic Analysis System for High-Throughput Clostridium Difficile Toxin Activity Screening. Applied Sciences, 8(9), 1512. https://doi.org/10.3390/app8091512