Electrical Stimulation through Conductive Substrate to Enhance Osteo-Differentiation of Human Dental Pulp-Derived Stem Cells


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture of Human Dental Pulp Stem Cells (hDPSCs) and Osteogenesis Induction
2.3. The Preparation of Polypyrrole (PPy) Films and the Fabrication of the Electrical Stimulation Device
2.4. Culture of hDPSCs on the Electrical Stimulation Devices and the Induction of Osteogenesis
2.5. Lactate Dehydrogenase (LDH) Assay
2.6. RNA Extraction and Real-Time Quantitative PCR (qPCR)
2.7. Alizarin Red S Staining
2.8. Quantitative and Qualitative Analyses of Calcium Deposition in the Extracellular Matrix (ECM)
2.9. Statistical Analysis
3. Results
3.1. Osteogenic Potential of Human Dental Pulp Stem Cells on Conductive PPy Films
3.2. Analysis of Gene Expression of the BMP Family and BMP Receptors in hDPSCs under Electrical Stimulation
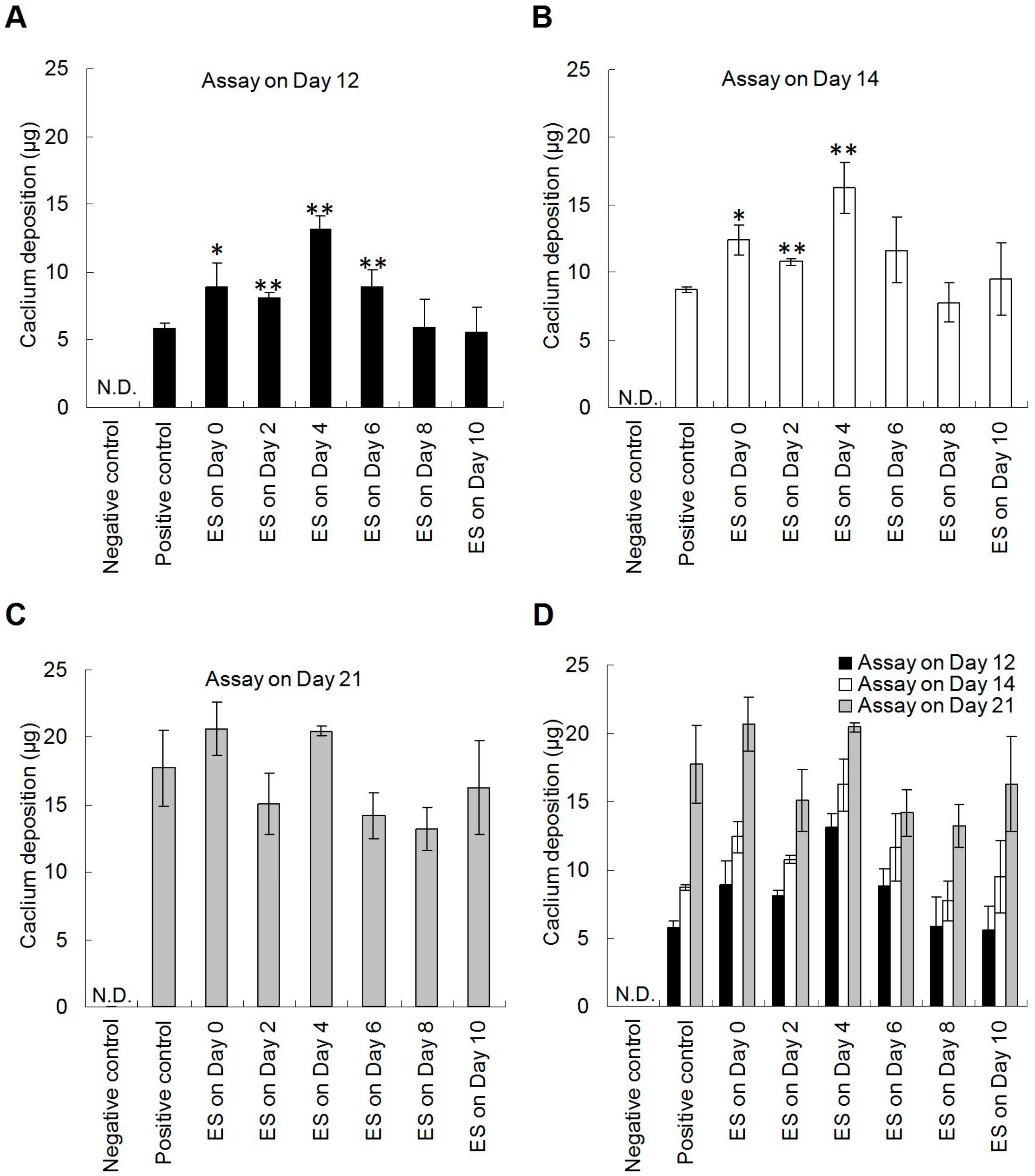
3.3. Electrical Stimulation Enhanced the Calcium Deposition of hDPSCs on PPy Films Under Osteogenesis Induction
3.4. Enhanced Potential Derived from ES in the Process of Osteogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grayson, W.L.; Bunnell, B.A.; Martin, E.; Frazier, T.; Hung, B.P.; Gimble, J.M. Stromal cells and stem cells in clinical bone regeneration. Nat. Rev. Endocrinol. 2015, 11, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Campanella, V. Dental stem cells: Current research and future applications. Eur. J. Paediatr. Dent. 2018, 19, 257. [Google Scholar] [PubMed]
- Ballini, A.; De Frenza, G.; Cantore, S.; Papa, F.; Grano, M.; Mastrangelo, F.; Tete, S.; Grassi, F.R. In vitro stem cell cultures from human dental pulp and periodontal ligament: New prospects in dentistry. Int. J. Immunopathol. Pharmacol. 2007, 20, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Garzon, I.; Martin-Piedra, M.A.; Alaminos, M. Human dental pulp stem cells. A promising epithelial-like cell source. Med. Hypotheses 2015, 84, 516–517. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (dpscs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Karaoz, E.; Demircan, P.C.; Saglam, O.; Aksoy, A.; Kaymaz, F.; Duruksu, G. Human dental pulp stem cells demonstrate better neural and epithelial stem cell properties than bone marrow-derived mesenchymal stem cells. Histochem. Cell Biol. 2011, 136, 455–473. [Google Scholar] [CrossRef]
- Li, J.H.; Liu, D.Y.; Zhang, F.M.; Wang, F.; Zhang, W.K.; Zhang, Z.T. Human dental pulp stem cell is a promising autologous seed cell for bone tissue engineering. Chin. Med. J. (Engl.) 2011, 124, 4022–4028. [Google Scholar]
- Hilkens, P.; Gervois, P.; Fanton, Y.; Vanormelingen, J.; Martens, W.; Struys, T.; Politis, C.; Lambrichts, I.; Bronckaers, A. Effect of isolation methodology on stem cell properties and multilineage differentiation potential of human dental pulp stem cells. Cell Tissue Res. 2013, 353, 65–78. [Google Scholar] [CrossRef]
- Iohara, K.; Zheng, L.; Wake, H.; Ito, M.; Nabekura, J.; Wakita, H.; Nakamura, H.; Into, T.; Matsushita, K.; Nakashima, M. A novel stem cell source for vasculogenesis in ischemia: Subfraction of side population cells from dental pulp. Stem Cells 2008, 26, 2408–2418. [Google Scholar] [CrossRef]
- Yang, R.; Chen, M.; Lee, C.H.; Yoon, R.; Lal, S.; Mao, J.J. Clones of ectopic stem cells in the regeneration of muscle defects in vivo. PLoS ONE 2010, 5, e13547. [Google Scholar] [CrossRef]
- Apel, C.; Forlenza, O.V.; de Paula, V.J.; Talib, L.L.; Denecke, B.; Eduardo, C.P.; Gattaz, W.F. The neuroprotective effect of dental pulp cells in models of alzheimer’s and parkinson’s disease. J. Neural Transm. (Vienna) 2009, 116, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Machado, E.; Fernandes, M.H.; Gomes Pde, S. Dental stem cells for craniofacial tissue engineering. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2012, 113, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.J.; Giannobile, W.V.; Helms, J.A.; Hollister, S.J.; Krebsbach, P.H.; Longaker, M.T.; Shi, S. Craniofacial tissue engineering by stem cells. J. Dent. Res. 2006, 85, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Heng, B.C.; Lo, E.C.; Zhang, C. Current advance and future prospects of tissue engineering approach to dentin/pulp regenerative therapy. Stem Cells Int. 2016, 2016, 9204574. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.L.; Stanford, C.M.; Keller, J.C. Calcium and phosphate supplementation promotes bone cell mineralization: Implications for hydroxyapatite (ha)-enhanced bone formation. J. Biomed. Mater. Res. 2000, 52, 270–278. [Google Scholar] [CrossRef]
- Cheng, C.C.; Chung, C.A.; Su, L.C.; Chien, C.C.; Cheng, Y.C. Osteogenic differentiation of placenta-derived multipotent cells in vitro. Taiwan J. Obstet. Gynecol. 2014, 53, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Reddi, A.H.; Reddi, A. Bone morphogenetic proteins (bmps): From morphogens to metabologens. Cytokine Growth Factor Rev. 2009, 20, 341–342. [Google Scholar] [CrossRef]
- Rogers, M.B.; Shah, T.A.; Shaikh, N.N. Turning bone morphogenetic protein 2 (bmp2) on and off in mesenchymal cells. J. Cell. Biochem. 2015, 116, 2127–2138. [Google Scholar] [CrossRef]
- Xie, H.; Cui, Z.; Wang, L.; Xia, Z.; Hu, Y.; Xian, L.; Li, C.; Xie, L.; Crane, J.; Wan, M.; et al. Pdgf-bb secreted by preosteoclasts induces angiogenesis during coupling with osteogenesis. Nat. Med. 2014, 20, 1270–1278. [Google Scholar] [CrossRef]
- McCaig, C.D.; Song, B.; Rajnicek, A.M. Electrical dimensions in cell science. J. Cell Sci. 2009, 122, 4267–4276. [Google Scholar] [CrossRef] [Green Version]
- Jeon, T.J.; Gao, R.; Kim, H.; Lee, A.; Jeon, P.; Devreotes, P.N.; Zhao, M. Cell migration directionality and speed are independently regulated by rasg and gbeta in dictyostelium cells in electrotaxis. Biol. Open 2019, 8, bio-042457. [Google Scholar] [CrossRef] [PubMed]
- Beugels, J.; Molin, D.G.M.; Ophelders, D.; Rutten, T.; Kessels, L.; Kloosterboer, N.; Grzymala, A.A.P.; Kramer, B.W.W.; van der Hulst, R.; Wolfs, T. Electrical stimulation promotes the angiogenic potential of adipose-derived stem cells. Sci. Rep. 2019, 9, 12076. [Google Scholar] [CrossRef] [PubMed]
- Peckham, P.H.; Knutson, J.S. Functional electrical stimulation for neuromuscular applications. Annu. Rev. Biomed. Eng. 2005, 7, 327–360. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Hei, W.H.; Kim, S.; Seo, Y.K.; Kim, S.M.; Jahng, J.W.; Lee, J.H. Co-treatment effect of pulsed electromagnetic field (pemf) with human dental pulp stromal cells and fk506 on the regeneration of crush injured rat sciatic nerve. Int. J. Neurosci. 2015, 125, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Im, A.-L.; Kim, J.; Lim, K.; Seonwoo, H.; Cho, W.; Choung, P.-H.; Chung, J.H. Effects of micro-electrical stimulation on regulation of behavior of electro-active stem cells. J. Biosyst. Eng. 2013, 38, 113–120. [Google Scholar] [CrossRef]
- Shamos, M.H.; Lavine, L.S.; Shamos, M.I. Piezoelectric effect in bone. Nature 1963, 197, 81. [Google Scholar] [CrossRef] [PubMed]
- Srirussamee, K.; Mobini, S.; Cassidy, N.J.; Cartmell, S.H. Direct electrical stimulation enhances osteogenesis by inducing bmp2 and spp1 expressions from macrophages and pre-osteoblasts. Biotechnol. Bioeng. 2019. [Google Scholar] [CrossRef]
- Hu, W.W.; Hsu, Y.T.; Cheng, Y.C.; Li, C.; Ruaan, R.C.; Chien, C.C.; Chung, C.A.; Tsao, C.W. Electrical stimulation to promote osteogenesis using conductive polypyrrole films. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 37, 28–36. [Google Scholar] [CrossRef]
- Shi, Z.; Gao, X.; Ullah, M.W.; Li, S.; Wang, Q.; Yang, G. Electroconductive natural polymer-based hydrogels. Biomaterials 2016, 111, 40–54. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, N.; Ma, M. Electroconductive hydrogels for biomedical applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, e1568. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Z. Polypyrrole as electrically conductive biomaterials: Synthesis, biofunctionalization, potential applications and challenges. Adv. Exp. Med. Biol. 2018, 1078, 347–370. [Google Scholar] [PubMed]
- Li, C.; Hsu, Y.-T.; Hu, W.-W. The regulation of osteogenesis using electroactive polypyrrole films. Polymers 2016, 8, 258. [Google Scholar] [CrossRef]
- Hu, W.W.; Chen, T.C.; Tsao, C.W.; Cheng, Y.C. The effects of substrate-mediated electrical stimulation on the promotion of osteogenic differentiation and its optimization. J. Biomed. Mater. Res. B Appl. Biomater. 2019, 107, 1607–1619. [Google Scholar] [CrossRef]
- Dawson, E.R.; Suzuki, R.K.; Samano, M.A.; Murphy, M.B. Increased internal porosity and surface area of hydroxyapatite accelerates healing and compensates for low bone marrow mesenchymal stem cell concentrations in critically-sized bone defects. Appl. Sci. 2018, 8, 366. [Google Scholar] [CrossRef]
- Kugimiya, F.; Ohba, S.; Nakamura, K.; Kawaguchi, H.; Chung, U.I. Physiological role of bone morphogenetic proteins in osteogenesis. J. Bone Miner. Metab. 2006, 24, 95–99. [Google Scholar] [CrossRef]
- Struys, T.; Moreels, M.; Martens, W.; Donders, R.; Wolfs, E.; Lambrichts, I. Ultrastructural and immunocytochemical analysis of multilineage differentiated human dental pulp- and umbilical cord-derived mesenchymal stem cells. Cells Tissues Organs 2011, 193, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Potdar, P.D.; Jethmalani, Y.D. Human dental pulp stem cells: Applications in future regenerative medicine. World J. Stem Cells 2015, 7, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gilmore, K.J.; Moulton, S.E.; Wallace, G.G. Electrical stimulation promotes nerve cell differentiation on polypyrrole/poly (2-methoxy-5 aniline sulfonic acid) composites. J. Neural Eng. 2009, 6, 065002. [Google Scholar] [CrossRef]
- Mobini, S.; Leppik, L.; Thottakkattumana Parameswaran, V.; Barker, J.H. In vitro effect of direct current electrical stimulation on rat mesenchymal stem cells. PeerJ 2017, 5, e2821. [Google Scholar] [CrossRef]
- Fakhry, M.; Hamade, E.; Badran, B.; Buchet, R.; Magne, D. Molecular mechanisms of mesenchymal stem cell differentiation towards osteoblasts. World J. Stem Cells 2013, 5, 136–148. [Google Scholar] [CrossRef]
- Terhi, J.H.; Teuvo, A.H. Differentiation of osteoblasts and osteocytes from mesenchymal stem cells. Curr. Stem Cell Res. Ther. 2008, 3, 131–145. [Google Scholar]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone morphogenetic protein (bmp) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Alves, G.G.; Zambuzzi, W.F.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Structure, biological function and therapeutic applications. Arch. Biochem. Biophys. 2014, 561, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. Tgf-β/bmp signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.; Greenspan, D.S. Developmental roles of the bmp1/tld metalloproteinases. Birth Defects Res. C Embryo Today 2006, 78, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Daluiski, A.; Engstrand, T.; Bahamonde, M.E.; Gamer, L.W.; Agius, E.; Stevenson, S.L.; Cox, K.; Rosen, V.; Lyons, K.M. Bone morphogenetic protein-3 is a negative regulator of bone density. Nat. Genet. 2001, 27, 84. [Google Scholar] [CrossRef] [PubMed]
- Gamer, L.W.; Ho, V.; Cox, K.; Rosen, V. Expression and function of bmp3 during chick limb development. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2008, 237, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Dudley, A.T.; Lyons, K.M.; Robertson, E.J. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef]
- Nguyen, V.; Meyers, C.A.; Yan, N.; Agarwal, S.; Levi, B.; James, A.W. Bmp-2-induced bone formation and neural inflammation. J. Orthop. 2017, 14, 252–256. [Google Scholar] [CrossRef]
- Shahrul Hisham Zainal, A.; Thanaletchumi, M.; Intan Zarina Zainol, A.; Rohaya Megat Abdul, W.; Sahidan, S. A perspective on stem cells as biological systems that produce differentiated osteoblasts and odontoblasts. Curr. Stem Cell Res. Ther. 2017, 12, 247–259. [Google Scholar]
- Yadin, D.; Knaus, P.; Mueller, T.D. Structural insights into bmp receptors: Specificity, activation and inhibition. Cytokine Growth Factor Rev. 2016, 27, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Hang, A.; Phan, E.; Wildsoet, C.F. Differential gene expression of bmp2 and bmp receptors in chick retina & choroid induced by imposed optical defocus. Vis. Neurosci. 2016, 33, E015. [Google Scholar] [PubMed]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. Camp/pka pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [PubMed]
- Zayzafoon, M. Calcium/calmodulin signaling controls osteoblast growth and differentiation. J. Cell. Biochem. 2006, 97, 56–70. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers |
|---|---|
| GAPDH | 5′-CTCTGCTCCTCCTGTTCGAC |
| 3′-ACGACCAAATCCGTTGACTC | |
| BMP1 | 5′-ACCCTGGGCAGCTACAAGT |
| 3′-TGAGGAATCCGCCACAAG | |
| BMP2 | 5′-CAGACCACCGGTTGGAGA |
| 3′-CCCACTCGTTTCTGGTAGTTCT | |
| BMP3 | 5′-CCCAAGTCCTTTGATGCCTA |
| 3′-TCTGGATGGTAGCATGATTTGA | |
| BMP4 | 5′-CTTTACCGGCTTCAGTCTGG |
| 3′-TGGGATGTTCTCCAGATGTTC | |
| BMP5 | 5′-AACCGCAATAAATCCAGCTC |
| 3′-TTTTGCTCACTTGTGTTATAATCTCC | |
| BMP6 | 5′-ACATGGTCATGAGCTTTGTGA |
| 3′-ACTCTTTGTGGTGTCGCTGA | |
| BMP7 | 5′-ACCACTGGGTGGTCAATCC |
| 3′-CAACTTGGGGTTGATGCTCT | |
| BMPR1A | 5′-GGACGAAAGCCTGAACAAAA |
| 3′-GCAATTGGTATTCTTCCACGA | |
| BMPR1B | 5′-CGAATGTAATAAAGACCTACACCCTA |
| 3′-GTGTATAGGTCCATCAACAAAATCTC | |
| BMPR2 | 5′-TCTGGATCTTTCAGCCACAA |
| 3′-TGCCATCTTGTGTTGACTCAC |
| Solutions for PPy Preparation | Sheet Resistances of PPy Films (kΩ/Square) |
|---|---|
| 0.1 M pyrrole | 25.72 ± 1.52 |
| 0.3 M pyrrole | 10.58 ± 0.65 |
| 0.5 M pyrrole | 7.83 ± 0.47 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.-C.; Chen, C.-H.; Kuo, H.-W.; Yen, T.-L.; Mao, Y.-Y.; Hu, W.-W. Electrical Stimulation through Conductive Substrate to Enhance Osteo-Differentiation of Human Dental Pulp-Derived Stem Cells. Appl. Sci. 2019, 9, 3938. https://doi.org/10.3390/app9183938
Cheng Y-C, Chen C-H, Kuo H-W, Yen T-L, Mao Y-Y, Hu W-W. Electrical Stimulation through Conductive Substrate to Enhance Osteo-Differentiation of Human Dental Pulp-Derived Stem Cells. Applied Sciences. 2019; 9(18):3938. https://doi.org/10.3390/app9183938
Chicago/Turabian StyleCheng, Yu-Che, Chien-Hsun Chen, Hong-Wei Kuo, Ting-Ling Yen, Ya-Yuan Mao, and Wei-Wen Hu. 2019. "Electrical Stimulation through Conductive Substrate to Enhance Osteo-Differentiation of Human Dental Pulp-Derived Stem Cells" Applied Sciences 9, no. 18: 3938. https://doi.org/10.3390/app9183938
APA StyleCheng, Y. -C., Chen, C. -H., Kuo, H. -W., Yen, T. -L., Mao, Y. -Y., & Hu, W. -W. (2019). Electrical Stimulation through Conductive Substrate to Enhance Osteo-Differentiation of Human Dental Pulp-Derived Stem Cells. Applied Sciences, 9(18), 3938. https://doi.org/10.3390/app9183938