Molecular Characterization of an Endo-β-1,4-Glucanase, CelAJ93, from the Recently Isolated Marine Bacterium, Cellulophaga sp. J9-3


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation of Microorganisms
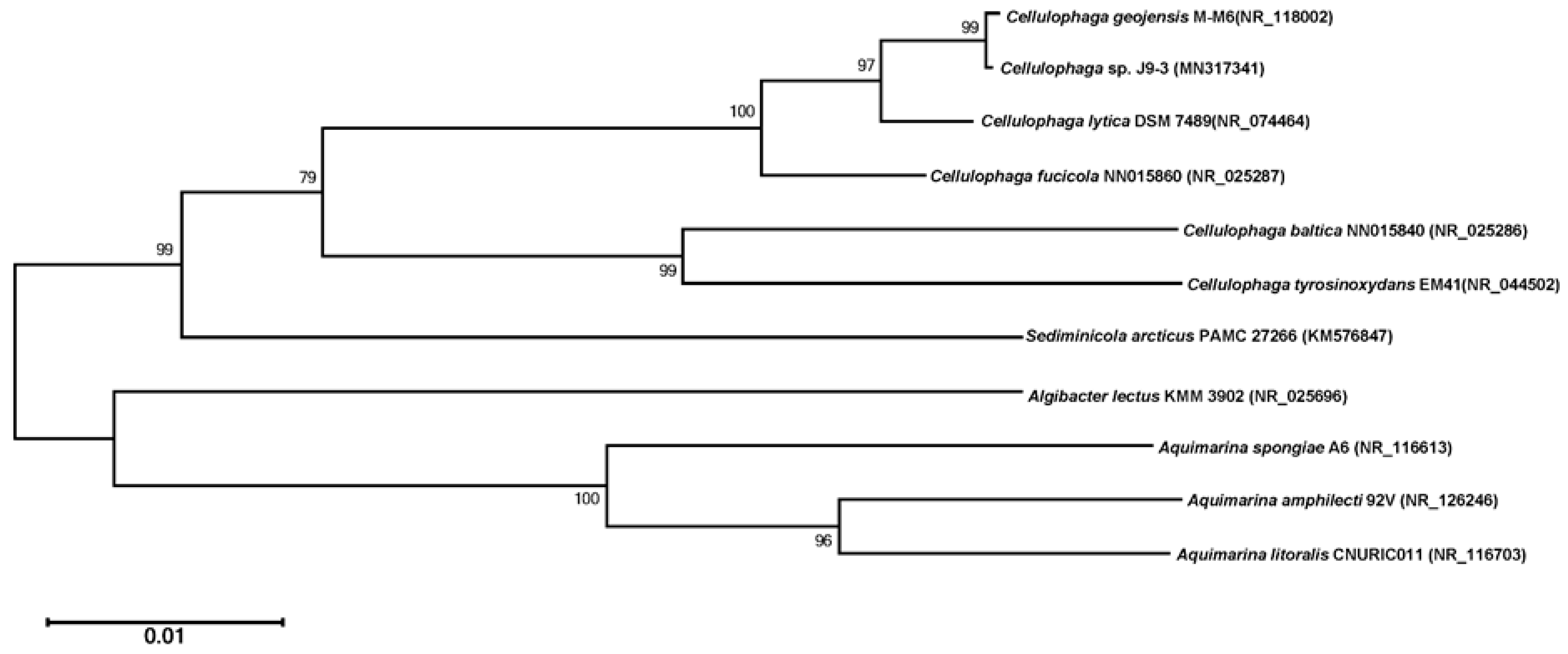
2.3. Phylogenetic Analysis through 16S rRNA Gene and Whole-Genome Sequencing
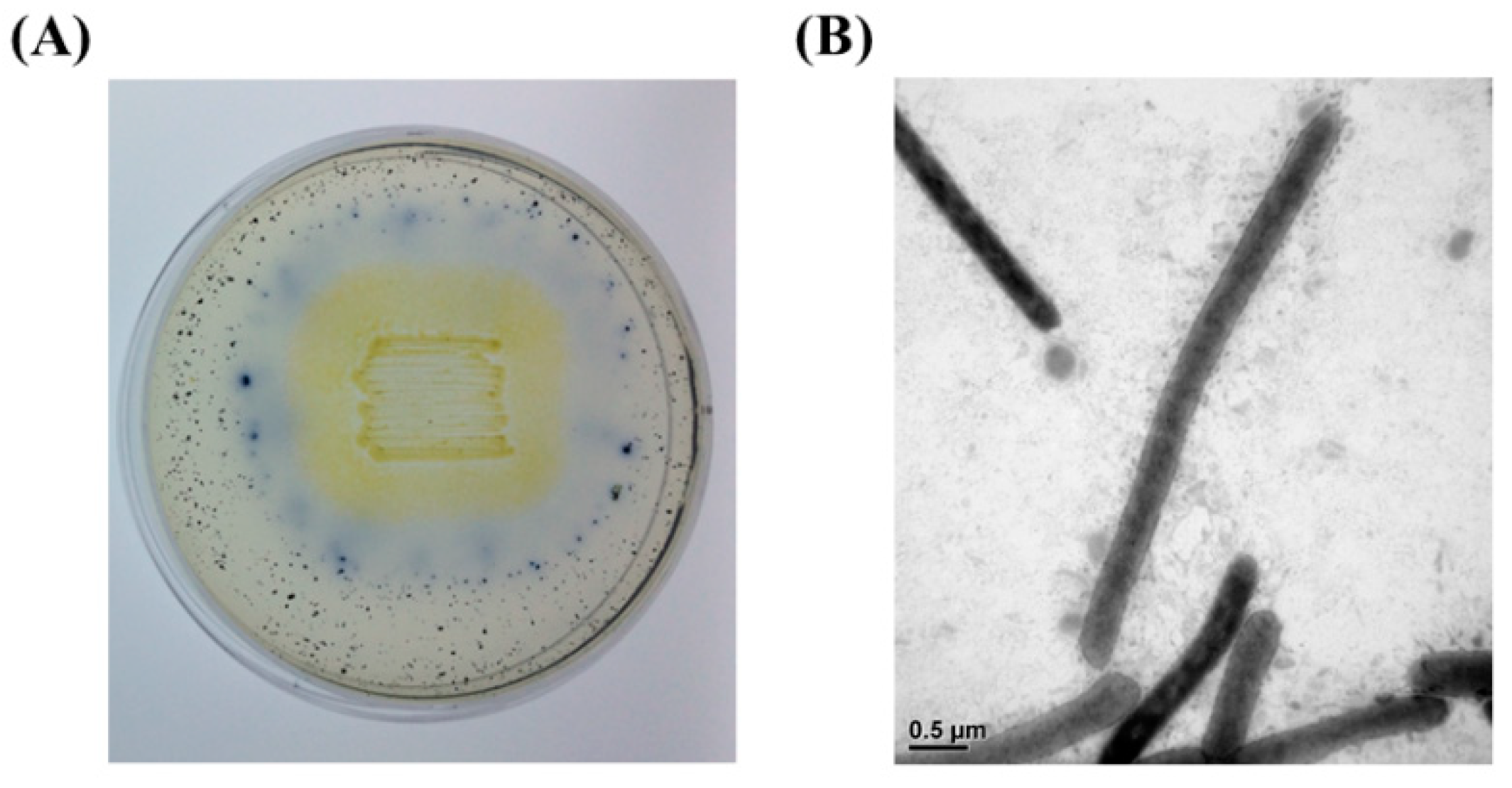
2.4. Morphological Characterization
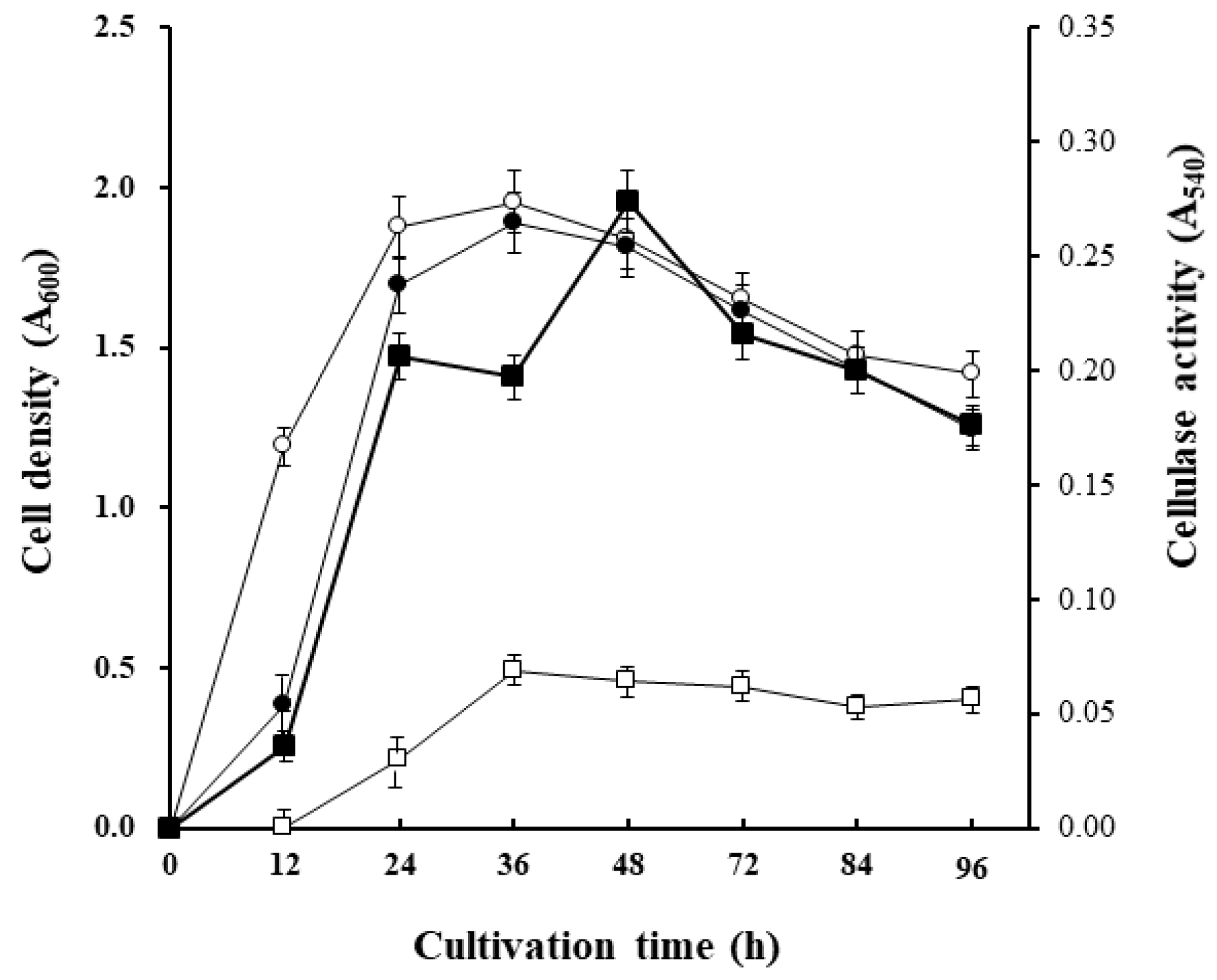
2.5. Cellulase Production by the J9-3 Strain in Liquid Culture
2.6. Measurement of Cellulase Activity by the DNS Method
2.7. Purification of Cellulase
2.8. Biochemical Characterization of Cellulase
2.9. Thin Layer Chromatography (TLC) Analysis of Enzyme Reaction
3. Results
3.1. Identification of Strain J9-3
3.2. Cellulase Production by Cellulophaga sp. J9-3 in Liquid Culture
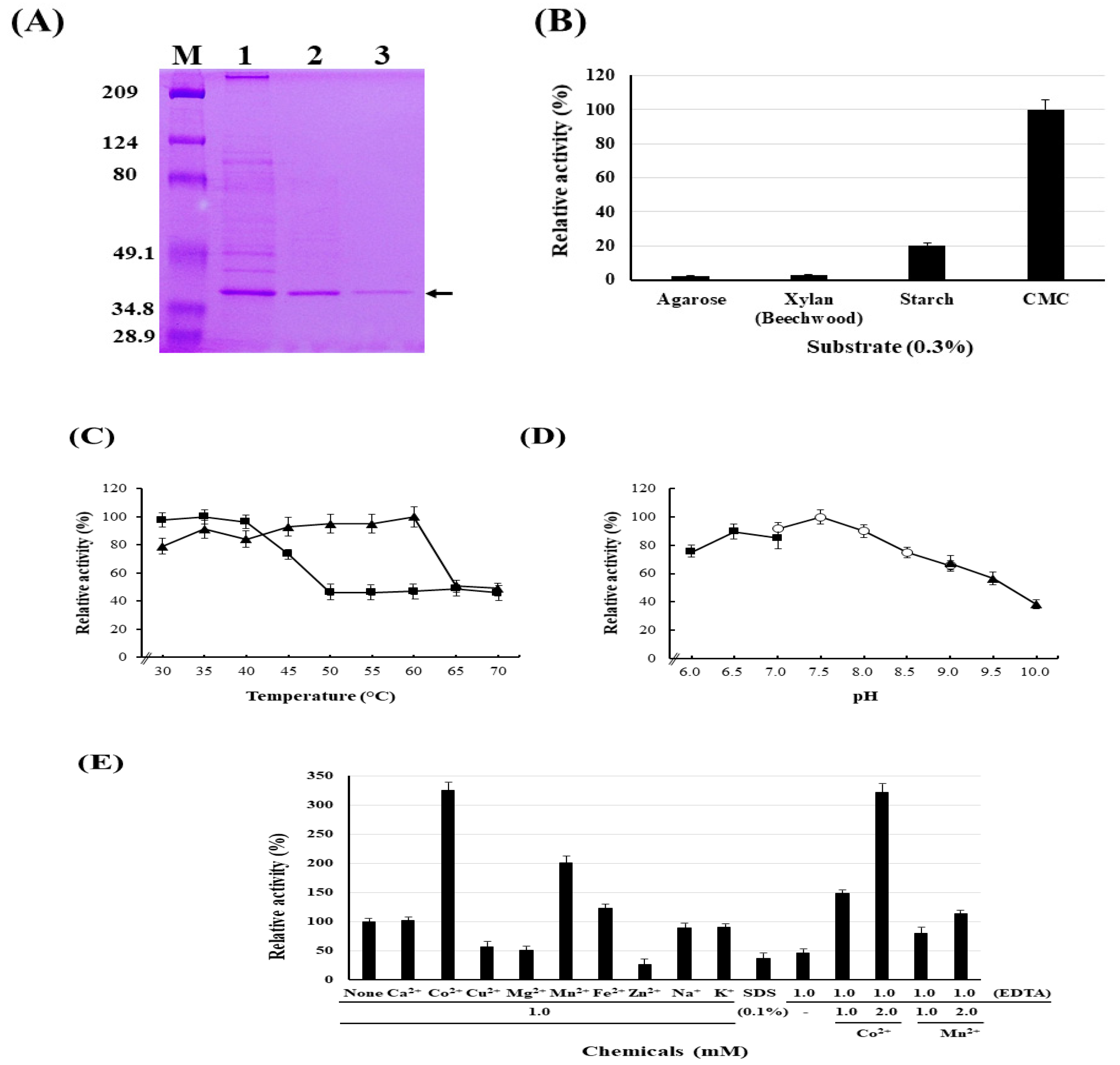
3.3. Purification of Cellulase
3.4. Identification of Cellulase Enzymes and their Genes
3.5. Biochemical Characterization of the Cellulase
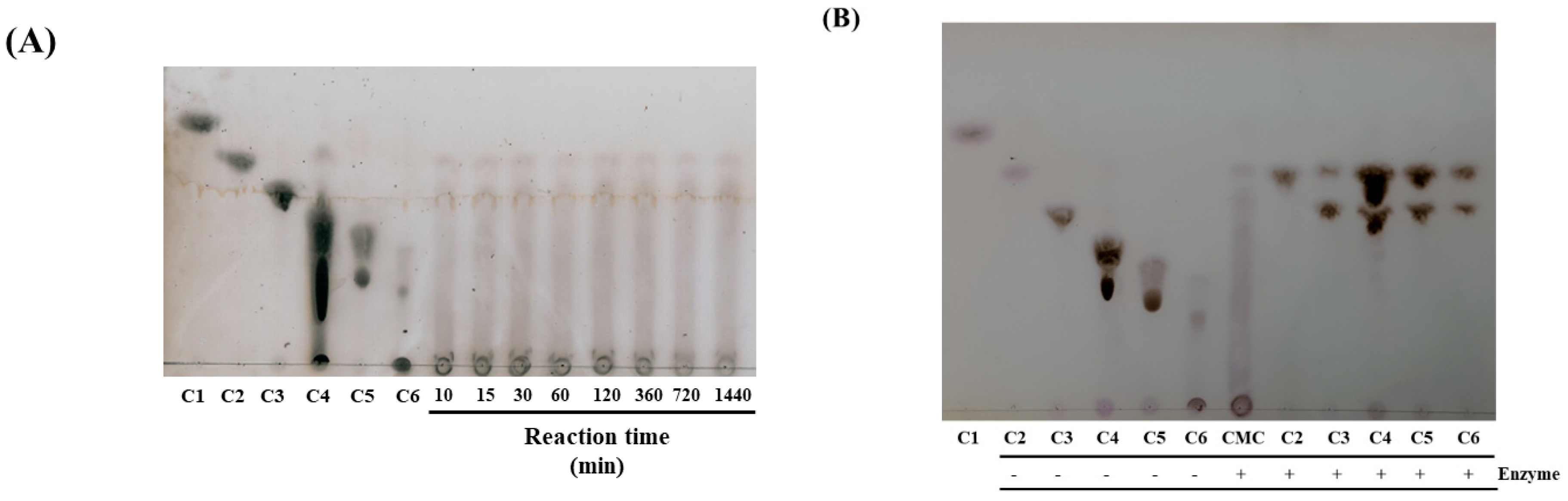
3.6. TLC Analysis of the Hydrolysate Produced by CelAJ93
3.7. Strain Deposit and GenBank Accession Number
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Klemm, D.; Heublein, B.; Fink, H.-P.; Bohn, A. Cellulose: Fascinating Biopolymer and Sustainable Raw Material. Angew. Chem. Int. Ed. 2005, 44, 3358–3393. [Google Scholar] [CrossRef] [PubMed]
- Shida, Y.; Furukawa, T.; Ogasawara, W. Deciphering the Molecular Mechanisms behind Cellulase Production in Trichoderma reesei, the Hyper-Cellulolytic Filamentous Fungus. Biosci. Biotechnol. Biochem. 2016, 80, 1712–1729. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.K. Cellulases and Related Enzymes in Biotechnology. Adv. Biotechnol. 2000, 1, 355–383. [Google Scholar] [CrossRef]
- Perez, J.; Munoz-Dorado, J.; de la Rubia, T.; Martinez, J. Biodegradation and Biological Treatments of Cellulose, Hemicellulose and Lignin: An overview. Int. Microbiol. 2002, 5, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Cavaco-Paulo, A. Mechanism of Cellulose Action in Textile Processes. Carbohydr. Polym. 1998, 37, 273–277. [Google Scholar] [CrossRef]
- Thomas, L.; Joseph, A.; Gottumukkala, L.D. Xylanase and Cellulase Systems of Clostridium sp.: An Insight on Molecular Approaches for Strain Improvement. Bioresour. Technol. 2014, 158, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.Y.; Gao, W.; Jin, I.H.; Yoo, S.Y.; Lee, J.H.; Chung, C.H.; Lee, J.W. Pretreatment and Saccharification of Rice Hulls for the Production of Fermentable Sugars. Biotechnol. Bioprocess Eng. 2009, 14, 828–834. [Google Scholar] [CrossRef]
- Cherry, J.R.; Fidantsef, A.L. Directed Evolution of Industrial Enzymes: An Update. Curr. Opin. Biotechnol. 2003, 14, 438–443. [Google Scholar] [CrossRef]
- Dos Santos, Y.Q.; de Veras, B.O.; de França, A.F.J.; Gorlach-Lira, K.; Velasques, J.; Migliolo, L.; Dos Santos, E.A. A New Salt-Tolerant Thermostable Cellulase from a Marine Bacillus sp. Strain. J. Microbiol. Biotechnol. 2018, 28, 1078–1085. [Google Scholar] [CrossRef]
- Zou, Z.; Zhao, Y.; Zhang, T.; Xu, J.; He, A.; Deng, Y. Efficient Isolation and Characterization of a Cellulase Hyperproducing Mutant Strain of Trichoderma reesei. J Microbiol. Biotechnol. 2018, 28, 1473–1481. [Google Scholar] [CrossRef]
- Nakamura, K.; Kappamura, K. Isolation and Identification of Crystalline Cellulose Hydrolyzing Bacterium and its Enzymatic Properties. J. Ferment. Technol. 1982, 60, 343–348. [Google Scholar]
- Galkiewicz, J.P.; Kellogg, C.A. Cross-kingdom Amplification Using Bacteria-specific Primers: Complications for Studies of Coral Microbial Ecology. Appl. Environ. Microbiol. 2008, 74, 7828–7831. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Molecular Cloning. A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, A.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Data Base. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Lee, J.H.; Jung, Y.Y.; Kim, M.J.; Kim, S.I.; Kim, B.K.; Lim, Y.W. ExTaxon: A Web-Based Tool for the Identification of Prokaryotes Based on 16S Ribosomal RNA Gene Sequences. Int. J. Syst. Evol. Microbiol. 2007, 57, 2259–2261. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Ghose, T.K. Measurement of Cellulase Activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Lim, J.H.; Lee, C.R.; Dhakshnamoorthy, V.; Park, J.S.; Hong, S.K. Molecular Characterization of Streptomyces coelicolor A(3) SCO6548 as a Cellulose 1,4-β-Cellobiosidase. FEMS Microbiol. Lett. 2016, 363, fnv245. [Google Scholar] [CrossRef]
- Park, S.; Oh, K.H.; Lee, S.Y.; Oh, T.K.; Yoon, J.H. Cellulophaga geojensis sp. nov., a member of the family Flavobacteriaceae isolated from marine sand. Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 6, 1354–1358. [Google Scholar] [CrossRef]
- Johansen, J.E.; Nielsen, P.; Sjøholm, C. Description of Cellulophaga baltica gen. nov., sp. nov. and Cellulophaga fucicola gen. nov., sp. nov. and reclassification of [Cytophaga] lytica to Cellulophaga lytica gen. nov., comb. nov. Int. J. Syst. Bacteriol. 1999, 49 Pt 3, 1231–1240. [Google Scholar] [CrossRef]
- Kahng, H.Y.; Chung, B.S.; Lee, D.H.; Jung, J.S.; Park, J.H.; Jeon, C.O. Cellulophaga tyrosinoxydans sp. nov., a tyrosinase-producing bacterium isolated from seawater. Int. J. Syst. Evol. Microbiol. 2009, 59 Pt 4, 654–657. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Han, S.O.; Yukawa, H.; Inui, M.; Doi, R.H. Molecular cloning and transcriptional and expression analysis of engO, encoding a new noncellulosomal family 9 enzyme, from Clostridium cellulovorans. J. Bacteriol. 2005, 187, 4884–4889. [Google Scholar] [CrossRef] [PubMed]
- Park, J.D.; Kim, Y.A.; Yoon, K.-H. Properties of a Bacillus licheniformis cellulase produced by recombinant Escherichia coli. Korean J. Microbiol. 2009, 45, 257–262. [Google Scholar]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Schneewind, O.; Missiakas, D. Sec-secretion and sortase-mediated anchoring of proteins in Gram-positive bacteria. Biochim. Biophys. Acta. 2014, 1843, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, W.; Han, F. Expression and Characterization of a Cold-adapted, Thermotolerant and Denaturant-stable GH5 Endoglucanase Celal_2753 that Withstands Boiling from the Psychrophilic Bacterium Cellulophaga algicola IC166(T). Biotechnol. Lett. 2016, 38, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Lee, E.J.; Lee, S.U.; Li, J.; Chung, C.H.; Lee, J.W. Enhanced Carboxymethylcellulase Production by a Newly Isolated Marine Bacterium, Cellulophaga lytica LBH-14, using rice bran. J. Microbiol. Biotechnol. 2012, 22, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Chen, Y.C.; Huang, C.T.; Hseu, R.S. Cloning and Characterization of a Thermostable and pH-stable Cellobiohydrolase from Neocallimastix patriciarum J11. Protein Expr. Purif. 2013, 90, 153–159. [Google Scholar] [CrossRef]
- Chang, L.; Ding, M.; Bao, L.; Chen, Y.; Zhou, J.; Lu, H. Characterization of a Bifunctional Xylanase/Endoglucanase from Yak Rumen Microorganisms. Appl. Microbiol. Biotechnol. 2011, 90, 1933–1942. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Zhu, Y.; Han, L.; Hefferon, K.L.; Silvaggi, N.R.; Wilson, D.B.; McBride, M.J. Periplasmic Cytophaga hutchinsonii Endoglucanases Are Required for Use of Crystalline Cellulose as the Sole Source of Carbon and Energy. Appl. Environ. Microbiol. 2016, 82, 4835–4845. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Volume (mL) | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 880 | 2551.12 | 110,588.57 | 43.35 | 1.00 |
| Ammonium sulfate precipitation | 75 | 279.68 | 14,705.36 | 52.58 | 1.21 |
| Diethylaminoethyl sepharose Fast Flow chromatography | 15 | 14.85 | 5721.98 | 385.32 | 8.89 |
| Superdex 75 chromatography | 2 | 1.92 | 1838.03 | 957.31 | 22.98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.S.; Chi, W.-J.; Hong, S.-K. Molecular Characterization of an Endo-β-1,4-Glucanase, CelAJ93, from the Recently Isolated Marine Bacterium, Cellulophaga sp. J9-3. Appl. Sci. 2019, 9, 4061. https://doi.org/10.3390/app9194061
Kim DS, Chi W-J, Hong S-K. Molecular Characterization of an Endo-β-1,4-Glucanase, CelAJ93, from the Recently Isolated Marine Bacterium, Cellulophaga sp. J9-3. Applied Sciences. 2019; 9(19):4061. https://doi.org/10.3390/app9194061
Chicago/Turabian StyleKim, Da Som, Won-Jae Chi, and Soon-Kwang Hong. 2019. "Molecular Characterization of an Endo-β-1,4-Glucanase, CelAJ93, from the Recently Isolated Marine Bacterium, Cellulophaga sp. J9-3" Applied Sciences 9, no. 19: 4061. https://doi.org/10.3390/app9194061
APA StyleKim, D. S., Chi, W. -J., & Hong, S. -K. (2019). Molecular Characterization of an Endo-β-1,4-Glucanase, CelAJ93, from the Recently Isolated Marine Bacterium, Cellulophaga sp. J9-3. Applied Sciences, 9(19), 4061. https://doi.org/10.3390/app9194061